
Modification of Antibiotic Activity by Fixed Oil of the Artocarpus heterophyllus Almond against Standard and Multidrug-Resistant Bacteria Strains
 , , , , , ,
, , , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction of the Fixed Oil
2.3. Determination of Fatty Acids in the Fixed Oil
2.4. Antibacterial Analysis of the Fixed Oil
2.4.1. Preparation of Analysis Substances
2.4.2. Bacterial Material
2.4.3. Elaboration and Standardization of Bacterial Inoculum
2.4.4. Determination of the Minimum Inhibitory Concentration (MIC) Determination
2.4.5. Ability to Modulate Antibiotic Activity
2.4.6. Statistical Analysis
3. Results and Discussion
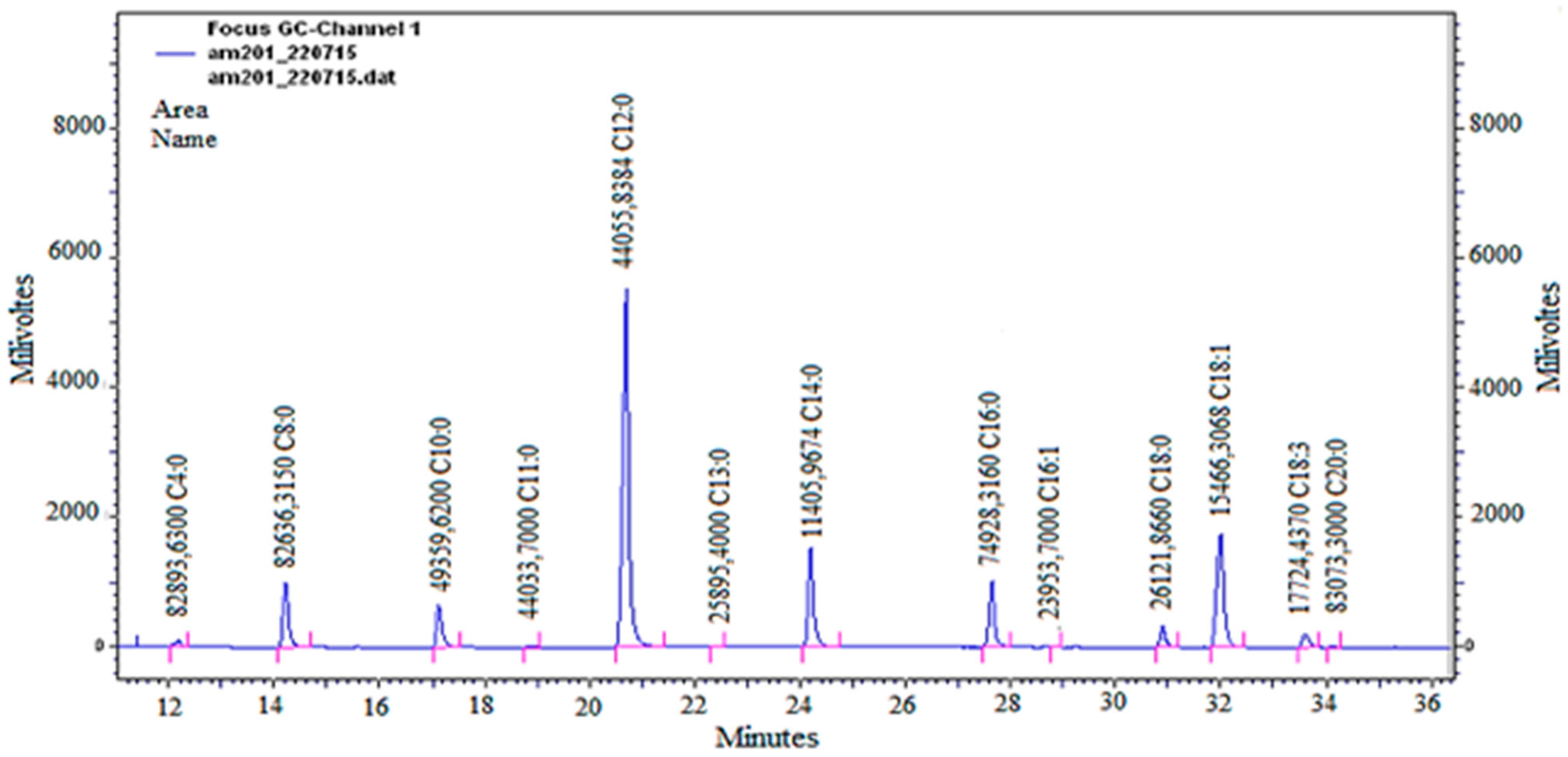
3.1. Analysis of Fatty Acid Profile of the Fixed Oil
3.2. Profile of Antibacterial Activity
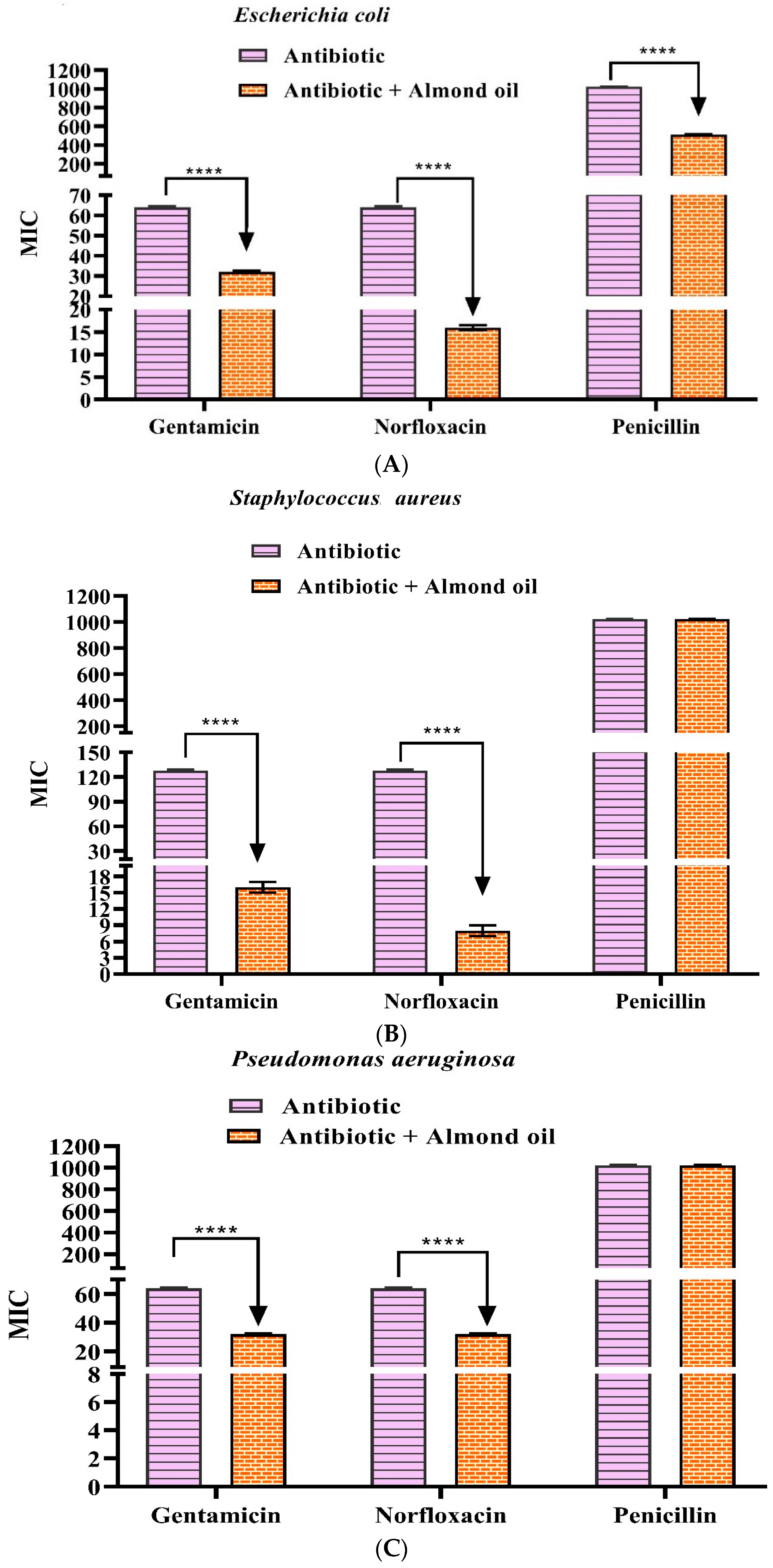
3.3. Antibiotic Modifying Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pereira, F.F.G.; Feitosa, M.K.S.B.; Costa, M.S.; Tintino, S.R.; Rodrigues, F.F.G.; Menezes, I.R.A.; Coutinho, H.D.M.; Costa, J.G.M.; Sousa, E.O. Characterization, antibacterial activity and antibiotic modifying action of the Caryocar coriaceum Wittm. pulp and almond fixed oil. Nat. Prod. Res. 2020, 34, 1–5. [Google Scholar] [CrossRef]
- Tintino, S.R.; Oliveira-Tintino, C.D.M.; Campina, F.F.; Costa, M.S.; Cruza, R.P.; Pereira, R.L.S.; Andrade, J.C.; Sousa, E.O.; Siqueira-Junior, J.P.; Coutinho, H.D.M.; et al. Cholesterol and ergosterol affect the activity of Staphylococcus aureus antibiotic efflux pumps. Microb. Pathogenesis. 2017, 104, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Nobre, C.B.; Sousa, E.O.; Silva, J.M.L.; Coutinho, H.D.; Costa, J.G. Chemical composition and antibacterial activity of fixed oils of Mauritia flexuosa and Orbignya speciosa associated with aminoglycosides. Eur. J. Integr. Med. 2018, 1, 84–89. [Google Scholar] [CrossRef]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.D.T.; Benjamim, M.A.; Silva, M.T.G.; Leite, D.O.D.; Sousa, E.O. Aspecto químico e bioatividade da jaca (Artocarpus heterophyllus): Uma revisão. In Pesquisas e Atualizações em Ciência dos Alimentos, 1st ed.; Agrom Food Academy: Jardim do Sérico, Brazil, 2022; Volume 2, pp. 196–205. [Google Scholar] [CrossRef]
- Sousa, E.O.; Costa, M.S.; Oliveira-Tintino, D.M.; Nonato, C.F.A.; Pinheiro, J.C.A.; Coutinho, H.D.M.; Menezes, I.R.A.; Costa, J.G.M. Analysis chemical of the fixed oil of Harconia speciosa and modulation of the antibiotic activity against MDR strains and other bacterial. Separations 2022, 9, 249. [Google Scholar] [CrossRef]
- Machado, J.F.; Costa, M.S.; Tintino, S.R.; Rodrigues, F.F.G.; Nobre, C.B.; Coutinho, H.D.M.; Costa, J.G.M.; Menezes, I.R.A.; Sousa, E.O. Antibiotic activity potentiation and physicochemical characterization of the fixed Orbignya speciosa almond oil against MDR Staphylococcus aureus and other bactéria. Antibiotics 2019, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, Y.F.; Costa, M.D.S.; Tintino, S.R.; Rocha, J.E.; Rodrigues, F.F.G.; Feitosa, M.K.S.B.; Menezes, I.R.A.; Coutinho, H.D.M.; Costa, J.G.M.; Sousa, E.O. Modulation of the antibiotic activity by Mauritia flexuosa (Buriti) fixed oil against Methicillin-Resistant Staphylococcus aureus (MRSA) and other Multidrug-Resistant (MDR) bacterial strains. Pathogens 2018, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Freitas, P.R.; Araújo, A.C.J.; Barbosa, C.R.S.; Muniz, D.F.; Silva, A.C.A.; Rocha, J.E.; Oliveira-Tintino, C.D.M.; Ribeiro-Filho, J.; Silva, L.E.; Confortin, C.; et al. GC-MS-FID and potentiation of the antibiotic activity of the essential oil of Baccharis reticulate (Ruiz & Pav.) Pers. and α-pinene. Ind. Crops Prod. 2020, 145, 1–5. [Google Scholar] [CrossRef]
- Fu, Y.H.; Guo, J.M.; Xie, Y.T.; Yu, X.M.; Su, Q.T.; Lei, Q.T.; Kong, L.Y.; Liu, Y.P. Prenylated chromones from thef fruits of Artocarpus heterophyllus and their potential Anti-HIV-1 activities. J. Agric. Food Chem. 2020, 68, 2024–2030. [Google Scholar] [CrossRef]
- Liu, Y.P.; Yu, X.M.; Zhang, W.; Wang, T.; Jiang, B.; Tang, H.X.; Su, Q.T.; Fu, Y.H. Prenylated chromones and flavonoids from Artocarpus heterophyllus with their potential antiproliferative and anti-inflammatory activities. Bioorg. Chem. 2020, 101, 104030. [Google Scholar] [CrossRef]
- Benjamim, M.A.; Nascimento, N.A.; Silva, M.M.A.; Feitosa, M.K.S.B.; Sousa, E.O. Elaboração e análise de bolo enriquecido com a farinha proveniente da semente da jaca (Artocarpus heterophyllus). In Pesquisas e Atualizações em Ciência dos Alimentos, 1st ed.; Agrom Food Academy: Jardim do Sérico, Brazil, 2022; Volume 2, pp. 37–42. [Google Scholar] [CrossRef]
- Nascimento, N.A.; Benjamim, M.A.; Silva, M.M.A.; Sousa, E.O. Aspecto nutricional da farinha da semente da jaca e uso na elaboração de produtos alimentícios. In Pesquisas e Atualizações em Ciência dos Alimentos, 1st ed.; Agrom Food Academy: Jardim do Sérico, Brazil, 2022; Volume 2, pp. 131–137. [Google Scholar] [CrossRef]
- Pereira, V.J.; Kaplan, M.A.C. The high bioactivity of Artocarpus—An exotic genus. Floresta Ambiente 2013, 20, 1–15. [Google Scholar] [CrossRef]
- Instituto Adolfo Lutz. Normas Analíticas do Instituto Adolfo Lutz. In Métodos Físico-Químicos Para Análise de Alimentos, 4th ed.; Instituto Adolfo Lutz: Brasília, Brazil, 2005. [Google Scholar]
- Ajayi, I.A. Comparative study of the chemical composition and mineral element content of Artocarpus heterophyllus and Treculia africana seeds and seed oils. Bioresour. Technol. 2008, 99, 5125–51259. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, M.R.F.; Souza, V.F.; Bomdespacho, L.Q.; Ascheri, J.L.R. Perfil de ácidos graxos no óleo extraído das farinhas dos caroços de jaca (Artocapus integrifólia L.), abacate (Persea gratíssima Gaertner F.) e seriguela (Spondias purpúrea L.). In Proceedings of the Anais do XII Congresso Brasileiro de Engenharia Química em Iniciação Científica [=Blucher Chemical Engineering Proceedings, v. 1, n.4], São Paulo, Brazil, 16–19 July 2017; pp. 88–93. [Google Scholar]
- Chan, L.W.; Hern, K.E.; Ngambenjawong, C.; Lee, K.; Kwon, E.J.; Hung, D.T.; Bhatia, S.N. Selective permeabilization of gram-negative bacterial membranes using multivalent peptide constructs for antibiotic sensitization. ACS Infect. Dis. 2021, 7, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Sales, D.L.; Oliveira, O.P.; Cabral, M.E.; Dias, D.Q.; Kerntopf, M.R.; Coutinho, H.D.M.; Costa, J.G.M.; Freitas, F.R.; Ferreira, F.S.; Alves, R.R.; et al. Chemical identification and evaluation of the antimicrobial activity of fixed oil extracted from Rhinella jimi. Pharm. Biol. 2015, 53, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Mezni, F.; Aouadhi, C.; Khouja, M.L.; Khaldi, A.; Maaroufi, A. In vitro antimicrobial activity of Pistacia lentiscus L. edible oil and phenolic extract. Nat. Prod. Res. 2015, 29, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Rosa, A.; Marongiu, B.; Porcedda, S.; Falconieri, D.; Dessì, M.A.; Ozcelik, B.; Koc, U. Chemical composition and in vitro bioactivity of the volatile and fixed oils of Nigella sativa L. extracted by supercritical carbon dioxide. Ind. Crops. Prod. 2013, 46, 317–323. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Setzer, W.N. Chemical composition and biological activities of essential oils of Curcuma species. Nutrients 2018, 10, 1196. [Google Scholar] [CrossRef]
- Anzaku, A.A.; Akyala, J.I.; Juliet, A.; Obianuju, E.C. Antibacterial activity of lauric acid on some selected clinical isolates. Ann. Clin. Lab. Res. 2017, 5, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Sales, D.L.; Andrade, J.C.; Silva, A.R.P.; Tintino, S.R.; Oliveira-Tintino, C.D.M.; Delmondes, G.A.; Rocha, M.F.G.; Costa, J.G.M.; Alves, R.R.N.; Ferreira, F.S.; et al. Body fat modulated activity of Gallus gallus domesticus Linnaeus (1758) and Meleagris gallopavo Linnaeus (1758) in association with antibiotics against bacteria of veterinary interest. Microb. Pathog. 2018, 124, 163–169. [Google Scholar] [CrossRef]
- Kim, S.A.; Rhee, M.S. Highly enhanced bactericidal effects of medium chain fatty acids (caprylic, capric, and lauric acid) combined with edible plant essential oils (carvacrol, eugenol, β-resorcylic acid, trans-cinnamaldehyde, thymol, and vanillin) against Escherichia coli O157: H7. Food Control 2016, 60, 447–454. [Google Scholar] [CrossRef]
- Bera, S.; Zhanel, G.G.; Schweizer, F. Design, synthesis, and antibacterial activities of neomycin-lipid conjugates: Polycationic lipids with potent gram-positive activity. J. Med. Chem. 2008, 51, 6160–6164. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.J. Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casillas-Vargas, G.; Ocasio-Malavé, C.; Medina, S.; Morales-Guzmán, C.; Valle, R.G.D.; Carballeira, M.N.; Sanabria-Ríos, D.J. Antibacterial fatty acids: An update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Prog. Lipid Res. 2021, 82, 101093. [Google Scholar] [CrossRef]
- Dasagrandhi, C.; Kim, Y.S.; Kim, I.H.; Hou, C.T.; Kim, H.R. 7, 10-Epoxyoctadeca-7, 9-dienoic acid: A small molecule adjuvant that potentiates β-lactam antibiotics against multidrug-resistant Staphylococcus aureus. Indian J. Microbiol. 2017, 57, 461–469. [Google Scholar] [CrossRef]
- Júnior, J.G.A.S.; Coutinho, H.D.M.; Rodrigues, J.P.V.; Ferreira, V.P.G.; Neto, J.B.A.; Silva, M.M.C.; Araújo, A.C.J.; Pereira, R.L.S.; Aquino, P.E.A.; Oliveira-Tintino, C.D.M.; et al. Liposome evaluation in inhibiting pump efflux of NorA of Staphylococcus aureus. Chem. Phys. Lipids 2022, 245, 105204. [Google Scholar] [CrossRef]
- Glen, K.A.; Lamont, I.L. β-lactam Resistance in Pseudomonas aeruginosa: Current Status, Future Prospects. Pathogens 2021, 10, 1638. [Google Scholar] [CrossRef]
- Lade, H.; Kim, J.S. Bacterial targets of antibiotics in methicillin-resistant Staphylococcus aureus. Antibiotics 2021, 10, 398. [Google Scholar] [CrossRef]
- Song, Y.; Sawa, T.; Tsuchiya, M.; Horiuchi, Y.; Kondo, S.; Hamada, M.; Umezawa, H. The screening of β-lactamase inhibitors: Inhibition by fatty acids produced by bacteria. J. Antibiot. 1981, 34, 980–983. [Google Scholar] [CrossRef] [Green Version]
- Hobby, C.R.; Herndon, J.L.; Morrow, C.A.; Peters, R.E.; Symes, S.J.; Giles, D.K. Exogenous fatty acids alter phospholipid composition, membrane permeability, capacity for biofilm formation, and antimicrobial peptide susceptibility in Klebsiella pneumoniae. Microbiologyopen 2019, 8, e00635. [Google Scholar] [CrossRef]
- Goldstein, E.J. Norfloxacin, a fluoroquinolone antibacterial agent: Classification, mechanism of action, and in vitro activity. Am. J. Med. 1987, 82, 3–17. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bacteria | Source | Profile of the Multidrug-Resistant Strains |
|---|---|---|
| E. coli EC-06 | Uroculture | Cephalothin, Cephalexin, Cefadroxil, Ceftriaxone, Cefepime, and Ampicillin + Sulbactam |
| P. aeruginosa PA-24 | Nasal | Cefepime, Ceftazidime, Imipenem, Ciprofloxacin, Piperacillin + Tazobactam, Levofloxacin, Meropenem, and Amikacin |
| S. aureus SA-10 | Rectal swab | Cephalothin, Cephalexin, Cefadroxil, Oxacillin, Penicillin, Ampicillin, Ampicillin + Sulbactam, Amoxicillin, Moxifloxacin, Ciprofloxacin, Levofloxacin, Erythromycin, Clarithromycin, Azithromycin, and Clindamycin |
| Nº | Constituents | Nº Carbons | RT (min) | AO (%) |
|---|---|---|---|---|
| 1 | Butyric acid | C4:0 | 12.34 | 1.74 |
| 2 | Caprylic acid | C8:0 | 14.37 | 8.16 |
| 3 | Capric acid | C10:0 | 17.29 | 4.85 |
| 4 | Undecanoic acid | C11:0 | 19.04 | 0.04 |
| 5 | Lauric acid | C12:0 | 20.85 | 43.01 * |
| 6 | Tridecanoic acid | C13:0 | 22.55 | 0.03 |
| 7 | Myristic acid | C14:0 | 24.37 | 11.10 * |
| 8 | Palmitic acid | C16:0 | 27.79 | 6.95* |
| 9 | Palmitoleic acid | C16:1Δ9 | 28.99 | 0.02 |
| 10 | Stearic acid | C18:0 | 31.04 | 2.55 |
| 11 | Oleic acid | C18:1Δ9 | 32.16 | 15.32 * |
| 12 | Linoleic acid | C18:2Δ9,12 | 33.77 | 1.73 |
| 13 | Arachidic acid | C20:0 | 34.26 | 0.08 |
| Total saturated | 78.51 | |||
| Total unsaturated | 17.07 | |||
| Total identified | 95.58 | |||
| Bacterial Strains | MIC (µg/mL) |
|---|---|
| Escherichia coli EC–ATCC 25922 | ≥1024 |
| Escherichia coli EC–06 | ≥1024 |
| Pseudomonas aeruginosa PA–ATCC 9027 | ≥1024 |
| Pseudomonas aeruginosa PA–24 | ≥1024 |
| Staphyloccus aureus SA–ATCC 25923 | 256 |
| Staphyloccus aureus SA–10 | ≥1024 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, C.J.F.; Raposo, A.; de Sousa, C.D.T.; de Araújo-Neto, J.B.; Tintino, S.R.; Oliveira-Tintino, C.D.d.M.; Araújo, I.M.; Coutinho, H.D.M.; Costa, M.G.M.; Lima, C.G.; et al. Modification of Antibiotic Activity by Fixed Oil of the Artocarpus heterophyllus Almond against Standard and Multidrug-Resistant Bacteria Strains. Biology 2022, 11, 1835. https://doi.org/10.3390/biology11121835
Dias CJF, Raposo A, de Sousa CDT, de Araújo-Neto JB, Tintino SR, Oliveira-Tintino CDdM, Araújo IM, Coutinho HDM, Costa MGM, Lima CG, et al. Modification of Antibiotic Activity by Fixed Oil of the Artocarpus heterophyllus Almond against Standard and Multidrug-Resistant Bacteria Strains. Biology. 2022; 11(12):1835. https://doi.org/10.3390/biology11121835
Chicago/Turabian StyleDias, Cícera Janayne Ferreira, António Raposo, Cícera Dayane Thais de Sousa, José Bezerra de Araújo-Neto, Saulo Relison Tintino, Cícera Datiane de Morais Oliveira-Tintino, Isaac Moura Araújo, Henrique Douglas Melo Coutinho, Mayra Garcia Maia Costa, Cleidiane Gomes Lima, and et al. 2022. "Modification of Antibiotic Activity by Fixed Oil of the Artocarpus heterophyllus Almond against Standard and Multidrug-Resistant Bacteria Strains" Biology 11, no. 12: 1835. https://doi.org/10.3390/biology11121835