
Butterfly Wing Color Pattern Modification Inducers May Act on Chitin in the Apical Extracellular Site: Implications in Morphogenic Signals for Color Pattern Determination

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Butterflies
2.2. Mechanical Load Tests
2.3. Chemical Injections
2.4. Covering Material Experiments
2.5. Developmental Imaging
2.6. Statistical Analysis
3. Results
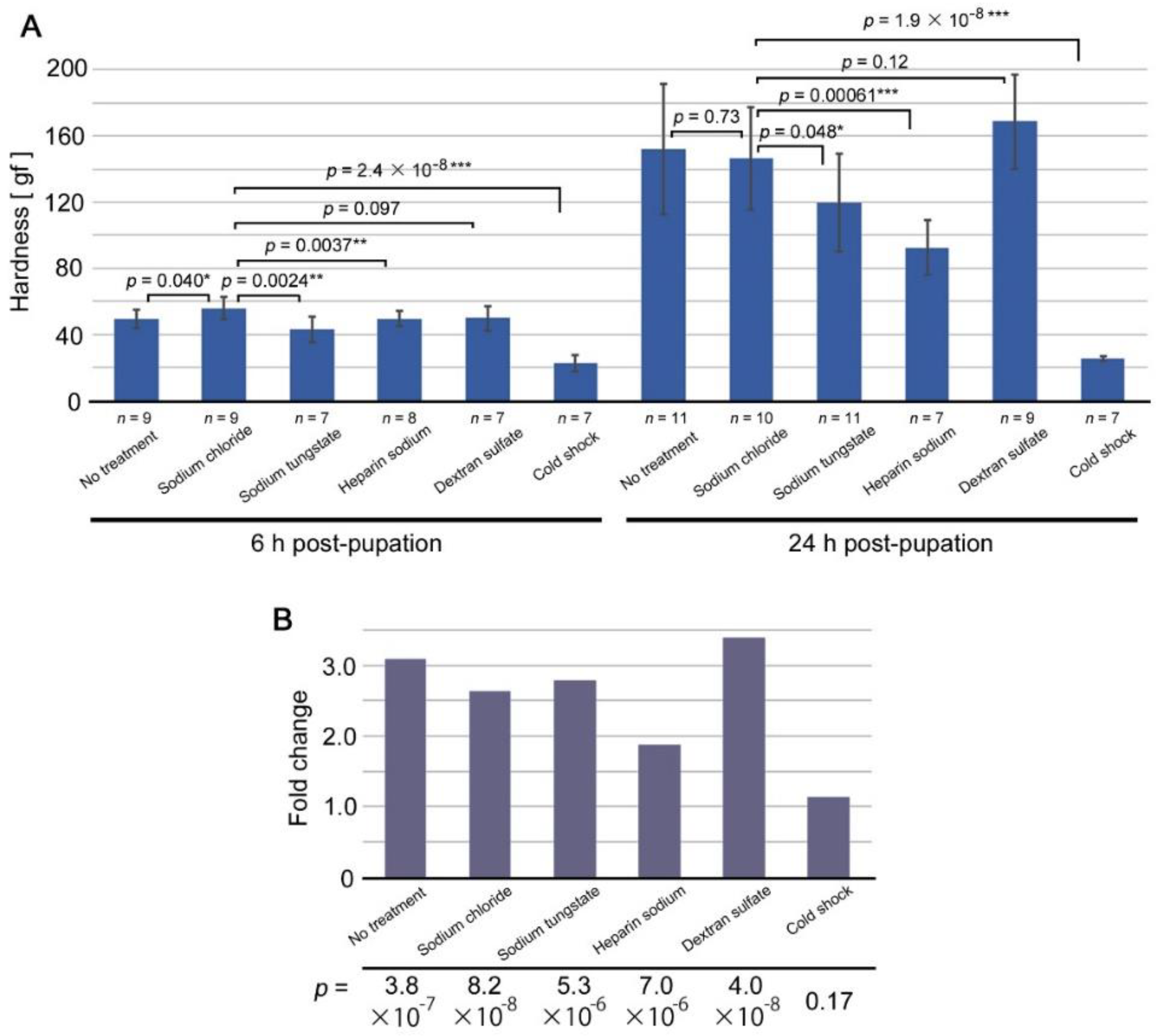
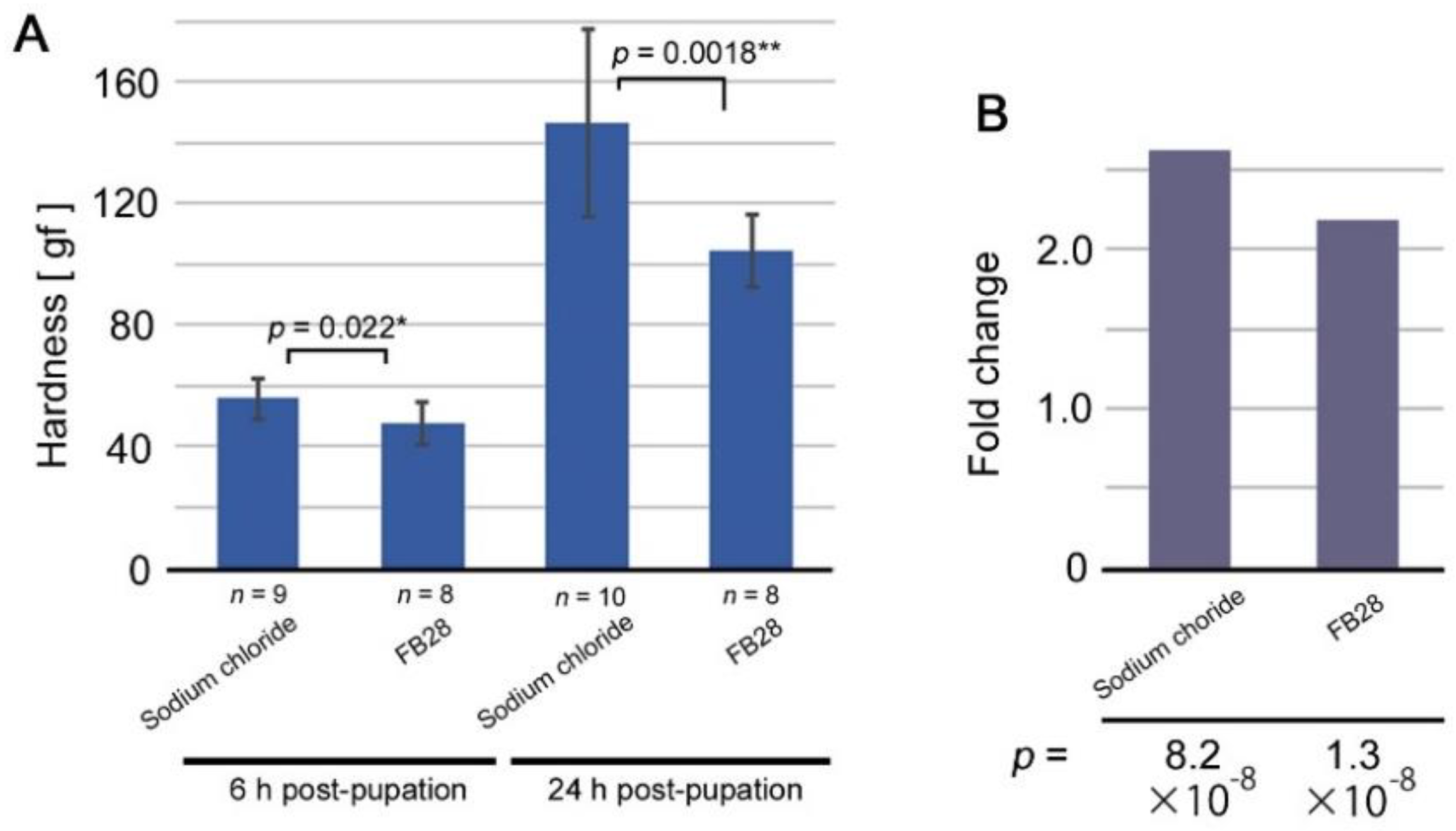
3.1. Mechanical Load Test for Cuticle Hardness
3.2. Inhibitors of Cuticle Formation Tested: Discovery of FB28 as a Modification Inducer
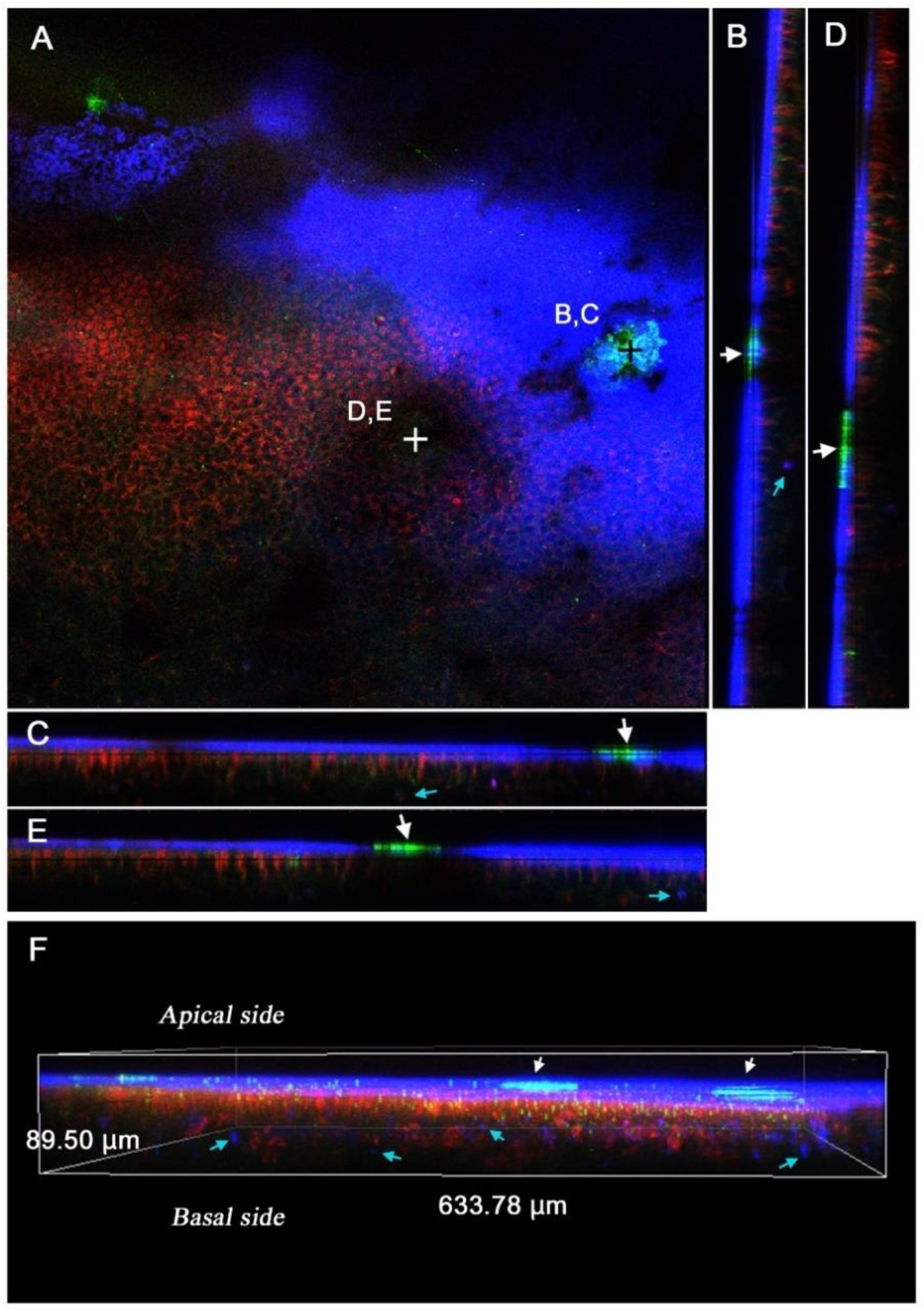
3.3. Sites of FB28 Staining
3.4. FB28 on the Pupal and Adult Cuticle Structures
3.5. Covering Materials and FB28
3.6. Covering Materials and Dextran Sulfate
4. Discussion
4.1. The Pupal Cuticle Hypothesis
4.2. Chitin: The Potential Site of Action of Various Modification Inducers
4.3. Chitin and Morphogenic Signals
4.4. Behaviors of Eyespots and Parafocal Elements
4.5. Chitin-Related Molecules and Transcription Factors
4.6. Molecular (Chemical) Morphogens and Mechanical (Physical) Morphogens
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nijhout, H.F. The Development and Evolution of Butterfly Wing Patterns; Smithsonian Institution Press: Washington, DC, USA, 1991. [Google Scholar]
- Nijhout, H.F. Elements of butterfly wing patterns. J. Exp. Zool. 2001, 291, 213–225. [Google Scholar] [CrossRef]
- Otaki, J.M. Color pattern analysis of nymphalid butterfly wings: Revision of the nymphalid groundplan. Zool. Sci. 2012, 29, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Otaki, J.M. Morphological and spatial diversity of the discal spot on the hindwings of nymphalid butterflies: Revision of the nymphalid groundplan. Insects 2020, 11, 654. [Google Scholar] [CrossRef] [PubMed]
- Otaki, J.M. The fractal geometry of the nymphalid groundplan: Self-similar configuration of color pattern symmetry systems in butterfly wings. Insects 2021, 12, 39. [Google Scholar] [CrossRef] [PubMed]
- Otaki, J.M. Self-similarity, distortion waves, and the essence of morphogenesis: A generalized view of color pattern formation in butterfly wings. In Diversity and Evolution of Butterfly Wing Patterns: An Integrated Approach; Sekimura, T., Nijhout, H.F., Eds.; Springer: Singapore, 2017; pp. 119–152. [Google Scholar] [CrossRef]
- Carroll, S.B.; Gates, J.; Keys, D.N.; Paddock, S.W.; Panganiban, G.E.; Selegue, J.E.; Williams, J.A. Pattern formation and eyespots determination in butterfly wings. Science 1994, 265, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Brakefield, P.M.; Gates, J.; Keys, D.; Kesbeke, F.; Wijngaarden, P.J.; Monteiro, A.; French, V.; Carroll, S.B. Development, plasticity and evolution of butterfly eyespot patterns. Nature 1996, 384, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Keys, D.N.; Lewis, D.L.; Selegue, J.E.; Pearson, B.J.; Goodrich, L.V.; Johnson, R.L.; Gates, J.; Scott, M.P.; Carroll, S.B. Recruitment of a hedgehog regulatory circuit in butterfly eyespot evolution. Science 1999, 283, 532–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, A.; Glaser, G.; Stockslager, S.; Glansdorp, N.; Ramos, D. Comparative insights into questions of lepidopteran wing pattern homology. BMC Dev. Biol. 2006, 6, 52. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.; Papa, R.; Nadeau, N.J.; Hill, R.I.; Counterman, B.A.; Halder, G.; Jiggins, C.D.; Kronforst, M.R.; Long, A.D.; McMillan, W.O.; et al. Diversification of complex butterfly wing patterns by repeated regulatory evolution of a Wnt ligand. Proc. Natl. Acad. Sci. USA 2012, 109, 12632–12637. [Google Scholar] [CrossRef]
- Monteiro, A.; Chen, B.; Ramos, D.M.; Oliver, J.C.; Tong, X.; Guo, M.; Wang, W.-K.; Fazzino, L.; Kamal, F. Distal-less regulates eyespot patterns and melanization in Bicyclus butterflies. J. Exp. Zool. B Mol. Dev. Evol. 2013, 320, 321–331. [Google Scholar] [CrossRef]
- Martin, A.; Reed, R.D. Wnt signaling underlies evolution and development of the butterfly wing pattern symmetry systems. Dev. Biol. 2014, 395, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, K.; Otaki, J.M. A single-wing removal method to assess correspondence between gene expression and phenotype in butterflies. The case of Distal-less. Zool. Sci. 2016, 33, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, B.; Ohno, Y.; Matayoshi, R.; Iwasaki, M.; Taira, W.; Adhikari, K.; Gurung, R.; Otaki, J.M. Distal-less induces elemental color patterns in Junonia butterfly wings. Zool. Lett. 2016, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Reed, R.D. Genome editing in butterflies reveals that spalt promotes and Distal-less represses eyespot colour patterns. Nat. Commun. 2016, 7, 11769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özsu, N.; Monteiro, A. Wound healing, calcium signaling, and other novel pathways are associated with the formation of butterfly eyespots. BMC Genom. 2017, 18, 788. [Google Scholar] [CrossRef] [Green Version]
- Özsu, N.; Chan, Q.Y.; Chen, B.; Gupta, M.D.; Monteiro, A. Wingless is a positive regulator of eyespot color patterns in Bicyclus anynana butterflies. Dev. Biol. 2017, 429, 177–185. [Google Scholar] [CrossRef]
- Mazo-Vargas, A.; Concha, C.; Livraghi, L.; Massardo, D.; Wallbank, R.W.R.; Zhang, L.; Papador, J.D.; Martinez-Najera, D.; Jiggins, C.D.; Kronforst, M.R.; et al. Macroevolutionary shifts of WntA function potentiate butterfly wing-pattern diversity. Proc. Natl. Acad. Sci. USA 2017, 114, 10701–10706. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Mazo-Vargas, A.; Reed, R.D. Single master regulatory gene coordinates the evolution and development of butterfly color and iridescence. Proc. Natl. Acad. Sci. USA 2017, 114, 10707–10712. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Martin, A.; Perry, M.W.; van der Burg, K.R.; Matsuoka, Y.; Monteiro, A.; Reed, R.D. Genetic basis of melanin pigmentation in butterfly wings. Genetics 2017, 205, 1537–1550. [Google Scholar] [CrossRef]
- Connahs, H.; Tlili, S.; van Creij, J.; Loo, T.Y.J.; Banerjee, T.D.; Saunders, T.E.; Monteiro, A. Activation of butterfly eyespots by Distal-less is consistent with a reaction-diffusion process. Development 2019, 146, dev169367. [Google Scholar] [CrossRef] [Green Version]
- Westerman, E.L.; VanKuren, N.M.; Massardo, D.; Tenger-Trolander, A.; Zhang, W.; Hill, R.I.; Perry, M.; Bayala, E.; Barr, K.; Chamberian, N.; et al. Aristaless controls butterfly wing color variation used in mimicry and mate choice. Curr. Biol. 2018, 28, 3469–3474. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, Y.; Monteiro, A. Melanin pathway genes regulate color and morphology of butterfly wing scales. Cell Rep. 2018, 24, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, A.; Monteiro, A. apterous A specifies dorsal wing patterns and sexual traits in butterflies. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, R.D.; Selegue, J.E.; Zhang, L.; Brunetti, C.R. Transcription factors underlying wing margin color patterns and pupal cuticle markings in butterflies. Evodevo 2020, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.L.; Mazo-Vargas, A.; Brack, B.J.; Reed, R.D. Multiple roles for laccase2 in butterfly wing pigmentation, scale development, and cuticle tanning. Evol. Dev. 2020, 22, 336–341. [Google Scholar] [CrossRef]
- van der Burg, K.R.; Lewis, J.J.; Brack, B.J.; Fandino, R.A.; Mazo-Vargas, A.; Reed, R.D. Genomic architecture of a genetically assimilated seasonal color pattern. Science 2020, 370, 721–725. [Google Scholar] [CrossRef]
- Nijhout, H.F. Colour pattern modification by cold-shock in Lepidoptera. J. Embryol. Exp. Morphol. 1984, 81, 287–305. [Google Scholar]
- Otaki, J.M. Colour-pattern modifications of butterfly wings induced by transfusion and oxyanions. J. Insect Physiol. 1998, 44, 1181–1190. [Google Scholar] [CrossRef]
- Otaki, J.M. Physiologically induced colour-pattern changes in butterfly wings: Mechanistic and evolutionary implications. J. Insect Physiol. 2008, 54, 1099–1112. [Google Scholar] [CrossRef]
- Otaki, J.M.; Ogasawara, T.; Yamamoto, H. Tungstate-induced color-pattern modifications of butterfly wings are independent of stress response and ecdysteroid effect. Zool. Sci. 2005, 22, 635–644. [Google Scholar] [CrossRef]
- Umebachi, Y.; Osanai, M. Perturbation of the wing color pattern of a swallowtail butterfly, Papilio xuthus, induced by acid carboxypeptidase. Zool. Sci. 2003, 20, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serfas, M.S.; Carroll, S.B. Pharmacologic approaches to butterfly wing patterning: Sulfated polysaccharides mimic or antagonize cold shock and alter the interpretation of gradients of positional information. Dev. Biol. 2005, 287, 416–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otaki, J.M. Reversed type of color-pattern modifications of butterfly wings: A physiological mechanism of wing-wide color-pattern determination. J. Insect Physiol. 2007, 53, 256–537. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, S.H.A.; Yamasaki, H.; Otaki, J.M. Heat-shock-induced color-pattern changes of the blue pansy butterfly Junonia orithya. J. Therm. Biol. 2011, 36, 312–321. [Google Scholar] [CrossRef]
- Mahdi, S.H.A.; Gima, S.; Tomita, Y.; Yamasaki, H.; Otaki, J.M. Physiological characterization of the cold-shock-induced humoral factor for wing color-pattern changes in butterflies. J. Insect Physiol. 2010, 56, 1022–1031. [Google Scholar] [CrossRef]
- Otaki, J.M. Asymmetrical color pattern of Vanessa cardui: A case report of a field-caught individual and experimental pattern modification. Butterflies 2003, 35, 50–56. [Google Scholar]
- Dhungel, B.; Otaki, J.M. Local pharmacological effects of tungstate on the color-pattern determination of butterfly wings: A possible relationship between the eyespot and parafocal element. Zool. Sci. 2009, 26, 758–764. [Google Scholar] [CrossRef] [Green Version]
- Otaki, J.M. Contact-mediated eyespot color-pattern changes in the peacock pansy butterfly: Contributions of mechanical force and extracellular matrix to morphogenic signal propagation. In Lepidoptera; Perveen, F.K., Ed.; InTech: Rijeka, Croatia, 2017; pp. 83–102. [Google Scholar] [CrossRef] [Green Version]
- Otaki, J.M. Butterfly eyespot color pattern formation requires physical contact of the pupal wing epithelium with extracellular materials for morphogenic signal propagation. BMC Dev. Biol. 2020, 20, 6. [Google Scholar] [CrossRef] [Green Version]
- Otaki, J.M.; Ogasawara, T.; Yamamoto, H. Morphological comparison of pupal wing cuticle patterns in butterflies. Zool. Sci. 2005, 22, 21–34. [Google Scholar] [CrossRef]
- Harrington, B.J.; Raper, K.B. Use of a fluorescent brightener to demonstrate cellulose in the cellular slime molds. Appl. Microbiol. 1968, 16, 106–113. [Google Scholar] [CrossRef]
- Herth, W.; Schnepf, E. The fluorochrome, Calcofluor white, binds oriented to structural polysaccharide fibrils. Protoplasma 1980, 105, 129–133. [Google Scholar] [CrossRef]
- Shapiro, M.; Robertson, J.L. Enhancement of gypsy moth (Lepidoptera: Lymantriidae) baculovirus activity by optical brighteners. J. Econ. Entomol. 1992, 85, 1120–1124. [Google Scholar] [CrossRef]
- Hamm, J.J.; Shapiro, M. Infectivity of fall armyworm (Lepidopterra: Noctuidae) nuclear polyhedrosis virus enhanced by a fluorescent brightener. J. Econ. Entomol. 1992, 85, 2149–2152. [Google Scholar] [CrossRef]
- Shapiro, M.; Shepard, B.M. Relative efficacies of congo red and Tinopal LPW on the activity of the gypsy moth (Lepidoptera: Lymantriidae), nucleopolyhedrovirus and cypovirus. J. Agric. Urban Entomol. 2008, 25, 233–243. [Google Scholar] [CrossRef]
- Arakawa, T.; Kamimura, M.; Furuta, Y.; Miyazawa, M.; Kato, M. Peroral infection of nuclear polyhedrosis virus budded particles in the host, Bombyx mori L., enabled by an optical brightener, Tinopal UNPA-GX. J. Virol. Methods 2000, 88, 145–152. [Google Scholar] [CrossRef]
- Flaven-Pouchon, J.; Moussian, B. Fluorescent microscopy-based detection of chitin in intact Drosophila melanogaster. Front. Physiol. 2022, 13, 856369. [Google Scholar] [CrossRef]
- Arakawa, T. Chitin synthesis inhibiting antifungal agents promote nucleopolyhedrovirus infection in silkworm, Bombyx mori (Lepidoptera: Bombycidae) larvae. J. Invertebr. Pathol. 2003, 83, 261–263. [Google Scholar] [CrossRef]
- Guo, H.F.; Fang, J.C.; Liu, B.S.; Wang, J.P.; Zhong, W.F.; Wan, F.H. Enhancement of biological activity of nucleopolyhedrovirus through disruption of the peritrophic matrix of insect larvae by chlorfluazuron. Pest Manag. Sci. 2007, 63, 68–74. [Google Scholar] [CrossRef]
- Hori, M.; Kakiki, K.; Suzuki, S.; Misato, T. Studies on the mode of action of polyoxyins. Part III. Relation of polyoxyin structure to chitin synthetase inhibition. Agric. Biol. Chem. 1971, 35, 1280–1291. [Google Scholar] [CrossRef]
- Tada, M.; Matsumoto, Y.; Mitsui, T.; Nobusawa, C.; Fukami, J. Inhibition of chitin synthesis by 1-(3,5-dichloro-2,4-difluorophenyl)-3-(2,6-difluorobenzoyl)urea (CME-134) in the cabbage armyworm, Mamestra brassicae L. J. Pestic. Sci. 1986, 11, 189–195. [Google Scholar] [CrossRef]
- Arakawa, T.; Yukuhiro, F.; Noda, H. Insecticidal effect of a fungicide containing polyoxyin B on the larvae of Bombyx mori (Lepidoptera, Bombycidae), Mamestra brassicae, Mythimna separata, and Spodoptera litura (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 2008, 43, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Kusaba, K.; Otaki, J.M. Positional dependence of scale size and shape in butterfly wings: Wing-wide phenotypic coordination of color-pattern elements and background. J. Insect Physiol. 2009, 55, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Ohno, Y.; Otaki, J.M. Real-time in vivo imaging of butterfly wing development: Revealing the cellular dynamics of the pupal wing tissue. PLoS ONE 2014, 9, e89500. [Google Scholar] [CrossRef] [Green Version]
- Ohno, Y.; Otaki, J.M. Live cell imaging of butterfly pupal and larval wings in vivo. PLoS ONE 2015, 10, e0128332. [Google Scholar] [CrossRef] [Green Version]
- Taira, W.; Otaki, J.M. Butterfly wings are three-dimensional: Pupal cuticle focal spots and their associated structures in Junonia butterflies. PLoS ONE 2016, 11, e0146348. [Google Scholar] [CrossRef] [Green Version]
- Moussian, B. Chitin: Structure, chemistry and biology. In Targeting Chitin-Containing Organisms. Advances in Experimental Medicine and Biology; Yang, Q., Fukamizo, T., Eds.; Springer: Singapore, 2019; Volume 1142, pp. 5–18. [Google Scholar] [CrossRef]
- Chaudhary, V.; Boutros, M. Exocyst-mediated apical Wg secretion activates signaling in the Drosophila wing epithelium. PLoS Genet. 2019, 15, e1008351. [Google Scholar] [CrossRef] [Green Version]
- Moussian, B.; Schwarz, H.; Bartoszewski, S.; Nüsslein-Volhard, C. Involvement of chitin in exoskeleton morphogenesis in Drosophila melanogaster. J. Morphol. 2005, 264, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Muthukrishnan, S.; Merzendorfer, H.; Arakane, Y.; Kramer, K.J. Chitin metabolism in insects. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Academic Press: New Yok, NY, USA, 2012; pp. 193–235. [Google Scholar] [CrossRef]
- Floor, M.; Schenk, K.M.; Kieboom, A.P.G.; van Bekkum, H. Oxidation of maltodextrins and starch by the system tungstate-hydrogen peroxide. Starch 1989, 41, 303–309. [Google Scholar] [CrossRef]
- Lukasiewicz, M.; Bednarz, S.; Ptaszek, A. Environmental friendly polysaccharide modification—Microwave-assisted oxidation of starch. Starch 2011, 63, 268–273. [Google Scholar] [CrossRef]
- Cumpstey, I. Chemical modification of polysaccharides. ISRN Org. Chem. 2013, 2013, 417672. [Google Scholar] [CrossRef]
- Lu, X.; Li, J.; Liu, X.; Li, X.; Ma, J. Characterization and expression analysis of six chitinase genes from the desert beetle Microdera punctipennis in response to low temperature. CryoLetters 2014, 35, 438–448. [Google Scholar]
- Binari, R.C.; Staveley, B.E.; Johnson, W.A.; Godavarti, R.; Sasisekharan, R.; Manoukian, A.S. Genetic evidence that heparin-like glycosaminoglycans are involved in wingless signaling. Development 1997, 124, 2623–2632. [Google Scholar] [CrossRef] [PubMed]
- Baeg, G.H.; Lin, X.; Khare, N.; Baumgartner, S.; Perrimon, N. Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development 2001, 128, 87–94. [Google Scholar] [CrossRef]
- Moussian, B. Recent advances in understanding mechanisms of insect cuticle differentiation. Insect Biochem. Mol. Biol. 2010, 40, 363–375. [Google Scholar] [CrossRef]
- Andersen, S.O. Insect cuticular sclerotization: A review. Insect Biochem. Mol. Biol. 2010, 40, 166–178. [Google Scholar] [CrossRef]
- Andersen, S.O. Cuticle sclerotization and tanning. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Academic Press: New Yok, NY, USA, 2012; pp. 167–192. [Google Scholar] [CrossRef]
- Baker, J.D.; Truman, J.W. Mutations in the Drosophila glycoprotein hormone receptor, rickets, eliminate neuropeptide-induced tanning and selectively block a stereotyped behavioral program. J. Exp. Biol. 2002, 205, 2555–2565. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.-W.; Dewey, E.M.; Sudo, S.; Ewer, J.; Hsu, S.Y.; Honegger, H.-W.; Hsueh, J.W. Bursicon, the insect cuticle-hardening hormone, is a heterodimeric cysteine knot protein that activates G protein-coupled receptor LGR2. Proc. Natl. Acad. Sci. USA 2005, 102, 2820–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, L.; Li, Y.; Xiong, G.; Liu, X.; He, S.; Tong, X.; Wu, S.; Hu, H.; Wang, R.; Hu, H.; et al. Effects of altered catecholamine metabolism on pigmentation and physical properties of sclerotized regions in the silkworm melanism mutant. PLoS ONE 2012, 7, e42968. [Google Scholar] [CrossRef]
- Otaki, J.M. Color-pattern analysis of eyespots in butterfly wings: A critical examination of morphogen gradient models. Zool. Sci. 2011, 28, 403–413. [Google Scholar] [CrossRef]
- Otaki, J.M. Generation of butterfly wing eyespot patterns: A model for morphological determination of eyespot and parafocal element. Zool. Sci. 2011, 28, 817–827. [Google Scholar] [CrossRef]
- Iwata, M.; Tsutsumi, M.; Otaki, J.M. Developmental dynamics of butterfly wings: Real-time in vivo whole-wing imaging of twelve butterfly species. Sci. Rep. 2018, 8, 16848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, M.; Otaki, J.M. Insights into eyespot color-pattern formation mechanisms from color gradients, boundary scales, and rudimentary eyespots in butterfly wings. J. Insect Physiol. 2019, 114, 68–82. [Google Scholar] [CrossRef]
- Ramírez-Weber, F.-A.; Kornberg, T.B. Cytonemes: Cellular processes that project to the principal signaling center in Drosophila imaginal discs. Cell 1999, 97, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, T.B. Cytonemes and the dispersion of morphogens. WIREs Dev. Biol. 2014, 3, 445–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Scholpp, S. Cytonemes in development. Curr. Opin. Genet. Dev. 2019, 57, 25–30. [Google Scholar] [CrossRef]
- Rustom, A.; Saffrich, R.; Markovic, I.; Walther, P.; Gerdes, H.H. Nanotubular highways for intercellular organelle transport. Science 2004, 303, 1007–1010. [Google Scholar] [CrossRef] [Green Version]
- Korenkova, O.; Pepe, A.; Zurzolo, C. Fine intercellular connections in development: TNTs, cytonemes, or intercellular bridges? Cell Stress 2020, 4, 30–43. [Google Scholar] [CrossRef]
- Kaupp, U.B. Olfactory signalling in vertebrates and insects: Differences and commonalities. Nat. Rev. Neurosci. 2010, 11, 188–200. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | Concentration | Treated | Modified | Survived | MR(%) | SR(%) |
|---|---|---|---|---|---|---|
| Fluorescent brightener 28 (FB28) | 25% | 12 | 0 | 0 | NA | 0 |
| 12.5% | 13 | 2 | 2 | 100 | 15 | |
| 8.3% | 6 | 2 | 2 | 100 | 33 | |
| 7.5% | 29 | 16 | 16 | 100 | 55 | |
| 6.3% | 3 | 1 | 1 | 100 | 33 | |
| 5.0% | 9 | 6 | 9 | 67 | 100 | |
| 2.5% | 18 | 5 | 16 | 31 | 89 | |
| Sodium tungstate | 1.0 M | 8 | 6 | 6 | 100 | 75 |
| Heparin sodium | 3.0 mg/mL | 10 | 7 | 7 | 100 | 70 |
| Dextran sulfate | 10.0 mg/mL | 9 | 9 | 9 | 100 | 100 |
| Chlorfluazuron | 0.0010% | 7 | 0 | 7 | 0 | 100 |
| 0.00010% | 7 | 0 | 6 | 0 | 86 | |
| Captan | 0.0010% | 15 | 0 | 8 | 0 | 53 |
| Congo red | 10.0% | 11 | 0 | 6 | 0 | 55 |
| 1.0% | 7 | 0 | 5 | 0 | 71 | |
| 0.10% | 19 | 0 | 15 | 0 | 79 | |
| Polyoxin B | 1.0 mg/mL | 10 | 0 | 9 | 0 | 90 |
| 0.10 mg/mL | 6 | 0 | 5 | 0 | 83 | |
| Polyoxin D | 1.0 mg/mL | 14 | 0 | 1 | 0 | 7 |
| 0.10 mg/mL | 8 | 0 | 8 | 0 | 100 | |
| NaCl | 1.0 M | 10 | 0 | 10 | 0 | 100 |
| No treatment | NA | 51 | 0 | 50 | 0 | 98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otaki, J.M.; Nakazato, Y. Butterfly Wing Color Pattern Modification Inducers May Act on Chitin in the Apical Extracellular Site: Implications in Morphogenic Signals for Color Pattern Determination. Biology 2022, 11, 1620. https://doi.org/10.3390/biology11111620
Otaki JM, Nakazato Y. Butterfly Wing Color Pattern Modification Inducers May Act on Chitin in the Apical Extracellular Site: Implications in Morphogenic Signals for Color Pattern Determination. Biology. 2022; 11(11):1620. https://doi.org/10.3390/biology11111620
Chicago/Turabian StyleOtaki, Joji M., and Yugo Nakazato. 2022. "Butterfly Wing Color Pattern Modification Inducers May Act on Chitin in the Apical Extracellular Site: Implications in Morphogenic Signals for Color Pattern Determination" Biology 11, no. 11: 1620. https://doi.org/10.3390/biology11111620