
The Two Domains of the Avian Double-β-Defensin AvBD11 Have Different Ancestors, Common with Potential Monodomain Crocodile and Turtle Defensins


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Sequences and Net Charge Determination
2.2. Multiple Sequence Alignment (MSA)
2.3. Construction of Phylogenetic Trees
2.4. Positive Selection Calculation
2.5. Searching for Homolog Sequences of AvBD11 in Non-Avian Sauropsids
3. Results and Discussion
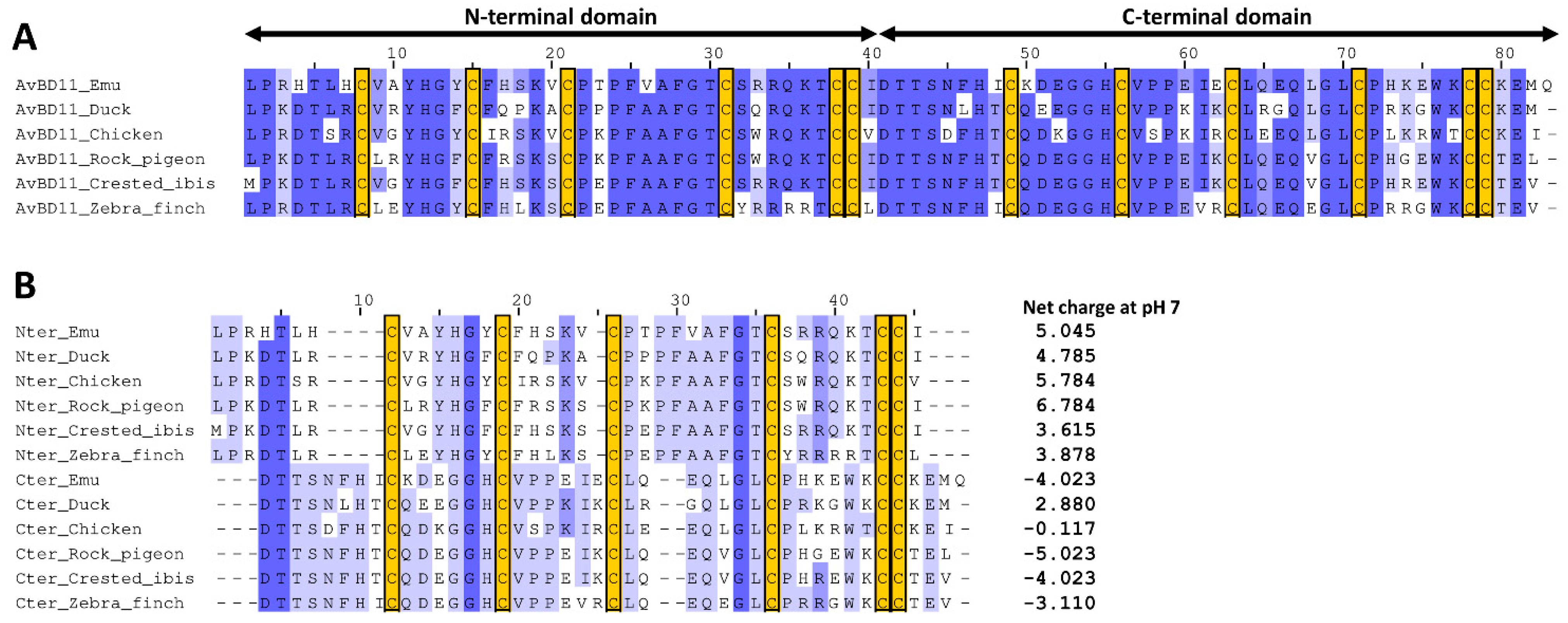
3.1. Degree of Divergence of the N- and C-Terminal Domains of AvBD11
3.2. Analysis of Positive Selection and Detection of Amino Acids under Potential Convergent Evolution in AvBD11 Sequences
3.3. Methodological Approach Used to Construct Phylogenetic Trees with β-Defensins
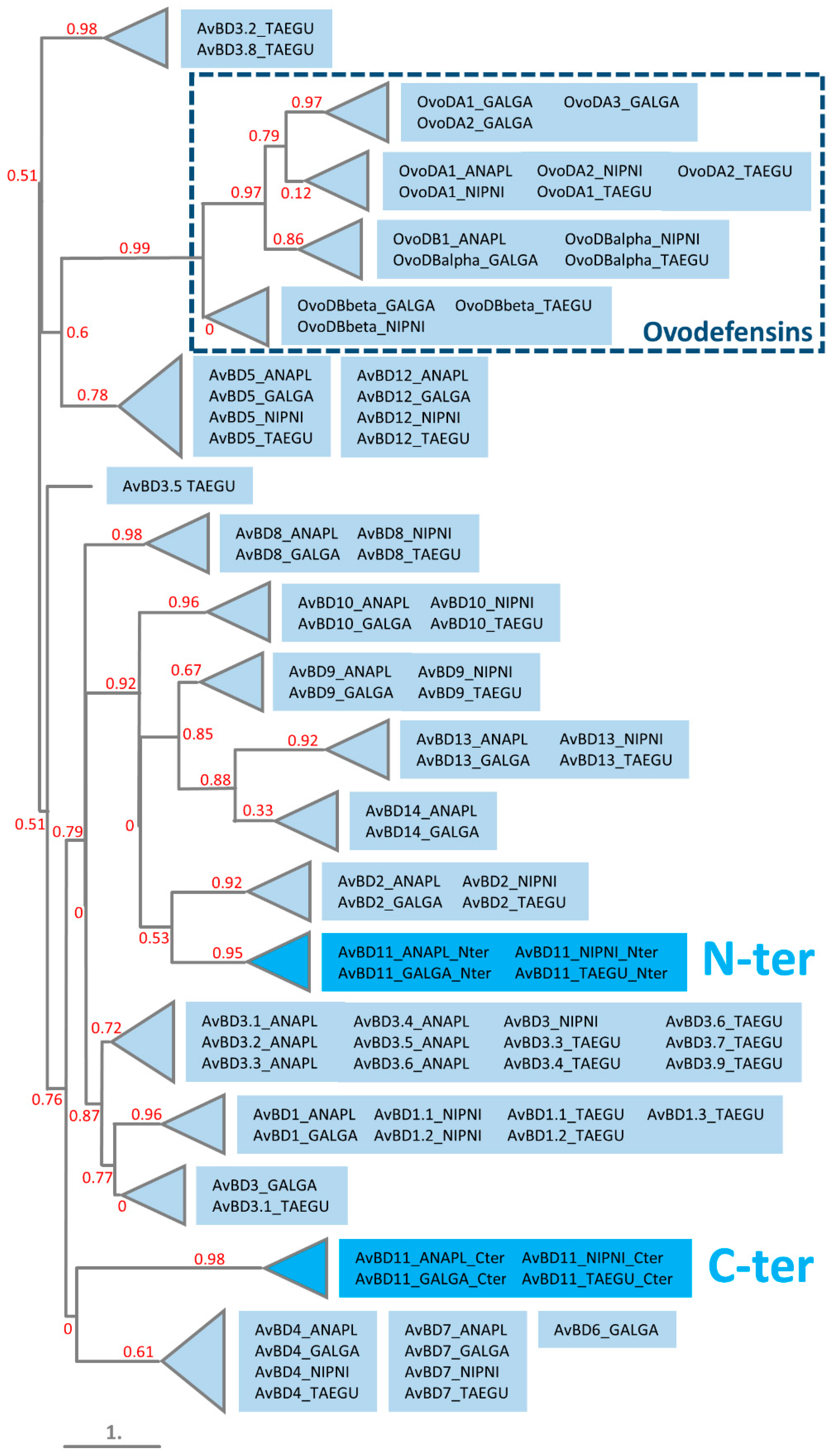
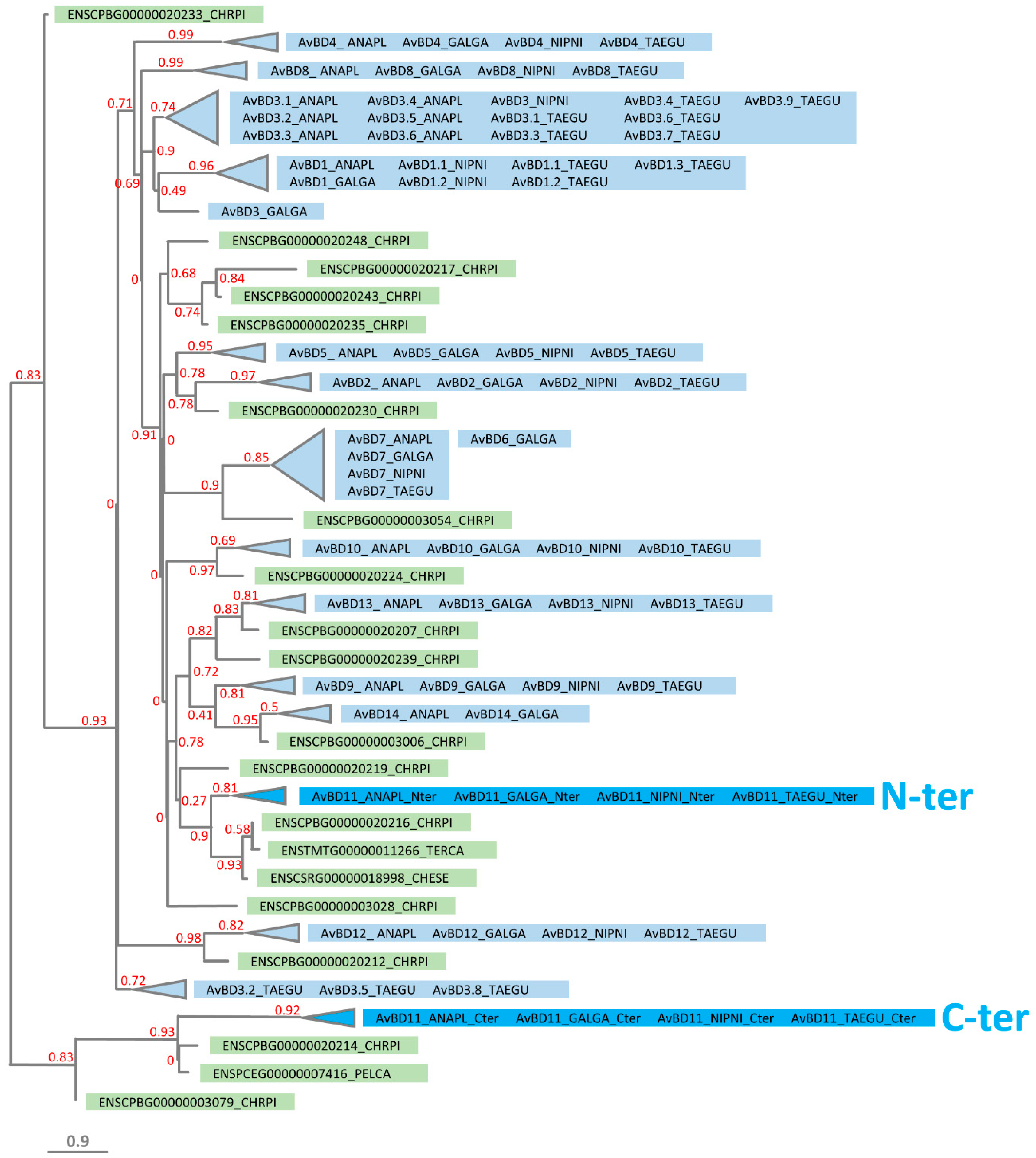
3.4. Phylogeny of N- and C-Terminal Domains of AvBD11 among AvBDs and OvoDs
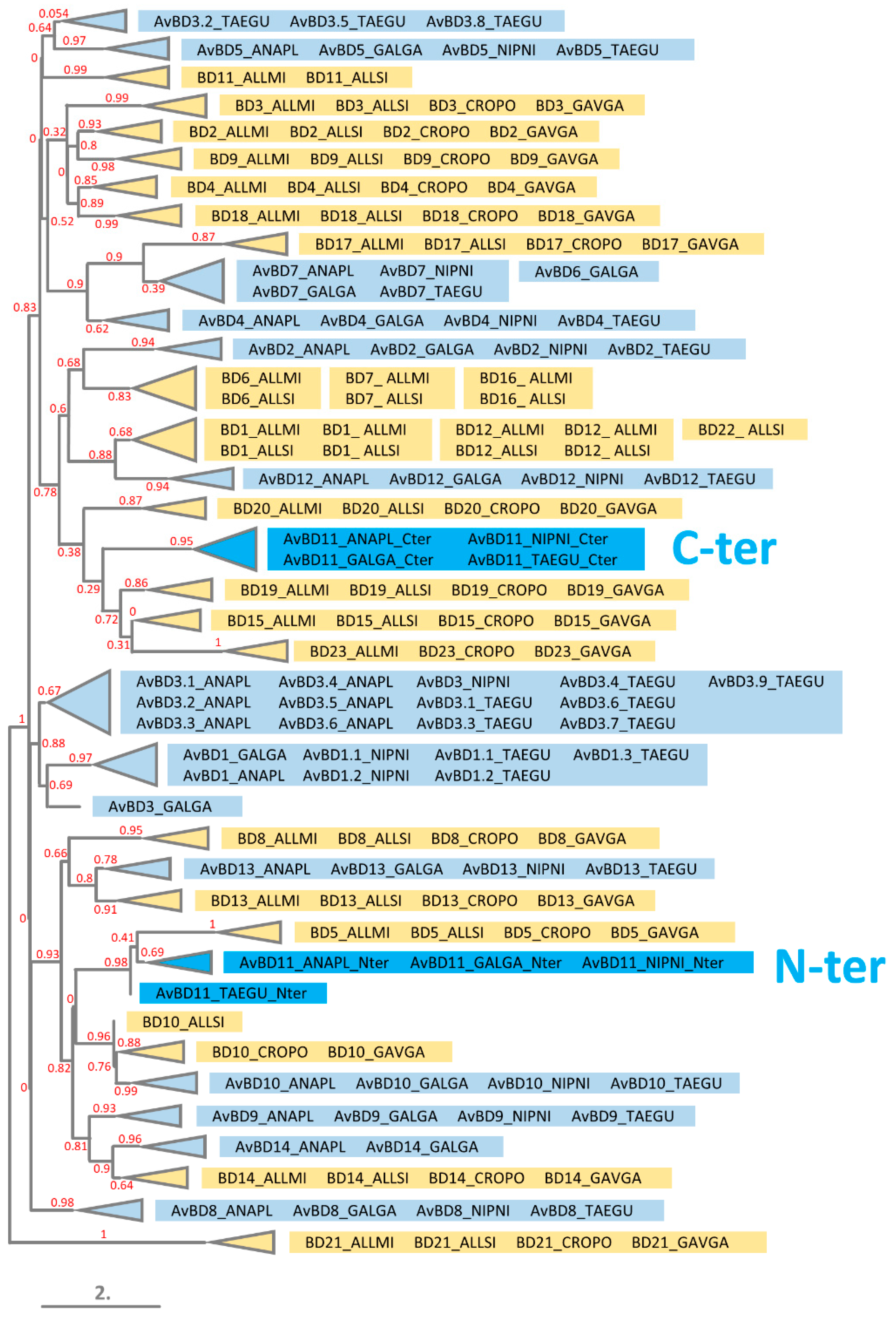
3.5. Phylogeny of N- and C-Terminal Domains of AvBD11 among AvBDs and Crocodile Defensins
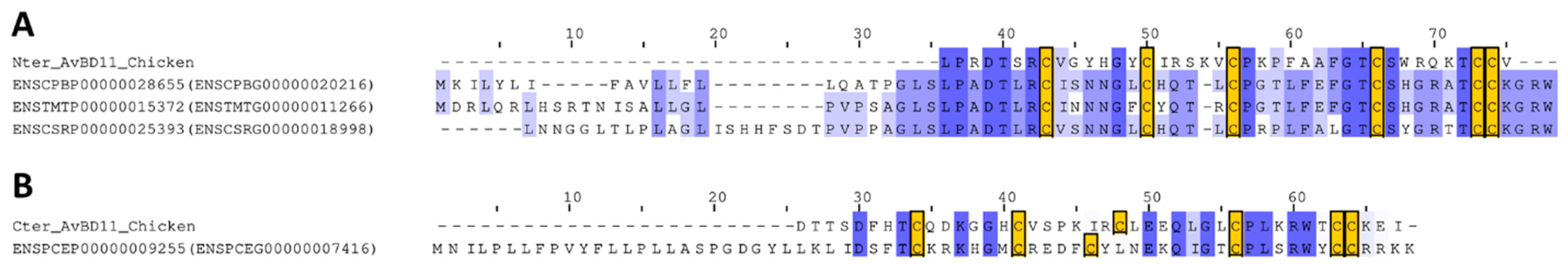
3.6. Identification of Reptilian Homologs for the N-Terminal and C-Terminal Domains of AvBD11
3.7. Phylogeny of N- and C-Terminal Domains of AvBD11 among AvBDs and Turtle Defensins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shafee, T.M.; Lay, F.T.; Phan, T.K.; Anderson, M.A.; Hulett, M.D. Convergent evolution of defensin sequence, structure and function. Cell Mol. Life Sci. 2017, 74, 663–682. [Google Scholar] [CrossRef] [PubMed]
- Shafee, T.M.; Lay, F.T.; Hulett, M.D.; Anderson, M.A. The Defensins Consist of Two Independent, Convergent Protein Superfamilies. Mol. Biol. Evol. 2016, 33, 2345–2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehrer, R.I.; Lu, W. alpha-Defensins in human innate immunity. Immunol. Rev. 2012, 245, 84–112. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L. Evolutionary diversification of the mammalian defensins. Cell Mol. Life Sci. 1999, 56, 94–103. [Google Scholar] [CrossRef]
- Semple, C.A.; Taylor, K.; Eastwood, H.; Barran, P.E.; Dorin, J.R. Beta-defensin evolution: Selection complexity and clues for residues of functional importance. Biochem. Soc. Trans. 2006, 34, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Prickett, M.D.; Gutowska, W.; Kuo, R.; Belov, K.; Burt, D.W. Evolution of the avian beta-defensin and cathelicidin genes. BMC Evol. Biol. 2015, 15, 188. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chen, D.; Yu, L.; Wei, Y.; Li, J.; Zhou, C. Genome-wide analysis of the ovodefensin gene family: Monophyletic origin, independent gene duplication and presence of different selection patterns. Infect Genet Evol. 2019, 68, 265–272. [Google Scholar] [CrossRef]
- Whenham, N.; Lu, T.C.; Maidin, M.B.; Wilson, P.W.; Bain, M.M.; Stevenson, M.L.; Stevens, M.P.; Bedford, M.R.; Dunn, I.C. Ovodefensins, an Oviduct-Specific Antimicrobial Gene Family, Have Evolved in Birds and Reptiles to Protect the Egg by Both Sequence and Intra-Six-Cysteine Sequence Motif Spacing. Biol. Reprod 2015, 92, 154. [Google Scholar] [CrossRef]
- Zhang, G.; Sunkara, L.T. Avian antimicrobial host defense peptides: From biology to therapeutic applications. Pharmaceuticals 2014, 7, 220–247. [Google Scholar] [CrossRef] [Green Version]
- Guyot, N.; Meudal, H.; Trapp, S.; Iochmann, S.; Silvestre, A.; Jousset, G.; Labas, V.; Reverdiau, P.; Loth, K.; Herve, V.; et al. Structure, function, and evolution of Gga-AvBD11, the archetype of the structural avian-double-beta-defensin family. Proc. Natl. Acad. Sci. USA 2020, 117, 337–345. [Google Scholar] [CrossRef]
- Herve, V.; Meudal, H.; Labas, V.; Rehault-Godbert, S.; Gautron, J.; Berges, M.; Guyot, N.; Delmas, A.F.; Nys, Y.; Landon, C. Three-dimensional NMR structure of Hen Egg Gallin (Chicken Ovodefensin) reveals a new variation of the beta-defensin fold. J. Biol. Chem. 2014, 289, 7211–7220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, K. Proteomic analysis of the chicken egg vitelline membrane. Proteomics 2008, 8, 2322–2332. [Google Scholar] [CrossRef]
- Kido, S.; Morimoto, A.; Kim, F.; Doi, Y. Isolation of a novel protein from the outer layer of the vitelline membrane. Biochem. J. 1992, 286 Pt 1, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, K. The chicken egg white proteome. Proteomics 2007, 7, 3558–3568. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.; Jeong, W.; Kim, J.; Yoshimura, Y.; Bazer, F.W.; Han, J.Y.; Song, G. Expression and regulation of beta-defensin 11 in the oviduct in response to estrogen and in ovarian tumors of chickens. Mol. Cell Endocrinol. 2013, 366, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Herve-Grepinet, V.; Rehault-Godbert, S.; Labas, V.; Magallon, T.; Derache, C.; Lavergne, M.; Gautron, J.; Lalmanach, A.C.; Nys, Y. Purification and characterization of avian beta-defensin 11, an antimicrobial peptide of the hen egg. Antimicrob Agents Chemother 2010, 54, 4401–4409. [Google Scholar] [CrossRef] [Green Version]
- Dalla Valle, L.; Benato, F.; Maistro, S.; Quinzani, S.; Alibardi, L. Bioinformatic and molecular characterization of beta-defensins-like peptides isolated from the green lizard Anolis carolinensis. Dev. Comp. Immunol. 2012, 36, 222–229. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Winter, K.; Hodgson, W.C.; Griesman, L.; Kwok, H.F.; Scanlon, D.; Karas, J.; Shaw, C.; Wong, L.; et al. Novel venom proteins produced by differential domain-expression strategies in beaded lizards and gila monsters (genus Heloderma). Mol. Biol. Evol. 2010, 27, 395–407. [Google Scholar] [CrossRef] [Green Version]
- van Hoek, M.L.; Prickett, M.D.; Settlage, R.E.; Kang, L.; Michalak, P.; Vliet, K.A.; Bishop, B.M. The Komodo dragon (Varanus komodoensis) genome and identification of innate immunity genes and clusters. BMC Genom. 2019, 20, 684. [Google Scholar] [CrossRef] [Green Version]
- Santana, F.L.; Estrada, K.; Ortiz, E.; Corzo, G. Reptilian beta-defensins: Expanding the repertoire of known crocodylian peptides. Peptides 2021, 136, 170473. [Google Scholar] [CrossRef]
- Yu, H.; Wang, H.; Liu, X.; Feng, L.; Qiao, X.; Cai, S.; Shi, N.; Wang, Y. Identification, eukaryotic expression and structure & function characterizations of beta-defensin like homologues from Pelodiscus sinensis. Dev. Comp. Immunol. 2017, 68, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.G.; Faircloth, B.C.; McCormack, J.E.; Brumfield, R.T.; Winker, K.; Glenn, T.C. More than 1000 ultraconserved elements provide evidence that turtles are the sister group of archosaurs. Biol. Lett. 2012, 8, 783–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemmell, N.J.; Rutherford, K.; Prost, S.; Tollis, M.; Winter, D.; Macey, J.R.; Adelson, D.L.; Suh, A.; Bertozzi, T.; Grau, J.H.; et al. The tuatara genome reveals ancient features of amniote evolution. Nature 2020, 584, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.F.; Dessimoz, C.; Gascuel, O. Survey of branch support methods demonstrates accuracy, power, and robustness of fast likelihood-based approximation schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef]
- Meslin, C.; Mugnier, S.; Callebaut, I.; Laurin, M.; Pascal, G.; Poupon, A.; Goudet, G.; Monget, P. Evolution of genes involved in gamete interaction: Evidence for positive selection, duplications and losses in vertebrates. PLoS ONE 2012, 7, e44548. [Google Scholar] [CrossRef]
- Grandchamp, A.; Monget, P. The membrane receptors that appeared before their ligand: The different proposed scenarios. PLoS ONE 2020, 15, e0231813. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shultz, A.J.; Sackton, T.B. Immune genes are hotspots of shared positive selection across birds and mammals. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Meslin, C.; Brimau, F.; Nagnan-Le Meillour, P.; Callebaut, I.; Pascal, G.; Monget, P. The evolutionary history of the SAL1 gene family in eutherian mammals. BMC Evol. Biol. 2011, 11, 148. [Google Scholar] [CrossRef] [Green Version]
- Dufourny, L.; Levasseur, A.; Migaud, M.; Callebaut, I.; Pontarotti, P.; Malpaux, B.; Monget, P. GPR50 is the mammalian ortholog of Mel1c: Evidence of rapid evolution in mammals. BMC Evol. Biol. 2008, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Rivas, E.; Eddy, S.R. Probabilistic phylogenetic inference with insertions and deletions. PLoS Comput. Biol. 2008, 4, e1000172. [Google Scholar] [CrossRef] [Green Version]
- Saurabh, K.; Holland, B.R.; Gibb, G.C.; Penny, D. Gaps: An elusive source of phylogenetic information. Syst. Biol. 2012, 61, 1075–1082. [Google Scholar] [CrossRef] [Green Version]
- Donath, A.; Stadler, P.F. Split-inducing indels in phylogenomic analysis. Algorithms Mol. Biol. 2018, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Dessimoz, C.; Gil, M. Phylogenetic assessment of alignments reveals neglected tree signal in gaps. Genome Biol. 2010, 11, R37. [Google Scholar] [CrossRef] [Green Version]
- Meade, K.G.; Higgs, R.; Lloyd, A.T.; Giles, S.; O’Farrelly, C. Differential antimicrobial peptide gene expression patterns during early chicken embryological development. Dev. Comp. Immunol. 2009, 33, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, O.; Ekblom, R. Evolution of a cluster of innate immune genes (beta-defensins) along the ancestral lines of chicken and zebra finch. Immunome Res. 2010, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilroy, D.; van Oosterhout, C.; Komdeur, J.; Richardson, D.S. Avian β-defensin variation in bottlenecked populations: The Seychelles warbler and other congeners. Conserv. Genet. 2016, 17, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Peng, K.; Gao, Q.; Xiang, M.; Liu, H.; Song, H.; Yang, K.; Huang, H.; Xiao, K. Molecular cloning, characterization and tissue distribution of two ostrich beta-defensins: AvBD2 and AvBD7. Gene 2014, 552, 1–7. [Google Scholar] [CrossRef]
- Chen, H.; Ma, M.Y.; Sun, L.; Fang, S.G.; Wan, Q.H. Genomic structure and evolution of beta-defensin genes in the golden pheasant and hwamei. Sci. Bull. 2015, 60, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Chen, H.; Chen, L.C.; Wang, B.B.; Sun, L.; Ma, M.Y.; Fang, S.G.; Wan, Q.H. The first report of a Pelecaniformes defensin cluster: Characterization of beta-defensin genes in the crested ibis based on BAC libraries. Sci. Rep. 2014, 4, 6923. [Google Scholar] [CrossRef] [Green Version]
- Ishige, T.; Hara, H.; Hirano, T.; Mannen, H.; Kono, T.; Hanzawa, K. Basic characterization of avian beta-defensin genes in the Japanese quail, Coturnix japonica. Anim. Sci. J. 2016, 87, 311–320. [Google Scholar] [CrossRef]
- Ma, D.; Lin, L.; Zhang, K.; Han, Z.; Shao, Y.; Wang, R.; Liu, S. Discovery and characterization of Coturnix chinensis avian beta-defensin 10, with broad antibacterial activity. J. Pept. Sci. 2012, 18, 224–232. [Google Scholar] [CrossRef]
- Hellgren, O.; Sheldon, B.C. Locus-specific protocol for nine different innate immune genes (antimicrobial peptides: Beta-defensins) across passerine bird species reveals within-species coding variation and a case of trans-species polymorphisms. Mol. Ecol. Resour. 2011, 11, 686–692. [Google Scholar] [CrossRef]
- Lynn, D.J.; Higgs, R.; Lloyd, A.T.; O’Farrelly, C.; Herve-Grepinet, V.; Nys, Y.; Brinkman, F.S.; Yu, P.L.; Soulier, A.; Kaiser, P.; et al. Avian beta-defensin nomenclature: A community proposed update. Immunol. Lett. 2007, 110, 86–89. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, G.; Yu, H.; Geng, Y.; Wu, W.; Tu, H.; Price, M.; Fan, Z.; Meng, Y.; Yue, B. Genome-wide analysis reveals the genomic features of the turkey vulture (Cathartes aura) as a scavenger. Mol. Genet. Genom. 2019, 294, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Hughes, A.L.; Ando, J.; Matsuda, Y.; Cheng, J.F.; Skinner-Noble, D.; Zhang, G. A genome-wide screen identifies a single beta-defensin gene cluster in the chicken: Implications for the origin and evolution of mammalian defensins. BMC Genom. 2004, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pais, F.S.; Ruy, P.C.; Oliveira, G.; Coimbra, R.S. Assessing the efficiency of multiple sequence alignment programs. Algorithms Mol. Biol. 2014, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Chiari, Y.; Cahais, V.; Galtier, N.; Delsuc, F. Phylogenomic analyses support the position of turtles as the sister group of birds and crocodiles (Archosauria). BMC Biol. 2012, 10, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guyot, N.; Landon, C.; Monget, P. The Two Domains of the Avian Double-β-Defensin AvBD11 Have Different Ancestors, Common with Potential Monodomain Crocodile and Turtle Defensins. Biology 2022, 11, 690. https://doi.org/10.3390/biology11050690
Guyot N, Landon C, Monget P. The Two Domains of the Avian Double-β-Defensin AvBD11 Have Different Ancestors, Common with Potential Monodomain Crocodile and Turtle Defensins. Biology. 2022; 11(5):690. https://doi.org/10.3390/biology11050690
Chicago/Turabian StyleGuyot, Nicolas, Céline Landon, and Philippe Monget. 2022. "The Two Domains of the Avian Double-β-Defensin AvBD11 Have Different Ancestors, Common with Potential Monodomain Crocodile and Turtle Defensins" Biology 11, no. 5: 690. https://doi.org/10.3390/biology11050690