
Foliar Spray or Soil Drench: Microalgae Application Impacts on Soil Microbiology, Morpho-Physiological and Biochemical Responses, Oil and Fatty Acid Profiles of Chia Plants under Alkaline Stress
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Location and Soil Analysis
2.2. Plant Material and Cultivation
2.3. Microorganisms Source and Culture
2.4. Treatments and Experimental Design
2.5. Morpho-Physiological Traits
2.6. Yield and Yield Components
2.7. Gas–Liquid Chromatography (GLC)
2.8. Biochemical Analysis
2.9. Nutrients Determinations
2.10. Soil pH and Microbial Count
2.11. Statistical Analysis
3. Results
3.1. Morpho-Physiological Traits
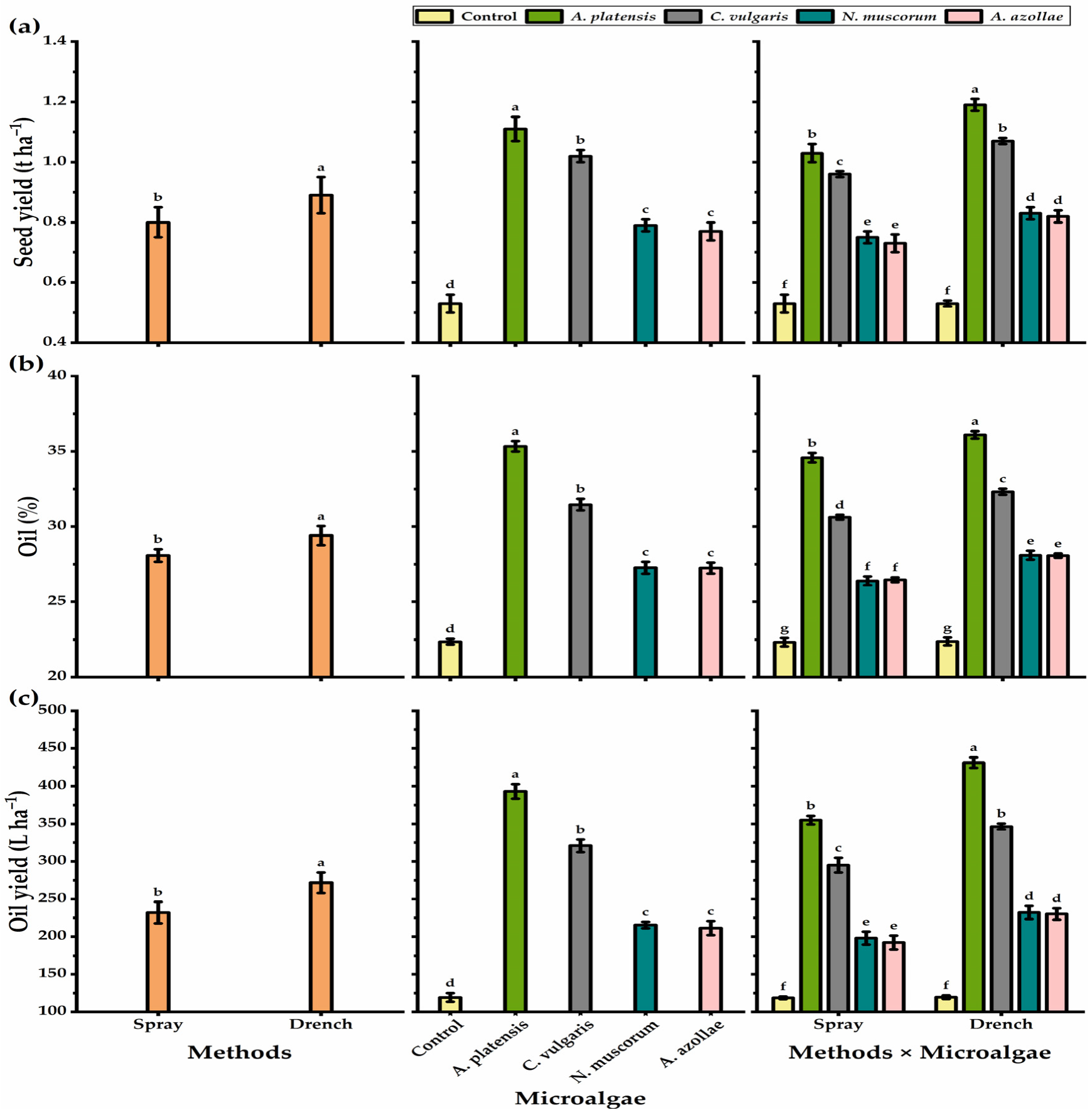
3.2. Yield and Yield Components
3.3. Fatty Acid Profile
3.4. Biochemical Traits
3.5. Nutrient Content
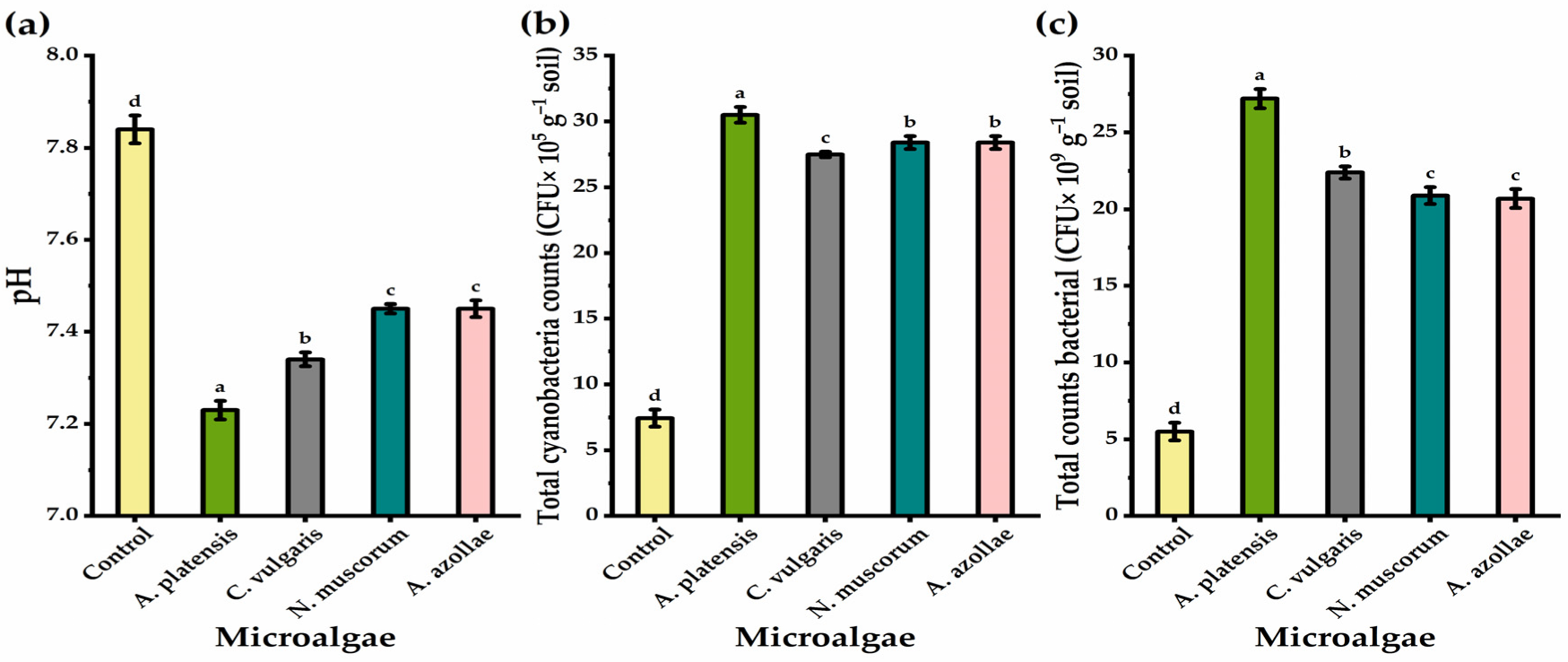
3.6. Soil pH and Microbial Count
4. Discussion
4.1. Growth and Yield Attributes
4.2. Oil and Fatty Acid Profiles
4.3. Physiological and Biochemical Analysis
4.4. Nutrient Content
4.5. Soil pH and Microbial Traits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cahill, J.P. Genetic diversity among varieties of chia (Salvia hispanica L.). Genet. Resour. Crop Evol. 2004, 51, 773–781. [Google Scholar]
- Busilacchi, H.; Quiroga, M.; Bueno, M.; Di Sapio, O.; Flores, V.; Severin, Y.C. Evaluation of Salvia hispanica L. cultivated in the south of Santa Fe (Argentina). Cult. Trop. 2013, 34, 55–59. [Google Scholar]
- Baginsky, C.; Arenas, J.; Escobar, H.; Garrido, M.; Valero, N.; Tello, D.; Pizarro, L.; Valenzuela, A.; Morales, L.; Silva, H. Growth and yield of chia (Salvia hispanica L.) in the Mediterranean and desert climates of Chile. Chil. J. Agric. Res. 2016, 76, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Aty, A.M.; Elsayed, A.M.; Salah, H.A.; Bassuiny, R.I.; Mohamed, S.A. Egyptian chia seeds (Salvia hispanica L.) during germination: Upgrading of phenolic profile, antioxidant, antibacterial properties and relevant enzymes activities. Food Sci. Biotechnol. 2021, 30, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Hrncic, M.K.; Ivanovski, M.; Cör, D.; Knez, Z. Chia seeds (Salvia Hispanica L.): An overview—Phytochemical profile, isolation methods, and application. Molecules 2020, 25, 11. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Aliberti, L.; Amato, M.; De Feo, V.; Camele, I. Chemical composition and antimicrobial activity of chia (Salvia hispanica L.) essential oil. Eur. Food Res. Technol. 2018, 244, 1675–1682. [Google Scholar] [CrossRef]
- De Falco, B.; Amato, M.; Lanzotti, V. Chia seeds products: An overview. Phytochem. Rev. 2017, 16, 745–760. [Google Scholar] [CrossRef]
- Ixtaina, V.Y.; Nolasco, S.M.; Tomas, M.C. Oxidative stability of chia (Salvia hispanica L.) seed oil: Effect of antioxidants and storage conditions. J. Am. Oil Chem. Soc. 2012, 89, 1077–1090. [Google Scholar] [CrossRef]
- Yanez-Yazlle, M.F.; Romano-Armada, N.; Acreche, M.M.; Rajal, V.B.; Irazusta, V.P. Halotolerant bacteria isolated from extreme environments induce seed germination and growth of chia (Salvia hispanica L.) and quinoa (Chenopodium quinoa Willd.) under saline stress. Ecotoxicol. Environ. Saf. 2021, 218, 112273. [Google Scholar] [CrossRef]
- Atteya, A.K.G.; El-Serafy, R.S.; El-Zabalawy, K.M.; Elhakem, A.; Genaidy, E.A.E. Brassinolide maximized the fruit and oil yield, induced the secondary metabolites, and stimulated linoleic acid synthesis of Opuntia ficus-indica oil. Horticulturae 2022, 8, 452. [Google Scholar] [CrossRef]
- Fageria, N.K.; Filho, M.P.B.; Moreira, A.; Guimarães, C.M. Foliar fertilization of crop plants. J. Plant Nutr. 2009, 32, 1044–1064. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, Z.; Ahmad, R.; Saifullah; Ashraf, M.Y. Foliar applied phosphorous enhanced growth, chlorophyll contents, gas exchange attributes and PUE in wheat (Triticum aestivum L.). J. Plant Nutr. 2015, 38, 1929–1943. [Google Scholar] [CrossRef]
- Tuyenm, D.D.; Zhang, H.M.; Xu, D.H. Validation and high-resolution mapping of a major quantitative trait locus for alkaline salt tolerance in soybean using residual heterozygous line. Mol. Breed. 2013, 31, 79–86. [Google Scholar] [CrossRef]
- El-Hady, O.A.; Abo-Sedera, S.A. Conditioning effect of composts and acrylamide hydrogels on a sandy calcareous soil. II-Physicobio-chemical properties of the soil. Int. J. Agric. Biol. 2006, 8, 876–884. [Google Scholar]
- Atteya, A.K.G.; Albalawi, A.N.; El-Serafy, R.S.; Albalawi, K.N.; Bayomy, H.M.; Genaidy, E.A.E. Response of Moringa oleifera seeds and fixed oil production to vermicompost and NPK fertilizers under calcareous soil conditions. Plants 2021, 10, 1998. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, K.Z.; Hasham, M.M.A.; El-Sheshtawy, A.-N.A.; El-Serafy, R.S.; Sheta, M.H. Biochar stimulated actual evapotranspiration and wheat productivity under water deficit conditions in sandy soil based on non-weighing lysimeter. Plants 2022, 11, 3346. [Google Scholar] [CrossRef]
- Sun, J.; Wang, J.; Guo, W.; Yin, T.; Zhang, S.; Wang, L.; Xie, D.; Zou, D. Identifcation of alkali-tolerant candidate genes using the NGS-assisted BSA strategy in rice. Mol. Breed. 2021, 41, 44. [Google Scholar] [CrossRef]
- Fu, S.F.; Sun, P.F.; Lu, H.Y.; Wei, J.Y.; Xiao, H.S.; Fang, W.T.; Cheng, B.Y.; Chou, J.Y. Plant growth-promoting traits of yeasts isolated from the phyllosphere and rhizosphere of Drosera spatulata Lab. Fungal Biol. 2016, 120, 433–448. [Google Scholar] [CrossRef]
- El-Serafy, R.S.; El-Sheshtawy, A.A. Effect of nitrogen fixing bacteria and moringa leaf extract on fruit yield, estragole content and total phenols of organic fennel. Sci. Hortic. 2020, 265, 109209. [Google Scholar] [CrossRef]
- Youssef, S.M.; Sayed, S.N.; Mahmoud, A.; Abdein, M.A.; Shamseldin, S.A. Response of stressed China aster (Callistephus chinensis) CV. Kamini plants to foliar application of benzyladenine (BA) and cycocel (CCC). Seybold Rep. 2020, 15, 30–42. [Google Scholar]
- La Bella, E.; Baglieri, A.; Rovetto, E.I.; Stevanato, P.; Puglisi, I. Foliar spray application of Chlorella vulgaris extract: Effect on the growth of lettuce seedlings. Agronomy 2021, 11, 308. [Google Scholar] [CrossRef]
- Yanni, Y.G.; Elashmouny, A.A.; Elsadany, A.Y. Differential response of cotton growth, yield and fiber quality to foliar application of Spirulina platensis and urea fertilizer. Asian J. Adv. Agric. Res. 2020, 12, 29–40. [Google Scholar]
- Abd El-Aal, A.A. Characterization of Anabaena azollae isolated from Azolla pinnata. Egypt. J. Agric. Res. 2013, 91, 801–809. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Kumaravel, R.; Gopalsamy, J.; Sikder, M.N.A.; Sampathkumar, P. Microalgae as bio-fertilizers for rice growth and seed yield productivity. Waste Biomass Valorization 2018, 9, 793–800. [Google Scholar] [CrossRef]
- Al-Sherif, E.M.; Abd El-Hameed, M.S.; Mahmoud, M.A.; Ahmed, H.S. Use of cyanobacteria and organic fertilizer mixture as soil bioremediation. Am.-Eurasian J. Agric. Environ. Sci. 2015, 15, 794–799. [Google Scholar]
- Abd Elhafiz, A.; Abd Elhafiz, A.; Gaur, S.S.; Hamdany, N.; Osman, M.; Lakshmi, T.V.R. Chlorella vulgaris and Chlorella pyrenoidosa live cells appear to be promising sustainable biofertilizer to grow rice, lettuce, cucumber and eggplant in the UAE soils. Recent Res. Sci. Technol. 2015, 7, 14–21. [Google Scholar]
- Page, A.L.; Miller, R.H.; Kenney, D.R. Methods of Soil Analysis. Chemical and Microbiological Properties, Part 2; SSSP, Inc.: Madison, WI, USA, 1982; pp. 39–41. [Google Scholar]
- Klute, A. Methods of Soil Analysis, Part 1, Physical and Mineralogical Methods, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1986. [Google Scholar]
- Allen, O.M. Experiments in Soil Bacteriology, 1st ed.; Burgess Publishing Co.: Minneapolis, MN, USA, 1959; Volume 11, pp. 1–42. [Google Scholar]
- Allen, M.M.; Stanier, R.Y. Selective isolation of blue-green algae from water and soil. Microbiology 1968, 51, 203–209. [Google Scholar] [CrossRef]
- Oster, J.D.; Jayawardane, N.S. Agricultural management of sodic soils. In Sodic soils: Distribution, Properties, Management and Environmental Consequences; Sumner, M.E., Naidu, R., Eds.; Oxford University Press: New York, NY, USA, 1998; pp. 125–147. [Google Scholar]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Zarrouk, C. Contribution a l’etude d’une cyanobacterie: Influence de divers facteurs physiques et chimiques sur la croissanceet la photosynthese de Spirulina maxima (Setchell et Gardner) Geitler. Ph. D. Thesis, University of Paris, Paris, France, 1966. [Google Scholar]
- Nichols, H.W.; Bold, H.C. Trichosarcina polymorpha gen. et Sp. Nov. J. Phycol. 1965, 1, 34–38. [Google Scholar] [CrossRef]
- Hegazi, A.Z.; Mostafa, S.S.; Ahmed, H.M. Influence of different cyanobacterial application methods on growth and seed production of common bean under various levels of mineral nitrogen fertilization. Nat. Sci. 2010, 8, 183–194. [Google Scholar]
- Souza, R.S.; Chaves, L.H.G. Initial growth of chia (Salvia hispanica L.) submitted to nitrogen, phosphorus and potassium fertilization. Aust. J. Crop Sci. 2017, 11, 610–615. [Google Scholar] [CrossRef]
- Jiang, C.; Johkan, M.; Hohjo, M.; Tsukagoshi, S.; Maruo, T. A correlation analysis on chlorophyll content and SPAD value in tomato leaves. HortResearch 2017, 71, 37–42. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophyll a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- AOAC Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International, 19th ed.; AOAC International: Washington, DC, USA, 2000. [Google Scholar]
- Alshallash, K.S.; Mohamed, M.F.; Dahab, A.A.; Abd El-Salam, H.S.; El-Serafy, R.S. Biostimulation of Plectranthus amboinicus (Lour.) spreng. with different yeast strains: Morphological performance, productivity, phenotypic plasticity, and antioxidant activity. Horticulturae 2022, 8, 887. [Google Scholar] [CrossRef]
- Dubois, M.; Smith, F.; Gillers, K.A.; Hamilton, J.K.; Robers, P.A. Colorimetric methods for determination of sugar and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Khatiwora, E.; Adsul, V.B.; Kulkarni, M.M.; Deshpande, N.R.; Kashalkar, R.V. Spectroscopic determination of total phenol and flavonoid contents of Ipomoea carnea. Int. J. ChemTech. Res. 2010, 2, 1698–1701. [Google Scholar]
- Shi, J.I.Y.; Zou, X.B.; Zhao, J.W.; Mel, H.; Wang, K.L.; Wang, X.; Chen, H. Determination of total flavonoids content in fresh Ginkgo biloba leaf with different colors using near infrared spectroscopy. Spectrochim. Acta A. Mol. Biomol. Spectrosc. 2012, 94, 271–276. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, W.; Xiong, S.; Tang, J.; Lou, Z.; Xie, M.; Xia, B.; Lin, L.; Liao, D. Determination of total flavonoids contents and antioxidant activity of Ginkgo biloba leaf by near-infrared reflectance method. Int. J. Anal. Chem. 2018, 2018, 8195784. [Google Scholar] [CrossRef] [Green Version]
- Rufino, M.S.M.; Alves, R.E.; De Brito, E.S.; De Morais, S.M.; Sampaio, C.G.; Pérez- Jiménez, J.; Saura-Calixto, F.D. Metodologia científica: Determinação da atividade antioxidante total em frutas pela captura do radical livre DPPH. Comunicado Técnico Embrapa 2007, 127, 1–4. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; New Jersy Prentice. Hall, Inc.: Hoboken, NJ, USA, 1973; p. 448. [Google Scholar]
- Wilde, S.A.; Corey, R.B.; Lyer, J.G.; Voigt, G.K. Soil and Plant Analysis for Tree Culture, 3rd ed.; Oxford and IBM Publishers: New Delhi, India, 1985; pp. 93–106. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soil, Plants and Water; University of California, Division of Agricultural Science: Berkeley, CA, USA, 1961. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Affiliated East-West Press: Delhi, India; Iowa State University Press: Ames, IA, USA, 1994. [Google Scholar]
- Koval, E.V.; Ogorodnikova, S.Y. The prospect of using the cyanobacterium Nostoc muscorum to improve vital activity of barley seedlings by various methods of seed treatment. BIO Web Conf. 2021, 36, 1–6. [Google Scholar] [CrossRef]
- Supraja, K.V.; Behera, B.; Balasubramanian, P. Efficacy of microalgal extracts as biostimulants through seed treatment and foliar spray for tomato cultivation. Ind. Crops Prod. 2020, 151, 112453. [Google Scholar]
- Verma, S.; Pandey, A.K. Enhancement of plant nutrient uptake by bacterial biostimulants. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 435–456. [Google Scholar]
- Godlewska, K.; Michalak, I.; Pacyga, P.; Baśladyńska, S.; Chojnacka, K. Potential applications of cyanobacteria: Spirulina platensis fltrates and homogenates in agriculture. World J. Microbiol. Biotechnol. 2019, 35, 80. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, I.K.; Sydney, E.B.; Sydney, A.C.N. Potential application of Spirulinain dermatology. J. Cosmet. Dermatol. 2022, 21, 4205–4214. [Google Scholar] [CrossRef]
- Kusvuran, S. Microalgae (Chlorella vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Hortic. Plant J. 2021, 7, 221–231. [Google Scholar] [CrossRef]
- Gitau, M.M.; Farkas, A.; Balla, B.; Ördög, V.; Futó, Z.; Maróti, G. Strain-specific biostimulant effects of Chlorella and Chlamydomonas green microalgae on Medicago truncatula. Plants 2021, 10, 1060. [Google Scholar] [CrossRef]
- Kumar, S.; Korra, T.; Singh, U.B.; Singh, S.; Bisen, K. Microalgal based biostimulants as alleviator of biotic and abiotic stresses in crop plants. In New and Future Developments in Microbial Biotechnology and Bioengineering: Sustainable Agriculture: Advances in Microbe-Based Biostimulants; Singh, H.B., Vaishnav, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 195–216. [Google Scholar]
- Guo, S.; Wang, P.; Wang, X.; Zou, M.; Liu, C.; Hao, J. Microalgae as biofertilizer in modern agriculture. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M.A., Xu, J.L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 397–411. [Google Scholar]
- Murata, M.M.; Morioka, L.R.I.; Marques, J.B.D.; Bosso, A.; Suguimoto, H.H. What do patents tell us about microalgae in agriculture? AMB Express 2021, 11, 154. [Google Scholar] [CrossRef]
- Wambacq, E.; Alloul, A.; Grunert, O.; Carrette, J.; Vermeir, P.; Spanoghe, J.; Sakarika, M.; Vlaeminck, S.E.; Haesaert, G. Aerobes and phototrophs as microbial organic fertilizers: Exploring mineralization, fertilization and plant protection features. PLoS ONE 2022, 17, e0262497. [Google Scholar] [CrossRef]
- Lu, Q.; Xiao, Y. From manure to high-value fertilizer: The employment of microalgae as a nutrient carrier for sustainable agriculture. Algal Res. 2022, 67, 102855. [Google Scholar] [CrossRef]
- Youssef, S.M.; Abdella, E.M.M.; Al-Elwany, O.A.; Alshallash, K.S.; Alharbi, K.; Ibrahim, M.T.S.; Tawfik, M.M.; Abu-Elsaoud, A.M.; Elkelish, A. Integrative application of foliar yeast extract and gibberellic acid improves morpho-physiological responses and nutrient uptake of Solidago virgaurea plant in alkaline soil. Life 2022, 12, 1405. [Google Scholar] [CrossRef]
- Refaay, D.A.; El-Marzoki, E.M.; Abdel-Hamid, M.I.; Haroun, S.A. Effect of foliar application with Chlorella vulgaris, Tetradesmus dimorphus, and Arthrospira platensis as biostimulants for common bean. J. Appl. Phycol. 2021, 33, 3807–3815. [Google Scholar] [CrossRef]
- El-Serafy, R.S.; El-Sheshtawy, A.A.; Dahab, A.A.; Al-Ashkar, I. Can yeast extract and chitosan-oligosaccharide improve fruit yield and modify the pharmaceutical active ingredients of organic fennel? Ind. Crops Prod. 2021, 173, 114130. [Google Scholar] [CrossRef]
- Mohamadineia, G.; Farahi, M.H.; Dastyaran, M. Foliar and soil drench application of humic acid on yield and berry properties of Askari grapevine. Agric. Commun. 2015, 3, 21–27. [Google Scholar]
- Faheed, F.A.; Abd-El Fattah, Z. Effect of Chlorella vulgaris as bio-fertilizer on growth parameters and metabolic aspects of lettuce plant. J. Agri. Soc. Sci. 2008, 4, 165–169. [Google Scholar]
- Agathokleous, E.; Kitao, M.; Shi, C.; Masui, N.; Abu-ElEla, S.; Hikino, K.; Satoh, F.; Koike, T. Ethylenediurea (EDU) spray effects on willows (Salix sachalinensis F. Schmid) grown in ambient or ozone-enriched air: Implications for renewable biomass production. J. For. Res. 2022, 33, 397–422. [Google Scholar] [CrossRef]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Stevanato, P.; Fascella, G.; Baglieri, A. Morpho-biometric and biochemical responses in lettuce seedlings treated by different application methods of Chlorella vulgaris extract: Foliar spray or root drench? J. Appl. Phycol. 2022, 34, 889–901. [Google Scholar] [CrossRef]
- Suchithra, M.R.; Muniswami, D.M.; Sri, M.S.; Usha, R.; Rasheeq, A.A.; Preethi, B.A. Effectiveness of green microalgae as biostimulants and biofertilizer through foliar spray and soil drench method for tomato cultivation. S. Afr. J. Bot. 2022, 146, 740–750. [Google Scholar] [CrossRef]
- Li, J.; Lens, P.N.; Ferrer, I.; Du Laing, G. Evaluation of selenium-enriched microalgae produced on domestic wastewater as biostimulant and biofertilizer for growth of selenium-enriched crops. J. Appl. Phycol. 2021, 33, 3027–3039. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of light conditions on microalgae growth and content of lipids, carotenoids, and fatty acid composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef]
- Nateghpour, B.; Kavoosi, G.; Mirakhorli, N. Amino acid profile of the peel of three citrus species and its effect on the combination of amino acids and fatty acids Chlorella vulgaris. J. Food Compos. Anal. 2021, 98, 103808. [Google Scholar] [CrossRef]
- Poh, Z.L.; Kadir, W.N.A.; Lam, M.K.; Uemura, Y.; Suparmaniam, U.; Lim, J.W.; Show, P.L.; Lee, K.T. The effect of stress environment towards lipid accumulation in microalgae after harvesting. Renew. Energy 2020, 154, 1083–1091. [Google Scholar] [CrossRef]
- Ragusa, I.; Nardone, G.N.; Zanatta, S.; Bertin, W.; Amadio, E. Spirulina for skin care: A bright blue future. Cosmetics 2021, 8, 7. [Google Scholar] [CrossRef]
- Khavari, F.; Saidijam, M.; Taheri, M.; Nouri, F. Microalgae: Therapeutic potentials and applications. Mol. Biol. Rep. 2021, 48, 4757–4765. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Bao, Q.; Wu, Y.; Chen, S.; Zhao, S.; Wu, H.; Fan, J. Evaluation of microalgae as immunostimulants and recombinant vaccines for diseases prevention and control in aquaculture. Front. Bioeng. Biotechnol. 2020, 8, 590431. [Google Scholar] [CrossRef] [PubMed]
- Castejón, N.; Luna, P.; Señoráns, F.J. Microencapsulation by spray drying of omega-3 lipids extracted from oilseeds and microalgae: Effect on polyunsaturated fatty acid composition. LWT 2021, 148, 111789. [Google Scholar] [CrossRef]
- Varia, J.; Kamaleson, C.; Lerer, L. Biostimulation with phycocyanin-rich Spirulina extract in hydroponic vertical farming. Sci. Hortic. 2022, 299, 111042. [Google Scholar] [CrossRef]
- Basheer, S.; Huo, S.; Zhu, F.; Qian, J.; Xu, L.; Cui, F.; Zou, B. Microalgae in human health and medicine. In Microalgae Biotechnology for Food, Health and High Value Products; Alam, M.A., Xu, J.L., Wang, Z., Eds.; Springer: Singapore, 2020; pp. 149–174. [Google Scholar]
- Ferrazzano, G.F.; Papa, C.; Pollio, A.; Ingenito, A.; Sangianantoni, G.; Cantile, T. Cyanobacteria and microalgae as sources of functional foods to improve human general and oral health. Molecules 2020, 25, 5164. [Google Scholar] [CrossRef]
- Shaikh, R.; Rizvi, A.; Pandit, S.; Desai, N.; Patil, R. Microalgae: Classification, bioactives, medicinal properties, industrial applications, and future prospectives. In An Integration of Phycoremediation Processes in Wastewater Treatment; Shah, M.P., Rodriguez-Couto, S., Vargas-De-La-Cruz, C., Biswas, J.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 451–486. [Google Scholar]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef]
- Malyan, S.K.; Bhatia, A.; Tomer, R.; Harit, R.C.; Jain, N.; Bhowmik, A.; Kaushik, R. Mitigation of yield-scaled greenhouse gas emissions from irrigated rice through Azolla, Blue-green algae, and plant growth–promoting bacteria. Environ. Sci. Pollut. Res. 2021, 28, 51425–51439. [Google Scholar] [CrossRef]
- Murchie, E.H.; Pinto, M.; Horton, P. Agriculture and the new challenges for photosynthesis research. New Phytol. 2009, 181, 532–552. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y. Microalgae: New source of plant biostimulants. Agronomy 2020, 10, 1240. [Google Scholar] [CrossRef]
- Danouche, M.; El Ghachtouli, N.; El Baouchi, A.; El Arroussi, H. Heavy metals phycoremediation using tolerant green microalgae: Enzymatic and non-enzymatic antioxidant systems for the management of oxidative stress. J. Environ. Chem. Eng. 2020, 8, 104460. [Google Scholar] [CrossRef]
- Atteya, A.K.G.; El-Serafy, R.S.; El-Zabalawy, K.M.; Elhakem, A.; Genaidy, E.A.E. Exogenously supplemented proline and phenylalanine improve growth, productivity, and oil composition of salted moringa by up-regulating osmoprotectants and stimulating antioxidant machinery. Plants 2022, 11, 1553. [Google Scholar] [CrossRef] [PubMed]
- Rachidi, F.; Benhima, R.; Kasmi, Y.; Sbabou, L.; El Arroussi, H. Evaluation of microalgae polysaccharides as biostimulants of tomato plant defense using metabolomics and biochemical approaches. Sci. Rep. 2021, 11, 930. [Google Scholar] [CrossRef] [PubMed]
- Rasool, S.; Hameed, A.; Azooz, M.M.; Siddiqi, T.O.; Ahmad, P. Salt stress: Causes, types and responses of plants. In Ecophysiology and Responses of Plants under Salt Stress; Springer: New York, NY, USA, 2013; pp. 1–24. [Google Scholar]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing sustainability by improving plant salt tolerance through macro- and micro-algal biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Mutale-joan, C.; Rachidi, F.; Mohamed, H.A.; El Mernissi, N.; Aasfar, A.; Barakate, M.; Mohammed, D.; Sbabou, L.; El Arroussi, H. Microalgae-cyanobacteria–based biostimulant effect on salinity tolerance mechanisms, nutrient uptake, and tomato plant growth under salt stress. J. Appl. Phycol. 2021, 33, 3779–3795. [Google Scholar] [CrossRef]
- Tran, T.L.N.; Miranda, A.F.; Abeynayake, S.W.; Mouradov, A. Differential production of phenolics, lipids, carbohydrates and proteins in stressed and unstressed aquatic plants, Azolla filiculoides and Azolla pinnata. Biology 2020, 9, 342. [Google Scholar] [CrossRef]
- Gr, S.; Yadav, R.K.; Chatrath, A.; Gerard, M.; Tripathi, K.; Govindsamy, V.; Abraham, G. Perspectives on the potential application of cyanobacteria in the alleviation of drought and salinity stress in crop plants. J. Appl. Phycol. 2021, 33, 3761–3778. [Google Scholar] [CrossRef]
- Singh, R.P.; Yadav, P.; Kujur, R.; Pandey, K.D.; Gupta, R.K. Cyanobacteria and salinity stress tolerance. In Cyanobacterial Lifestyle and Its Applications in Biotechnology; Singh, P.K., Fillat, M.F., Kumar, A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 253–280. [Google Scholar]
- Li, T.; Li, Y.; Sun, Z.; Xi, X.; Sha, G.; Ma, C.; Tian, Y.; Wang, C.; Zheng, X. Resveratrol alleviates the KCl salinity stress of Malus hupehensis Rhed. Front. Plant Sci. 2021, 12, 650485. [Google Scholar] [CrossRef]
- Rasheed, R.; Ashraf, M.A.; Ahmad, S.J.N.; Parveen, N.; Hussain, I.; Bashir, R. Taurine regulates ROS metabolism, osmotic adjustment, and nutrient uptake to lessen the effects of alkaline stress on Trifolium alexandrinum L. plants. S. Afr. J. Bot. 2022, 148, 482–498. [Google Scholar] [CrossRef]
- Geries, L.S.M.; Elsadany, A.Y. Maximizing growth and productivity of onion (Allium cepa L.) by Spirulina platensis extract and nitrogen-fixing endophyte Pseudomonas stutzeri. Arch. Microbiol. 2021, 203, 169–181. [Google Scholar] [CrossRef]
- Ertani, A.; Nardi, S.; Francioso, O.; Sanchez-Cortes, S.; Di Foggia, M.; Schiavon, M. Effects of two protein hydrolysates obtained from chickpea (Cicer arietinum L.) and Spirulina platensis on Zea mays (L.) plants. Front. Plant Sci. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gangwar, P.; Singh, R.; Trivedi, M.; Tiwari, R.K. Sodic soil: Management and reclamation strategies. In Environmental Concerns and Sustainable Development. Volume 2: Biodiversity, Soil and Waste Management; Shukla, V., Kumar, N., Eds.; Springer: Singapore, 2020; pp. 175–190. [Google Scholar]
- Schultz, E.; Chatterjee, A.; DeSutter, T.; Franzen, D. Sodic soil reclamation potential of gypsum and biochar additions: Influence on physicochemical properties and soil respiration. Commun. Soil Sci. Plant Anal. 2017, 48, 1792–1803. [Google Scholar] [CrossRef]
- Hu, L.; Zi, H.; Wu, P.; Wang, Y.; Lerdau, M.; Wu, X.; Wang, C. Soil bacterial communities in grasslands revegetated using Elymus nutans are largely influenced by soil pH and total phosphorus across restoration time. Land Degrad. Dev. 2019, 30, 2243–2256. [Google Scholar] [CrossRef]
- Priyanka; Kumar, C.; Chatterjee, A.; Wenjing, W.; Yadav, D.; Singh, P.K. Cyanobacteria: Potential and role for environmental remediation. In Abatement of Environmental Pollutants: Trends and Strategies; Singh, P., Kumar, A., Borthakur, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 193–202. [Google Scholar]
- Kapil, P.D.; Prabhu, S.N.; Deen, G.D.; Ajai, K.K. Cyanobacteria in alkaline soil and the effect of cyanobacteria inoculation with pyrite amendments on their reclamation. Biol. Fertil. Soils 2005, 41, 451–457. [Google Scholar]
- Kaushik, B.D. Algalization of rice in salt-affected soils. Ann. Agric. Res. 1994, 14, 105–106. [Google Scholar]
- Bayona-Morcillo, P.J.; Plaza, B.M.; Gómez-Serrano, C.; Rojas, E.; Jiménez-Becker, S. Effect of the foliar application of cyanobacterial hydrolysate (Arthrospira platensis) on the growth of Petunia x hybrida under salinity conditions. J. Appl. Phycol. 2020, 32, 4003–4011. [Google Scholar] [CrossRef]
- Alipour, S.; Kalari, S.; Morowvat, M.H.; Sabahi, Z.; Dehshahr, A. Green synthesis of selenium nanoparticles by cyanobacterium Spirulina platensis (abdf2224): Cultivation condition quality controls. BioMed Res. Int. 2021, 1, 6635297. [Google Scholar] [CrossRef]
- Zhang, F.; Man, Y.B.; Mo, W.Y.; Wong, M.H. Application of Spirulina in aquaculture: A review on wastewater treatment and fish growth. Rev. Aquac. 2020, 12, 582–599. [Google Scholar] [CrossRef]
- Alshuniaber, M.A.; Krishnamoorthy, R.; AlQhtani, W.H. Antimicrobial activity of polyphenolic compounds from Spirulina against food-borne bacterial pathogens. Saudi J. Biol. Sci. 2021, 28, 459–464. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil Characteristics | Value | |
|---|---|---|
| Particle Size Distribution (%) | 2019/2020 | 2020/2021 |
| Coarse sand | 7.39 | 7.42 |
| Fine sand | 40.51 | 40.48 |
| Silt | 18.89 | 18.92 |
| Clay | 33.21 | 33.18 |
| Texture class | Sandy clay Loam | |
| pH | 7.94 | 7.92 |
| ECe (dS m–1) | 3.77 | 3.75 |
| Soluble cations (mmolc L−1) | ||
| Ca++ | 5.13 | 5.10 |
| Mg++ | 4.12 | 4.10 |
| Na+ | 28.22 | 28.08 |
| K+ | 0.23 | 0.22 |
| Soluble anions (mmolc L−1) | ||
| CO3– | 16.82 | 16.84 |
| HCO3– | 3.15 | 2.95 |
| Cl– | 16.28 | 16.30 |
| SO4– | 1.45 | 1.41 |
| SAR | 13.44 | 13.37 |
| ESP% | 15.65 | 15.58 |
| Available Nutrients (%) | ||
| N | 0.17 | 0.18 |
| P | 0.20 | 0.22 |
| K | 0.29 | 0.30 |
| Biological (CFU g–1 soil) | ||
| Total bacterial count | 7.15 × 107 | 8.20 × 107 |
| Total cyanobacterial count | 6.14 × 102 | 6.89 × 102 |
| Treatment | Herb Fresh Weight (g) | Herb Dry Weight (g) | Total Chlorophyll (SPAD) | Total Carotenoids (mg mm−2) | |
|---|---|---|---|---|---|
| Season (S) | |||||
| SI | 400.2 ± 0.07 b | 101.2 ± 2.32 b | 61.3 ± 2.91 b | 0.39 ± b | |
| SII | 414.3 ± 0.05 a | 106.3 ± 2.21 a | 65.8 ± 2.90 a | 0.47 ± a | |
| Methods | |||||
| Spray | 417.2 ± 3.11 a | 105.0 ± 2.41 a | 62.9 ± 2.88 a | 0.43 ± 0.04 a | |
| Drench | 399.4 ± 3.14 b | 100.3 ± 2.71 b | 60.5 ± 2.67 b | 0.36 ± 0.05 b | |
| Microalgae | |||||
| Control | 323.1 ± 0.90 d | 80.5 ± 0.83 d | 36.5 ± 0.22 e | 0.23 ± 0.01 d | |
| Arthrospiraplatensis | 489.0 ± 3.40 a | 123.3 ± 1.85 a | 77.1 ± 0.67 a | 0.68 ± 0.02 a | |
| Chlorella vulgaris | 438.6 ± 3.54 b | 110.7 ± 1.63 b | 72.1 ± 0.69 b | 0.50 ± 0.03 b | |
| Nostoc muscorum | 394.2 ± 3.38 c | 99.6 ± 0.84 c | 63.9 ± 1.15 c | 0.28 ± 0.01 c | |
| Anabaena azollae | 396.8 ± 3.08 c | 99.4 ± 0.87 c | 59.0 ± 0.46 d | 0.28 ± 0.01 c | |
| Methods × Microalgae | |||||
| Spray | Control | 323.0 ± 1.17 g | 81.1 ± 0.42 g | 36.7 ± 0.03 i | 0.23 ± 0.01 g |
| A. platensis | 505.2 ± 2.87 a | 127.3 ± 0.72 a | 78.2 ± 0.61 a | 0.73 ± 0.01 a | |
| C. vulgaris | 453.0 ± 1.27 c | 114.3 ± 0.32 c | 73.6 ± 0.37 c | 0.58 ± 0.01 c | |
| N. muscorum | 401.5 ± 0.89 e | 101.4 ± 0.22 e | 66.3 ± 0.60 e | 0.30 ± 0.01 e | |
| A. azollae | 403.5 ± 0.91 e | 101.2 ± 0.55 e | 59.8 ± 0.30 g | 0.29 ± 0.01 e | |
| Drench | Control | 323.2 ± 1.64 g | 80.0 ± 1.72 g | 36.1 ± 0.39 i | 0.22 ± 0.01 g |
| A. platensis | 472.8 ± 1.77 b | 119.2 ± 0.47 b | 75.9 ± 0.70 b | 0.63 ± 0.01 b | |
| C. vulgaris | 424.2 ± 2.10 d | 107.0 ± 0.53 d | 70.6 ± 0.28 d | 0.43 ± 0.01 d | |
| N. muscorum | 386.9 ± 1.68 f | 97.7 ± 0.41 f | 61.6 ± 0.88 f | 0.26 ± 0.01 f | |
| A. azollae | 390.2 ± 1.54 f | 97.5 ± 0.39 f | 58.3 ± 0.61 h | 0.26 ± 0.01 f | |
| p-value | |||||
| S | <0.001 *** | <0.001 *** | 0.004 ** | <0.001 *** | |
| Methods | 0.002 ** | 0.008 ** | 0.006 ** | <0.001 *** | |
| Microalgae | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | |
| Methods × Microalgae | <0.001 *** | <0.001 *** | 0.006 ** | <0.001 *** | |
| Treatment | No. Inflorescence Plant−1 | Seed Yield Plant−1 (g) | 1000 Seed Weight (g) | |
|---|---|---|---|---|
| Season (S) | ||||
| SI | 16.3 ± 1.31 b | 14.56 ± 0.76 b | 1.80 ± 0.16 b | |
| SII | 19.5 ± 1.22 a | 16.78 ± 0.64 a | 1.89 ± 0.12 a | |
| Methods | ||||
| Spray | 17.9 ± 1.50 a | 14.40 ± 0.86 b | 1.81 ± 0.11 b | |
| Drench | 15.2 ± 1.38 b | 16.00 ± 0.1.11 a | 1.86 ± 0.11 a | |
| Microalgae | ||||
| Control | 9.2 ± 0.83 d | 9.59 ± 0.03 d | 1.08 ± 0.03 e | |
| Arthrospiraplatensis | 24.4 ± 0.95 a | 19.99 ± 0.69 a | 2.22 ± 0.01 a | |
| Chlorella vulgaris | 20.3 ± 0.83 b | 18.32 ± 0.45 b | 2.13 ± 0.02 b | |
| Nostoc muscorum | 14.8 ± 0.71 c | 14.19 ± 0.37 c | 1.93 ± 0.02 c | |
| Anabaena azollae | 13.9 ± 0.75 c | 13.91 ± 0.47 c | 1.81 ± 0.03 d | |
| Methods × Microalgae | ||||
| Spray | Control | 10.2 ± 0.62 a | 9.56 ± 0.03 f | 1.08 ± 0.00 g |
| A. platensis | 26.2 ± 0.64 a | 18.5 ± 0.05 b | 2.19 ± 0.00 b | |
| C. vulgaris | 21.5 ± 1.33 a | 17.3 ± 0.09 c | 2.09 ± 0.01 c | |
| N. muscorum | 16.2 ± 0.52 a | 13.51 ± 0.30 e | 1.92 ± 0.03 d | |
| A. azollae | 15.2 ± 0.58 a | 13.07 ± 0.54 e | 1.75 ± 0.00 f | |
| Drench | Control | 8.2 ± 0.52 a | 9.61 ± 0.05 f | 1.08 ± 0.01 g |
| A. platensis | 22.5 ± 0.86 a | 21.49 ± 0.36 a | 2.25 ± 0.01 a | |
| C. vulgaris | 19.2 ± 0.60 a | 19.30 ± 0.17 b | 2.16 ± 0.02 b | |
| N. muscorum | 13.4 ± 0.47 a | 14.87 ± 0.39 d | 1.94 ± 0.01 d | |
| A. azollae | 12.6 ± 0.86 a | 14.75 ± 0.32 d | 1.87 ± 0.01 e | |
| p-value | ||||
| S | 0.013 * | <0.001 *** | <0.001 *** | |
| Methods | 0.017 * | <0.004 ** | 0.021 * | |
| Microalgae | <0.001 *** | <0.001 *** | <0.001 *** | |
| Methods × Microalgae | 0.804 ns | 0.149 ns | 0.004 ** | |
| N. | Fatty Acids (%) | Spray | Drench | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | A. platensis | C. vulgaris | N. muscorum | A. azollae | Control | A. platensis | C. vulgaris | N. muscorum | A. azollae | ||
| 1 | Butyric (C4:0) | 0.01 | nd | nd | nd | nd | 0.04 | 0.07 | 0.13 | Nd | nd |
| 2 | Caproic (C6:0) | 0.01 | nd | nd | nd | 0.07 | 0.24 | 0.11 | 0.06 | Nd | 0.14 |
| 3 | Caprylic (C8:0) | 0.01 | nd | 0.09 | nd | 0.06 | 0.16 | 0.04 | 0.14 | Nd | 0.15 |
| 4 | Capric (C10:0) | 0.05 | 0.04 | 0.12 | 0.19 | 0.14 | 0.11 | 0.08 | 0.11 | Nd | 0.21 |
| 5 | Undecanoic (C11:0) | nd | nd | nd | 4.02 | nd | nd | Nd | nd | Nd | nd |
| 6 | Lauric (C12:0) | 0.02 | 0.04 | nd | 4.01 | 0.21 | 0.11 | 0.08 | 0.08 | Nd | nd |
| 7 | Myristic (C14:0) | 0.04 | 0.07 | nd | 0.71 | 0.08 | 0.06 | 0.04 | 0.08 | 0.05 | nd |
| 8 | Pentadecanoic (C15:0) | nd | 0.04 | nd | 0.28 | 0.09 | nd | 0.05 | nd | Nd | nd |
| 9 | Palmitic (C16:0) | 6.98 | 5.98 | 7.01 | 6.81 | 6.82 | 6.34 | 6.91 | 7.68 | 6.89 | 7.65 |
| 10 | Palmitoleic (C16:1) | 0.09 | nd | nd | nd | 0.11 | 0.17 | 0.18 | 0.16 | Nd | nd |
| 11 | Margaric (C17:0) | nd | nd | nd | nd | nd | nd | Nd | 0.06 | Nd | nd |
| 12 | Stearic (C18:0 n-9) | nd | 0.24 | nd | nd | nd | nd | Nd | nd | Nd | nd |
| 13 | Oleic (C18:1 n-9) | 11.01 | 6.78 | 9.38 | 7.01 | 15.59 | 12.06 | 5.69 | 7.6 | 8.01 | 7.88 |
| 14 | Linoleic (C18:2 n-6) | 17.26 | 20.15 | 18.19 | 18.63 | 17.01 | 16.01 | 20.97 | 19.92 | 20.03 | 19.27 |
| 15 | α-linolenic (C18:3 n-3) | 57.24 | 60.02 | 61.71 | 58.34 | 59.10 | 55.47 | 63.8 | 61.38 | 58.01 | 61.53 |
| 16 | γ-Linolenic (C18:3 n-6) | nd | 4.05 | 2.11 | nd | 0.62 | 0.94 | 1.46 | 1.98 | Nd | 0.57 |
| 17 | Arachidic (C20:0) | 7.23 | 0.83 | 0.66 | nd | 0.10 | 8.24 | 0.45 | 0.35 | 7.01 | 1.52 |
| 18 | Gadoleic (C20:1) | 0.05 | 0.82 | 0.42 | nd | nd | 0.05 | Nd | 0.14 | Nd | nd |
| 19 | Docosanoic (C22:0) | nd | 0.94 | 0.31 | nd | nd | nd | 0.07 | 0.13 | Nd | 0.68 |
| 20 | Tetracosanoic (C24:0) | nd | nd | nd | nd | nd | nd | Nd | nd | Nd | 0.4 |
| Area total | 100% | ||||||||||
| SFA | 7.04 | 7.31 | 7.32 | 11.81 | 7.20 | 6.51 | 7.15 | 8.03 | 6.94 | 8.73 | |
| MUFA | 18.38 | 8.43 | 10.46 | 7.01 | 15.80 | 20.52 | 6.32 | 8.25 | 15.02 | 9.40 | |
| PUFA | 74.50 | 84.22 | 82.01 | 76.97 | 76.73 | 72.42 | 86.23 | 83.28 | 78.04 | 81.37 | |
| n6/n3 ratio | 0.30 | 0.40 | 0.33 | 0.32 | 0.30 | 0.31 | 0.35 | 0.36 | 0.35 | 0.32 | |
| Treatment | Total Protein | Total Carbohydrates | Total Phenolic | Total Flavonoids | DPPH | |
|---|---|---|---|---|---|---|
| (%) | (mg 100 g−1) | (mg 100 g−1) | (μg mL−1) | |||
| Season (S) | ||||||
| SI | 24.1 ± 0.73 b | 36.1 ± 0.61 b | 34.5 ± 2.06 b | 15.7 ± 1.03 b | 65.2 ± 0.92 b | |
| SII | 25.4 ± 0.98 a | 37.5 ± 0.63 a | 36.5 ± 2.10 a | 17.6 ± 1.01 a | 66.9 ± 0.81 a | |
| Methods | ||||||
| Spray | 24.2 ± 0.89 b | 36.3 ± 0.58 b | 34.7 ± 2.04 b | 15.8 ± 1.15 b | 65.3 ± 0.86 b | |
| Drench | 25.2 ± 1.03 a | 37.3 ± 0.75 a | 36.3 ± 2.20 a | 17.5 ± 1.26 a | 66.8 ± 0.99 a | |
| Microalgae | ||||||
| Control | 18.3 ± 0.06 d | 33.9 ± 0.02 d | 21.2 ± 0.05 e | 9.64 ± 0.09 e | 60.7 ± 0.19 d | |
| Arthrospiraplatensis | 25.7 ± 0.24 b | 41.1 ± 0.46 a | 44.5 ± 0.37 a | 23.1 ± 0.39 a | 70.9 ± 0.35 a | |
| Chlorella vulgaris | 23.6 ± 0.23 c | 37.7 ± 0.23 b | 40.6 ± 0.44 b | 19.4 ± 0.46 b | 68.3 ± 0.44 b | |
| Nostoc muscorum | 28.0 ± 0.34 a | 35.7 ± 0.22 c | 35.2 ± 0.45 d | 15.9 ± 0.48 c | 65.2 ± 0.54 c | |
| Anabaena azollae | 28.0 ± 0.36 a | 35.7 ± 0.22 c | 36.0 ± 0.60 c | 15.3 ± 0.48 d | 65.1 ± 0.39 c | |
| Methods × Microalgae | ||||||
| Spray | Control | 18.3 ± 0.09 g | 33.8 ± 0.04 g | 21.1 ± 0.09 i | 9.46 ± 0.12 i | 60.7 ± 0.29 f |
| A. platensis | 25.1 ± 0.01 d | 40.1 ± 0.00 b | 43.7 ± 0.03 b | 22.3 ± 0.07 b | 70.2 ± 0.03 b | |
| C. vulgaris | 23.1 ± 0.01 f | 37.1 ± 0.01 d | 39.7 ± 0.06 d | 18.4 ± 0.07 d | 67.2 ± 0.03 c | |
| N. muscorum | 27.2 ± 0.03 b | 35.2 ± 0.04 f | 34.2 ± 0.07 h | 14.2 ± 0.06 h | 64.0 ± 0.46 e | |
| A. azollae | 27.2 ± 0.01 b | 35.2 ± 0.03 f | 34.6 ± 0.05 g | 14.8 ± 0.07 g | 64.4 ± 0.44 e | |
| Drench | Control | 18.3 ± 0.10 g | 33.9 ± 0.03 g | 21.2 ± 0.01 i | 9.81 ± 0.04 i | 60.8 ± 0.32 f |
| A. platensis | 26.2 ± 0.02 c | 42.2 ± 0.03 a | 45.3 ± 0.15 a | 23.9 ± 0.33 a | 71.7 ± 0.21 a | |
| C. vulgaris | 24.1 ± 0.01 e | 38.2 ± 0.03 c | 41.6 ± 0.17 c | 20.4 ± 0.17 c | 69.4 ± 0.16 b | |
| N. muscorum | 28.7 ± 0.10 a | 36.2 ± 0.01 e | 36.2 ± 0.04 f | 16.4 ± 0.10 f | 66.3 ± 0.06 d | |
| A. azollae | 28.8 ± 0.09 a | 36.2 ± 0.02 e | 37.3 ± 0.07 e | 16.9 ± 0.03 e | 65.7 ± 0.31 d | |
| p-value | ||||||
| S | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | |
| Methods | 0.004 ** | <0.001 *** | <0.001 *** | 0.002 ** | <0.001 *** | |
| Microalgae | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | |
| Methods × Microalgae | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** | 0.011 * | |
| Treatment | P | K+ | Ca2+ | Fe2+ | Mg2+ | Zn2+ | Na+ | |
|---|---|---|---|---|---|---|---|---|
| (mg g−1 Dry Seed) | ||||||||
| Season (S) | ||||||||
| SI | 3.38 ± 0.35 a | 22.9 ± 1.11 b | 11.2 ± 0.64 b | 7.51 ± 0.35 b | 6.72 ± 0.37 b | 3.91 ± 0.21 b | 2.00 ± 0.14 b | |
| SII | 3.41 ± 0.32 a | 25.8 ± 1.14 a | 13.1 ± 0.54 a | 8.39 ± 0.42 a | 7.95 ± 0.42 a | 4.51 ± 0.18 a | 2.13 ± 0.13 a | |
| Methods | ||||||||
| Spray | 3.37 ± 0.39 a | 23.7 ± 1.17 b | 11.4 ± 0.54 b | 7.70 ± 0.36 b | 6.81 ± 0.36 b | 3.95 ± 0.17 b | 2.12 ± 0.15 a | |
| Drench | 3.43 ± 0.28 a | 25.3 ± 1.23 a | 12.7 ± 0.71 a | 8.30 ± 0.45 a | 7.65 ± 0.48 a | 4.20 ± 0.19 a | 2.01 ± 0.15 b | |
| Microalgae | ||||||||
| Control | 2.15 ± 0.01 c | 17.5 ± 0.15 e | 9.13 ± 0.02 d | 6.07 ± 0.02 d | 5.53 ± 0.10 d | 3.15 ± 0.01 d | 3.08 ± 0.02 a | |
| Arthrospira platensis | 4.62 ± 0.23 a | 30.7 ± 0.28 a | 15.2 ± 1.05 a | 10.4 ± 0.15 a | 10.1 ± 0.38 a | 5.07 ± 0.03 a | 1.36 ± 0.05 e | |
| Chlorella vulgaris | 3.52 ± 0.19 ab | 27.6 ± 0.47 b | 13.8 ± 0.31 b | 8.88 ± 0.34 b | 7.64 ± 0.23 b | 4.50 ± 0.17 b | 1.87 ± 0.04 d | |
| Nostoc muscorum | 2.92 ± 0.04 bc | 23.5 ± 0.53 c | 11.1 ± 0.26 c | 7.34 ± 0.08 c | 6.46 ± 0.01 c | 3.83 ± 0.04 c | 1.94 ± 0.03 c | |
| Anabaena azollae | 3.80 ± 0.90 ab | 23.2 ± 0.39 d | 11.0 ± 0.25 c | 7.30 ± 0.09 c | 6.44 ± 0.13 c | 3.83 ± 0.05 c | 2.09 ± 0.02 b | |
| Methods × Microalgae | ||||||||
| Spray | Control | 2.17 ± 0.01 a | 17.2 ± 0.04i | 9.09 ± 0.02 a | 6.06 ± 0.04 g | 5.34 ± 0.06 h | 3.14 ± 0.01 g | 3.03 ± 0.02 a |
| A. platensis | 4.10 ± 0.01 a | 30.1 ± 0.01 b | 13.6 ± 1.67 a | 10.1 ± 0.04 b | 9.23 ± 0.06 b | 5.01 ± 0.01 b | 1.28 ± 0.06 a | |
| C. vulgaris | 3.10 ± 0.01 a | 26.6 ± 0.16 d | 13.1 ± 0.02 a | 8.11 ± 0.00 d | 7.14 ± 0.04 d | 4.11 ± 0.01 d | 1.82 ± 0.06 a | |
| N. muscorum | 2.84 ± 0.01 a | 22.4 ± 0.12 g | 10.5 ± 0.05 a | 7.13 ± 0.01 f | 6.18 ± 0.10 f | 3.75 ± 0.03 f | 1.90 ± 0.05 a | |
| A. azollae | 4.66 ± 1.82 a | 22.4 ± 0.26 g | 10.5 ± 0.03 a | 7.14 ± 0.01 f | 6.15 ± 0.04 f | 3.75 ± 0.06 f | 2.04 ± 0.03 a | |
| Drench | Control | 2.13 ± 0.01 a | 17.9 ± 0.03 h | 9.16 ± 0.04 a | 6.08 ± 0.03 g | 5.73 ± 0.10 g | 3.15 ± 0.02 g | 3.12 ± 0.01 a |
| A. platensis | 5.14 ± 0.02 a | 31.3 ± 0.01 a | 16.9 ± 0.06 a | 10.8 ± 0.03 a | 10.9 ± 0.01 a | 5.12 ± 0.02 a | 1.43 ± 0.05 a | |
| C. vulgaris | 3.95 ± 0.03 a | 28.6 ± 0.21 c | 14.4 ± 0.23 a | 9.64 ± 0.06 c | 8.15 ± 0.04 c | 4.88 ± 0.06 c | 1.91 ± 0.05 a | |
| N. muscorum | 2.99 ± 0.01 a | 24.7 ± 0.20 e | 11.7 ± 0.07 a | 7.47 ± 0.07 e | 6.74 ± 0.04 e | 3.91 ± 0.01 e | 1.99 ± 0.01 a | |
| A. azollae | 2.95 ± 0.01 a | 23.9 ± 0.36 f | 11.6 ± 0.04 a | 7.54 ± 0.03 e | 6.73 ± 0.01 e | 3.92 ± 0.00 e | 2.13 ± 0.01 a | |
| p-value | ||||||||
| S | 0.057 ns | <0.001 *** | 0.013 * | <0.001 *** | <0.001 ** | <0.001 ** | 0.004 ** | |
| Methods | 0.887 ns | 0.011 * | 0.046 * | 0.002 ** | 0.003 ** | 0.001 ** | 0.034 * | |
| Microalgae | 0.007 ** | <0.0001 *** | <0.001 *** | <0.001 *** | <0.001 ** | <0.001 ** | <0.001 ** | |
| Methods × Microalgae | 0.184 ns | <0.001 *** | 0.092 ns | <0.001 *** | <0.001 ** | <0.001 ** | 0.853 ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, S.M.; El-Serafy, R.S.; Ghanem, K.Z.; Elhakem, A.; Abdel Aal, A.A. Foliar Spray or Soil Drench: Microalgae Application Impacts on Soil Microbiology, Morpho-Physiological and Biochemical Responses, Oil and Fatty Acid Profiles of Chia Plants under Alkaline Stress. Biology 2022, 11, 1844. https://doi.org/10.3390/biology11121844
Youssef SM, El-Serafy RS, Ghanem KZ, Elhakem A, Abdel Aal AA. Foliar Spray or Soil Drench: Microalgae Application Impacts on Soil Microbiology, Morpho-Physiological and Biochemical Responses, Oil and Fatty Acid Profiles of Chia Plants under Alkaline Stress. Biology. 2022; 11(12):1844. https://doi.org/10.3390/biology11121844
Chicago/Turabian StyleYoussef, Samah M., Rasha S. El-Serafy, Kholoud Z. Ghanem, Abeer Elhakem, and Azza A. Abdel Aal. 2022. "Foliar Spray or Soil Drench: Microalgae Application Impacts on Soil Microbiology, Morpho-Physiological and Biochemical Responses, Oil and Fatty Acid Profiles of Chia Plants under Alkaline Stress" Biology 11, no. 12: 1844. https://doi.org/10.3390/biology11121844