
Adverse Effect of Blue Light on DNA Integrity Is Accelerated by 5-Aminolevulinic Acid in HaCaT Human Keratinocyte Cells and B16F1 Murine Melanoma Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. MTT Assay
2.3. Neutral Comet Assay
2.4. Alkaline Comet Assay
2.5. Two-Tailed Comet Assay (TT-Comet Assay)
2.6. Enzyme-Modified Comet Assay
2.7. Statistical Analysis
3. Results
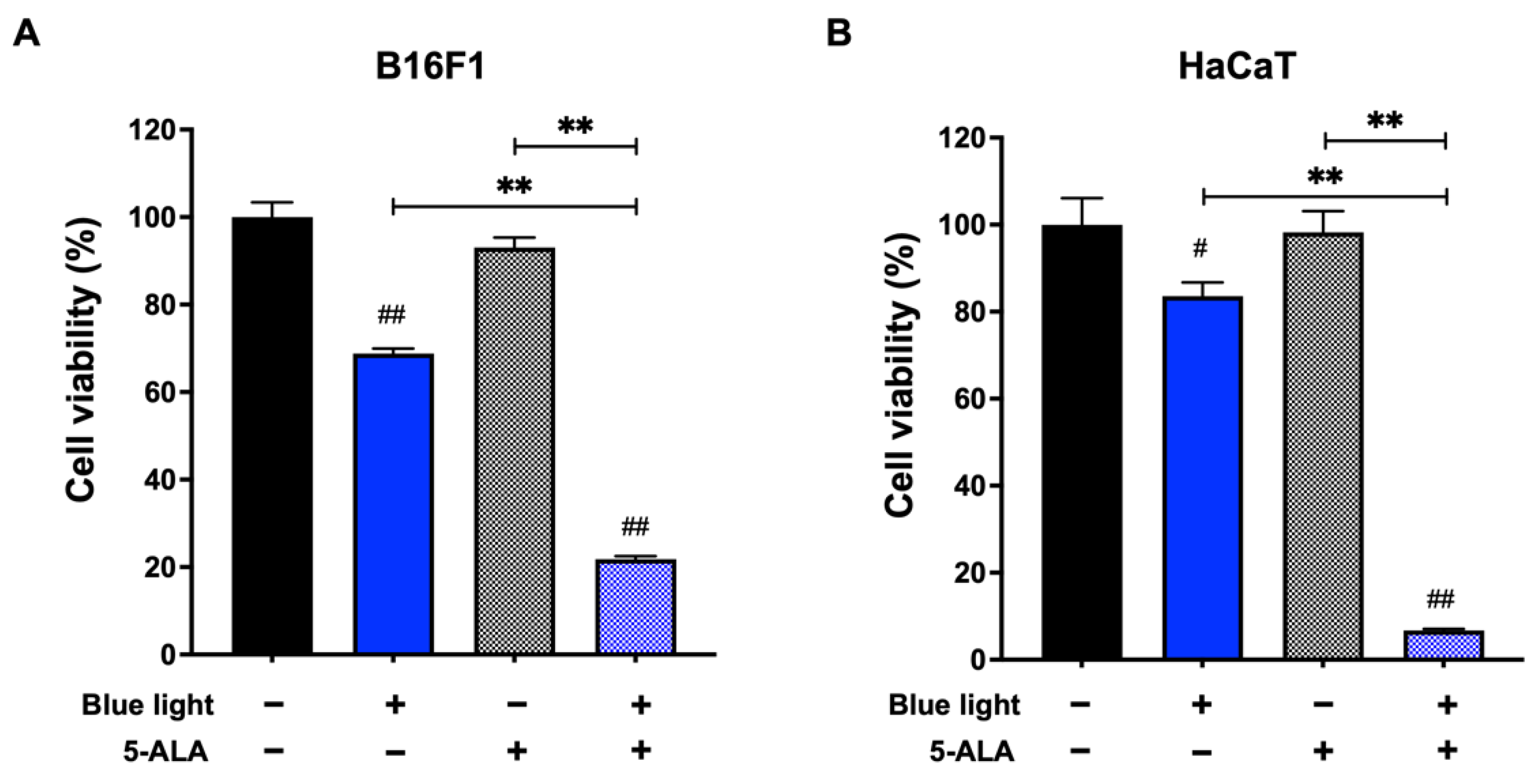
3.1. Effect of Blue Light and 5-ALA on Cell Viability
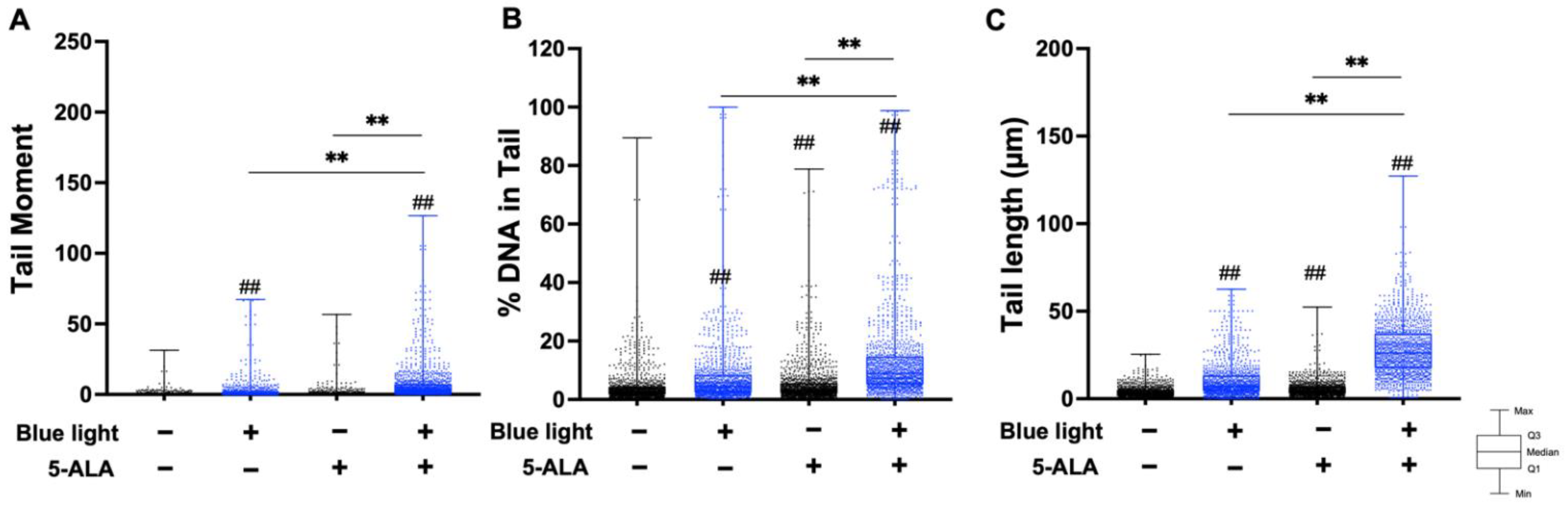
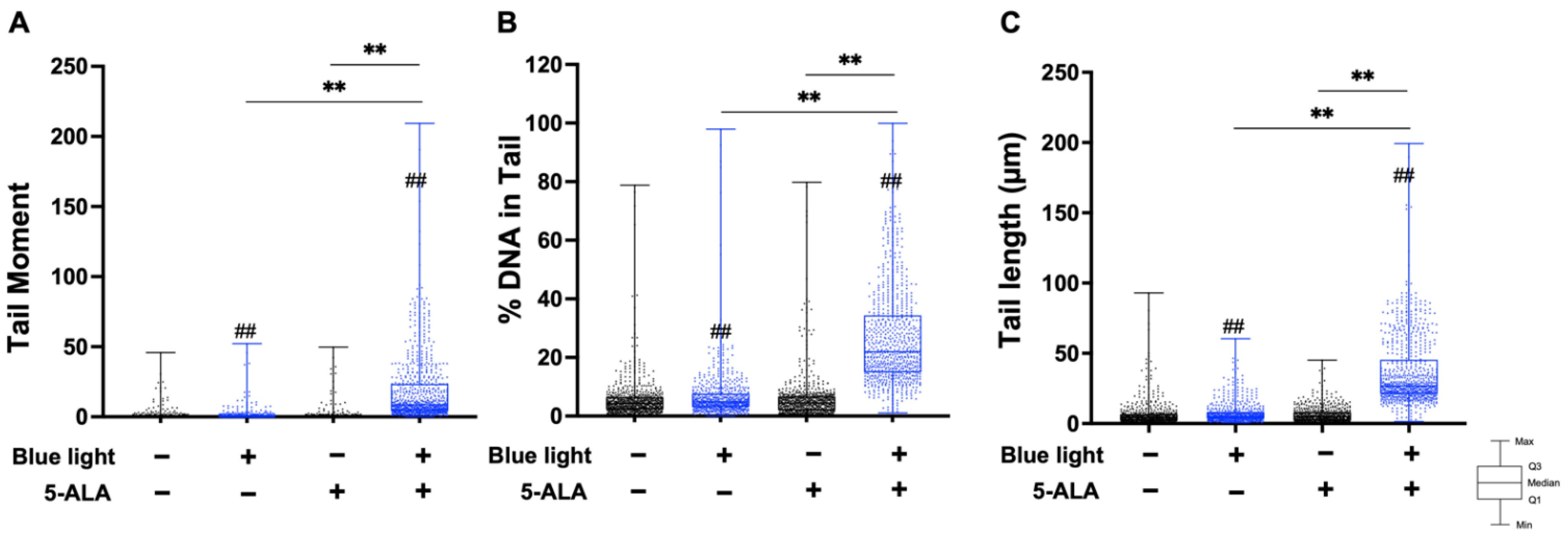
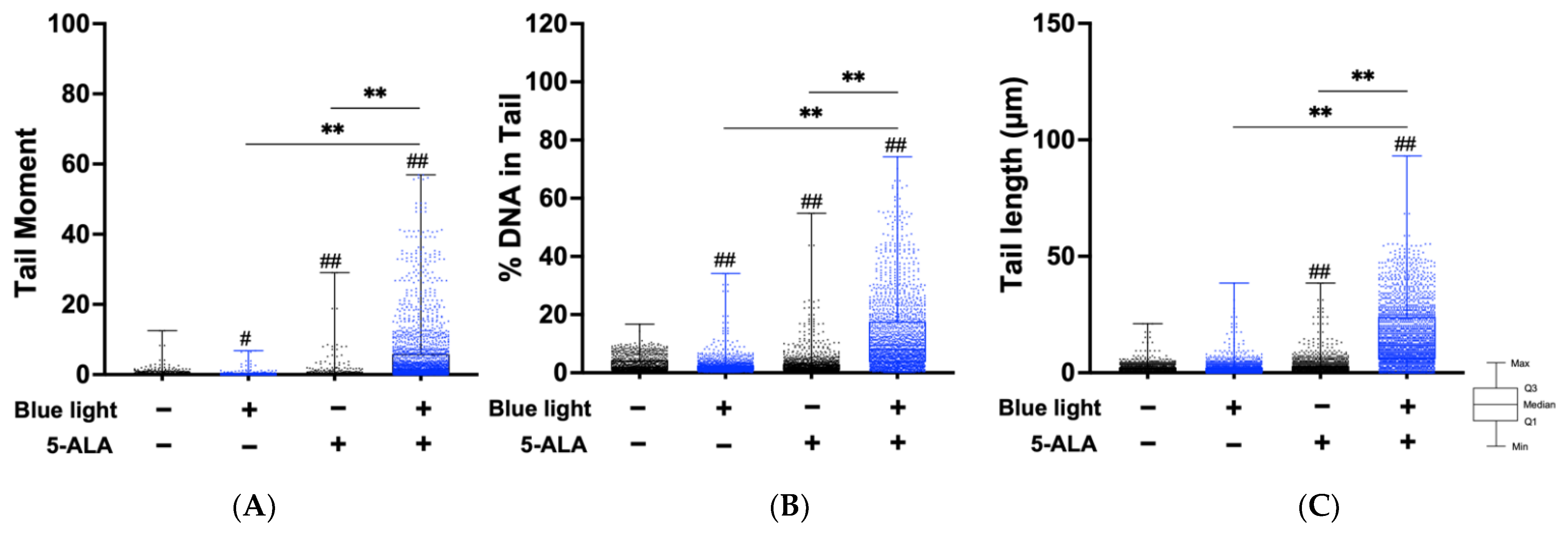
3.2. Co-Treatment with Blue Light and 5-ALA Induces DNA Strand Breaks in B16F1 and HaCaT Cells
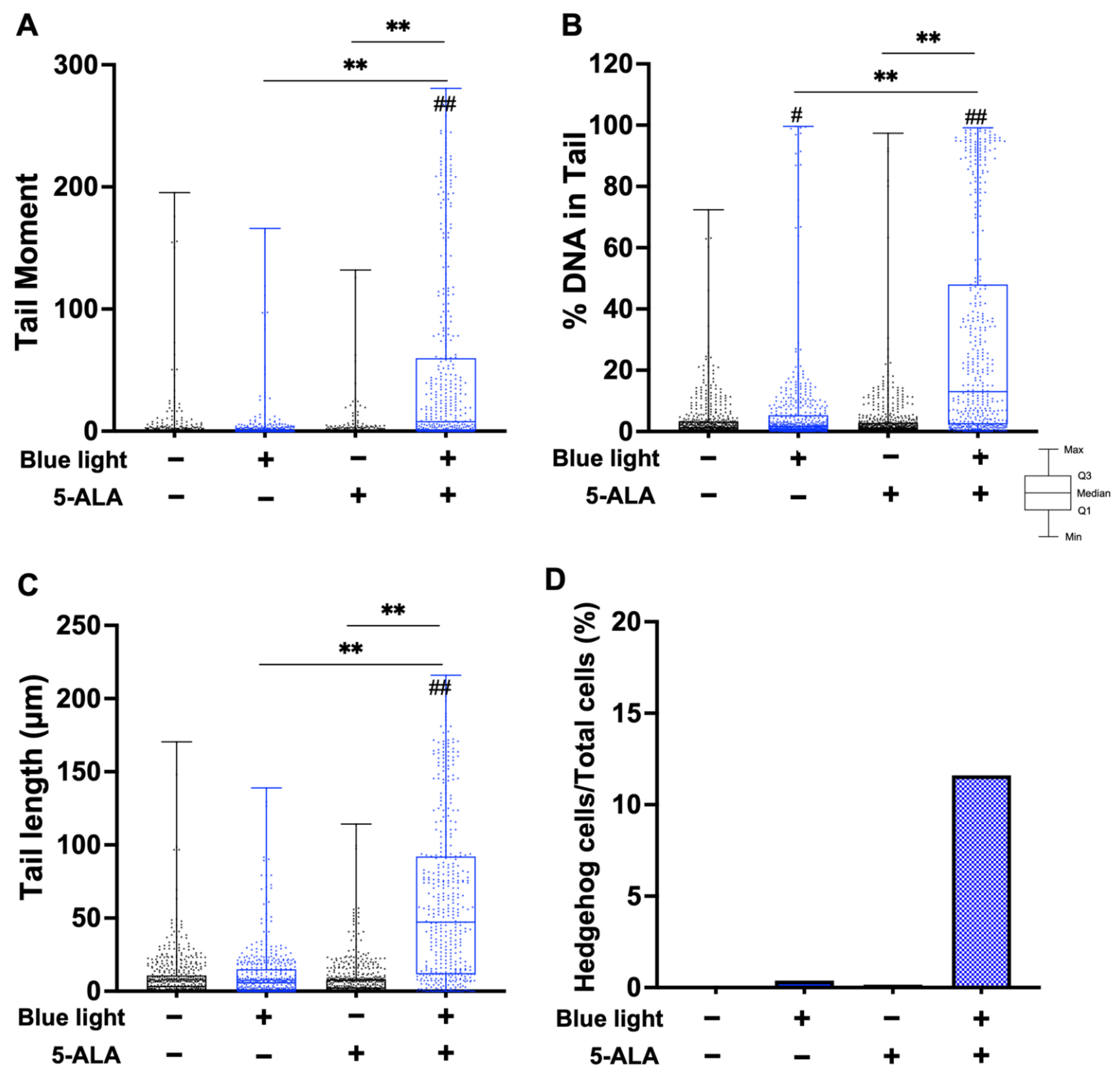
3.3. TT-Comet Assay
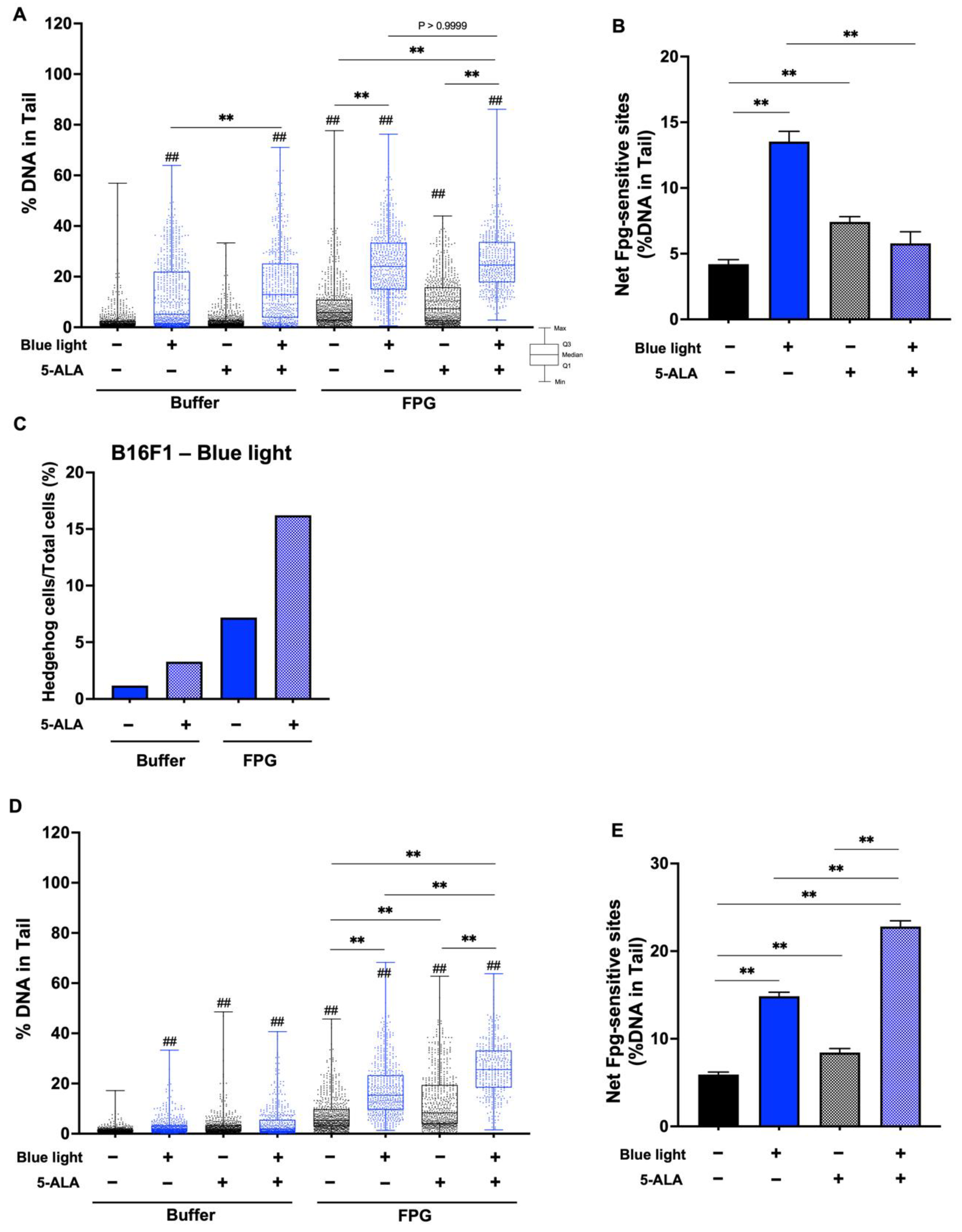
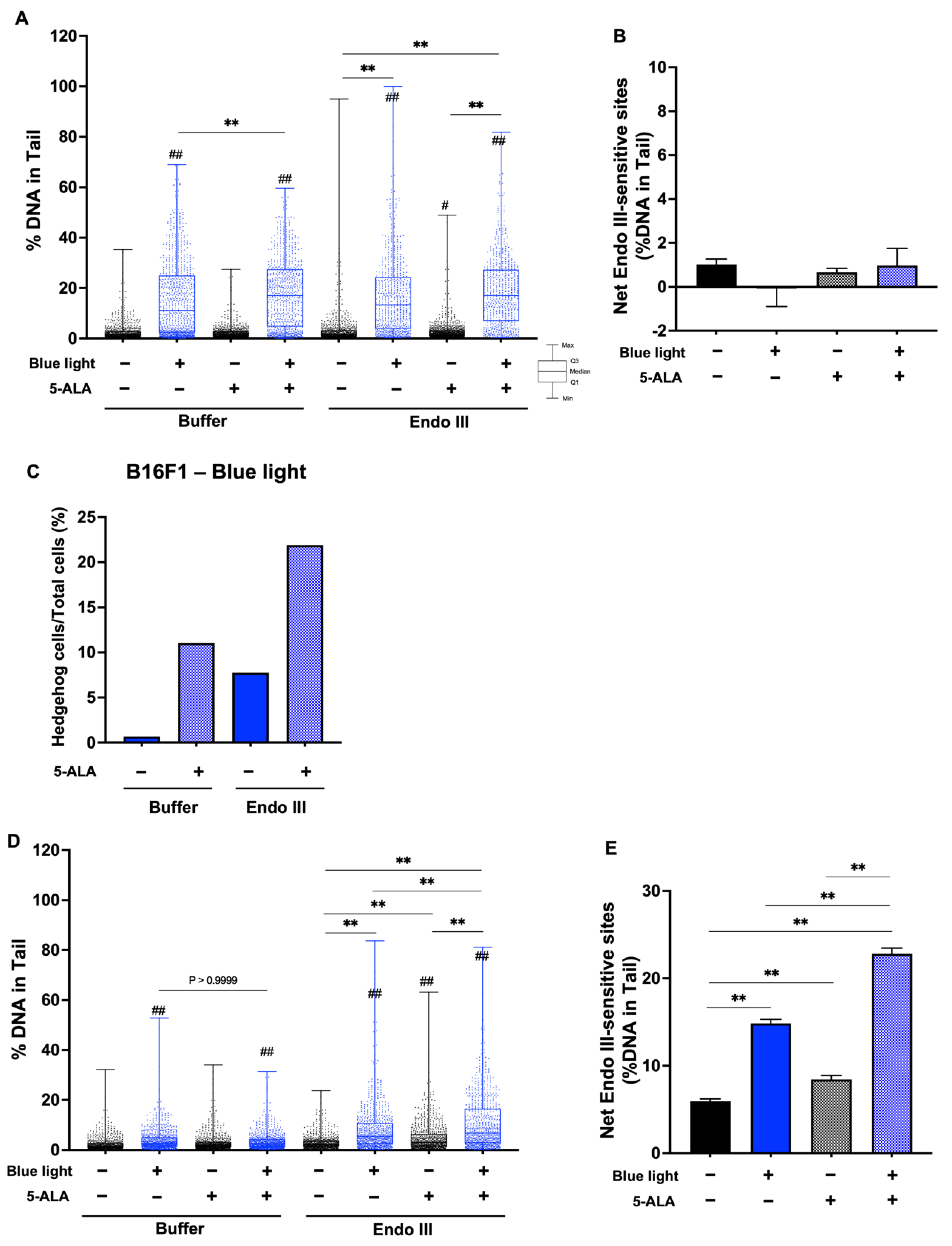
3.4. Fpg- and Endo III-Modified Comet Assay

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niaz, K.; Shah, S.Z.A.; Khan, F.; Bule, M. Ochratoxin A-induced genotoxic and epigenetic mechanism lead to Alzheimer disease: Its modulaton with strategies. Environ. Sci. Pollut. Res. 2020, 27, 44673–44700. [Google Scholar] [CrossRef]
- Muthumalage, T.; Lamb, T.; Friedman, M.R.; Rahman, I. E-cigarette flavored pods induce inflammation, epithelial barrier dysfunction, and DNA damage in lung epithelial cells and monocytes. Sci. Rep. 2019, 9, 19035. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, R.P.; Kumar, A.; Tyagi, M.B.; Sinha, R.P. Molecular mechanisms of ultraviolet radiation-induced DNA damage and repair. J. Nucleic Acids 2010, 2010, 592980. [Google Scholar] [CrossRef] [Green Version]
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet radiation-induced skin aging: The role of DNA damage and oxidative stress in epidermal stem cell damage mediated skin aging. Stem Cells Int. 2016, 2016, 7370642. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Gaddameedhi, S. Solar ultraviolet-induced DNA damage response: Melanocytes story in transformation to environmental melanomagenesis. Environ. Mol. Mutagen. 2020, 61, 736–751. [Google Scholar] [CrossRef]
- Ito, H.; Kurokawa, H.; Suzuki, H.; Indo, H.P.; Majima, H.J.; Matsui, H. 5-Aminolevulinic acid induced apoptosis via oxidative stress in normal gastric epithelial cells. J. Clin. Biochem. Nutr. 2019, 65, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Hijioka, M.; Kitamura, K.; Yanagisawa, D.; Nishimura, K.; Takata, K.; Inden, M.; Kitamura, Y. Neuroprotective effects of 5-aminolevulinic acid against neurodegeneration in rat models of parkinson’s disease and stroke. J. Pharmacol. Sci. 2020, 144, 183–187. [Google Scholar] [CrossRef]
- Islam, M.A.; Noguchi, Y.; Taniguchi, S.; Yonekura, S. Protective effects of 5-aminolevulinic acid on heat stress in bovine mammary epithelial cells. Anim. Biosci. 2021, 34, 1006–1013. [Google Scholar] [CrossRef]
- Harada, Y.; Murayama, Y.; Takamatsu, T.; Otsuji, E.; Tanaka, H. 5-aminolevulinic acid-induced protoporphyrin IX fluorescence imaging for tumor detection: Recent advances and challenges. Int. J. Mol. Sci. 2022, 23, 6478. [Google Scholar] [CrossRef]
- Sato, K.; Minai, Y.; Watanabe, H. Effect of monochromatic visible light on intracellular superoxide anion production and mitochondrial membrane potential of B16F1 and B16F10 murine melanoma cells. Cell Biol. Int. 2013, 37, 633–637. [Google Scholar] [CrossRef]
- Maytin, E.V.; Kaw, U.; Ilyas, M.; Mack, J.A.; Hu, B. Blue light versus red light for photodynamic therapy of basal cell carcinoma in patients with Gorlin syndrome: A bilaterally controlled comparison study. Photodiagn. Photodyn. Ther. 2018, 22, 7–13. [Google Scholar] [CrossRef]
- Helander, L.; Krokan, H.E.; Johnsson, A.; Gederaas, O.A.; Plaetzer, K. Red versus blue light illumination in hexyl 5-aminolevulinate photodynamic therapy: The influence of light color and irradiance on the treatment outcome in vitro. J. Biomed. Opt. 2018, 19, 088002. [Google Scholar] [CrossRef] [Green Version]
- Szeto, Y.T.; Collins, A.R.; Benzie, I.F.F. Effects of dietary antioxidants on DNA damage in lysed cells using a modified comet assay procedure. Mutat. Res. 2002, 500, 31–38. [Google Scholar] [CrossRef]
- Ghassemi-Barghi, N.; Varshosaz, J.; Etebari, M.; Dehkordi, A.J. Role of recombinant human erythropoietin loading chitosan-tripolyphosphate nanoparticles in busulfan-induced genotoxicity: Analysis of DNA fragmentation via comet assay in cultured HepG2 cells. Toxicol. In Vitro 2016, 36, 46–52. [Google Scholar] [CrossRef]
- Azqueta, A.; Slyskova, J.; Langie, S.A.S.; Gaivão, I.O.; Collins, A. Comet assay to measure DNA repair: Approach and applications. Front. Genet. 2014, 5, 288. [Google Scholar] [CrossRef] [Green Version]
- Cascajosa-Lira, A.; Puerto, M.; Prieto, A.I.; Pichardo, S.; Jiménez, L.D.-Q.; Baños, A.; Guillamón, E.; Moyano, R.; Molina-Hernández, V.; Jos, Á.; et al. Genotoxicity evaluation of Propyl-Propane-Thiosulfinate (PTS) from Allium genus essential oils by a combination of micronucleus and comet assays in rats. Foods 2021, 10, 989. [Google Scholar] [CrossRef]
- Enciso, M.; Sarasa, J.; Agarwal, A.; Fernández, J.L.; Gosálvez, J. A two-tailed comet assay for assessing DNA damage in spermatozoa. Reprod. Biomed. Online 2009, 18, 609–616. [Google Scholar] [CrossRef]
- Gyori, B.M.; Venkatachalam, G.; Thiagarajan, P.S.; Hsu, D.; Clement, M.-V. OpenComet: An automated tool for comet assay image analysis. Redox Biol. 2014, 2, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Azqueta, A.; Collins, A.R. The essential comet assay: A comprehensive guide to measuring DNA damage and repair. Arch. Toxicol. 2013, 87, 949–968. [Google Scholar] [CrossRef]
- Palazzese, L.; Gosalvez, J.; Anzalone, D.A.; Loi, P.; Saragusty, J. DNA fragmentation in epididymal freeze-dried ram spermatozoa impairs embryo development. J. Reprod. Dev. 2018, 64, 393–400. [Google Scholar] [CrossRef]
- Agarwal, A.; Barbărosie, C.; Ambar, R.; Finelli, R. The impact of single- and double-strand DNA breaks in human spermatozoa on assisted reproduction. Int. J. Mol. Sci. 2020, 21, 3882. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.M.; Al-Khalifa, M.K.; Al-Hamamah, M.A.; Alotaibi, M.R.; Attia, M.S.M.; Ahmad, S.F.; Ansari, M.A.A.; Nadeem, A.; Bakheet, S.A.B. Vorinostat is genotoxic and epigenotoxic in the mouse bone marrow cells at the human equivalent doses. Toxicology 2020, 441, 152507. [Google Scholar] [CrossRef] [PubMed]
- Dorier, M.; Tisseyre, C.; Dussert, F.; Béal, D.; Arnal, M.-E.; Douki, T.; Valdiglesias, V.; Laffon, B.; Fraga, S.; Brandão, F.; et al. Toxicological impact of acute exposure to E171 food additive and TiO2 nanoparticles on a co-culture of CaCO-2 and HT29-MTX intestinal cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 845, 402980. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, Y.; Costa, S.; Collins, A.R.; Azqueta, A. The comet assay, DNA damage, DNA repair and cytotoxicity: Hedgehogs are not always dead. Mutagenesis 2013, 28, 427–432. [Google Scholar] [CrossRef]
- Hobbs, C.A.; Recio, L.; Streicker, M.; Boyle, M.H.; Tanaka, J.; Shiga, A.; Witt, K.L. Comet assay evaluation of six chemicals of known genotoxic potential in rats. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 786–788, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, S.Y.; Cha, H.-J.; Molagoda, I.M.N.; Kim, M.Y.; Kim, S.Y.; Hwangbo, H.; Lee, H.; Kim, G.-Y.; Kim, D.-H.; Hyun, J.W.; et al. Suppression of lipopolysaccharide-induced inflammatory and oxidative response by 5-aminolevulinic acid in RAW 264.7 macrophages and zebrafish Larvae. Biomol. Ther. 2021, 29, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Copp, M.E.; Chubinskaya, S.; Bracey, D.N.; Shine, J.; Sessions, G.; Loeser, R.F.; Diekman, B.O. Comet assay for quantification of the increased DNA damage burden in primary human chondrocytes with aging and osteoarthritis. Aging Cell 2022, 21, e13698. [Google Scholar] [CrossRef]
- Saitoh, S.; Okano, S.; Nohara, H.; Nakano, H.; Shirasawa, N.; Naito, A.; Yamamoto, M.; Kelly, V.P.; Takahashi, K.; Tanaka, T.; et al. 5-aminolevulinic acid (ALA) deficiency causes impaired glucose tolerance and insulin resistance coincident with an attenuation of mitochondrial function in aged mice. PLoS ONE 2018, 13, e0189593. [Google Scholar] [CrossRef] [Green Version]
- Kamei, Y.; Hatazawa, Y.; Uchitomi, R.; Yoshimura, R.; Miura, S. Regulation of skeletal muscle function by amino acids. Nutrients 2020, 12, 261. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Hu, X.; Zhou, L.; He, Y.; Zhang, X.; Yang, J.; Ju, Z.; Liou, Y.-C.; Shen, H.-M.; Luo, G.; et al. Photodynamic therapy accelerates skin wound healing through promoting re-epithelialization. Burns Trauma 2021, 9, tkab008. [Google Scholar] [CrossRef]
- Morokuma, Y.; Yamazaki, M.; Maeda, T.; Yoshino, I.; Ishizuka, M.; Tanaka, T.; Ito, Y.; Tsuboi, R. Hair growth stimulatory effect by a combination of 5-aminolevulinic acid and iron ion. Int. J. Dermatol. 2008, 47, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Breusing, N.; Grimm, S.; Mvondo, D.; Flaccus, A.; Biesalski, H.K.; Grune, T. Light-induced cytotoxicity after aminolevulinic acid treatment is mediated by heme and not by iron. J. Photochem. Photobiol. B. 2010, 99, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, P.; Liu, J.; Du, P.; Wang, Z.; Chen, W.; Liu, C.; Wu, Y. 5-Aminolevulinic acid-based photodynamic therapy restrains pathological hyperplasia of fibroblast. Med. Sci. Monit. 2017, 23, 46–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, S.; Rollakanti, K.R.; Brankov, N.; Brash, D.E.; Hasan, T.; Maytin, E.V. Fluorouracil enhances photodynamic therapy of squamous cell carcinoma via a p53-independent mechanism that incrases protoporphyrin IX levels and tumor cell death. Mol. Cancer Ther. 2017, 16, 1092–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sznarkowska, A.; Maleńczyk, K.; Kadziński, L.; Bielawski, K.P.; Banecki, B.; Zawacka-Pankau, J. Targeting of p53 and its homolog p73 by protoporphyrin IX. FEBS Lett. 2011, 585, 255–260. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, T.; Sato, K. Adverse Effect of Blue Light on DNA Integrity Is Accelerated by 5-Aminolevulinic Acid in HaCaT Human Keratinocyte Cells and B16F1 Murine Melanoma Cells. Biology 2022, 11, 1743. https://doi.org/10.3390/biology11121743
Sato T, Sato K. Adverse Effect of Blue Light on DNA Integrity Is Accelerated by 5-Aminolevulinic Acid in HaCaT Human Keratinocyte Cells and B16F1 Murine Melanoma Cells. Biology. 2022; 11(12):1743. https://doi.org/10.3390/biology11121743
Chicago/Turabian StyleSato, Taiki, and Kazuomi Sato. 2022. "Adverse Effect of Blue Light on DNA Integrity Is Accelerated by 5-Aminolevulinic Acid in HaCaT Human Keratinocyte Cells and B16F1 Murine Melanoma Cells" Biology 11, no. 12: 1743. https://doi.org/10.3390/biology11121743