
A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete
Siena College, Department of Biological Sciences, Loudonville, NY 12211, USA
Biology 2022, 11(10), 1485; https://doi.org/10.3390/biology11101485
Submission received: 3 September 2022
/
Revised: 27 September 2022
/
Accepted: 5 October 2022
/
Published: 11 October 2022
(This article belongs to the Special Issue Diopatra: The Amazing Ecosystem Engineering Polychaetous Annelid)
Abstract
:Simple Summary
Most of the world is ocean, and most of the ocean bottom is mud or sand. Understanding the ecology of sedimentary habitats is therefore important for understanding marine ecosystems writ large. Marine sediments are typically occupied by burrowing and tube-building animals that physically structure the habitat. In coastal sediments, an especially widespread example is the annelid worm Diopatra, which builds large tubes up to 2 m deep and 1 cm in diameter. These tubes have extensive physical effects on other organisms in the habitat, including commercially important fish and crustaceans. Diopatra are currently being impacted by climate change, species invasions, and (in some areas) the bait-digging industry. In this article, I review what we know about Diopatra ecology with an eye to identifying major open questions and future threats facing this important architect of coastal marine systems.
Abstract
A well-known example of marine ecosystem engineering is the annelid genus Diopatra, which builds large tubes in coastal sediments worldwide. Early studies of Diopatra were among the first to recognize the importance of facilitation in ecology, and Diopatra has become a key marine soft-sediment application of the ecosystem engineering concept. Here, I review our current knowledge of Diopatra ecology, including its natural history, ecosystem engineering effects, and trophic relationships. I particularly explore how human activities are influencing Diopatra in terms of climate change, bait fishing, and species invasions. Most of what we know about Diopatra ecology comes from focal studies of a few species in a few well-known regions. Further evaluating how our current understanding applies to other species and/or other regions will help to refine and deepen our understanding of structure and function in marine systems.
1. Introduction
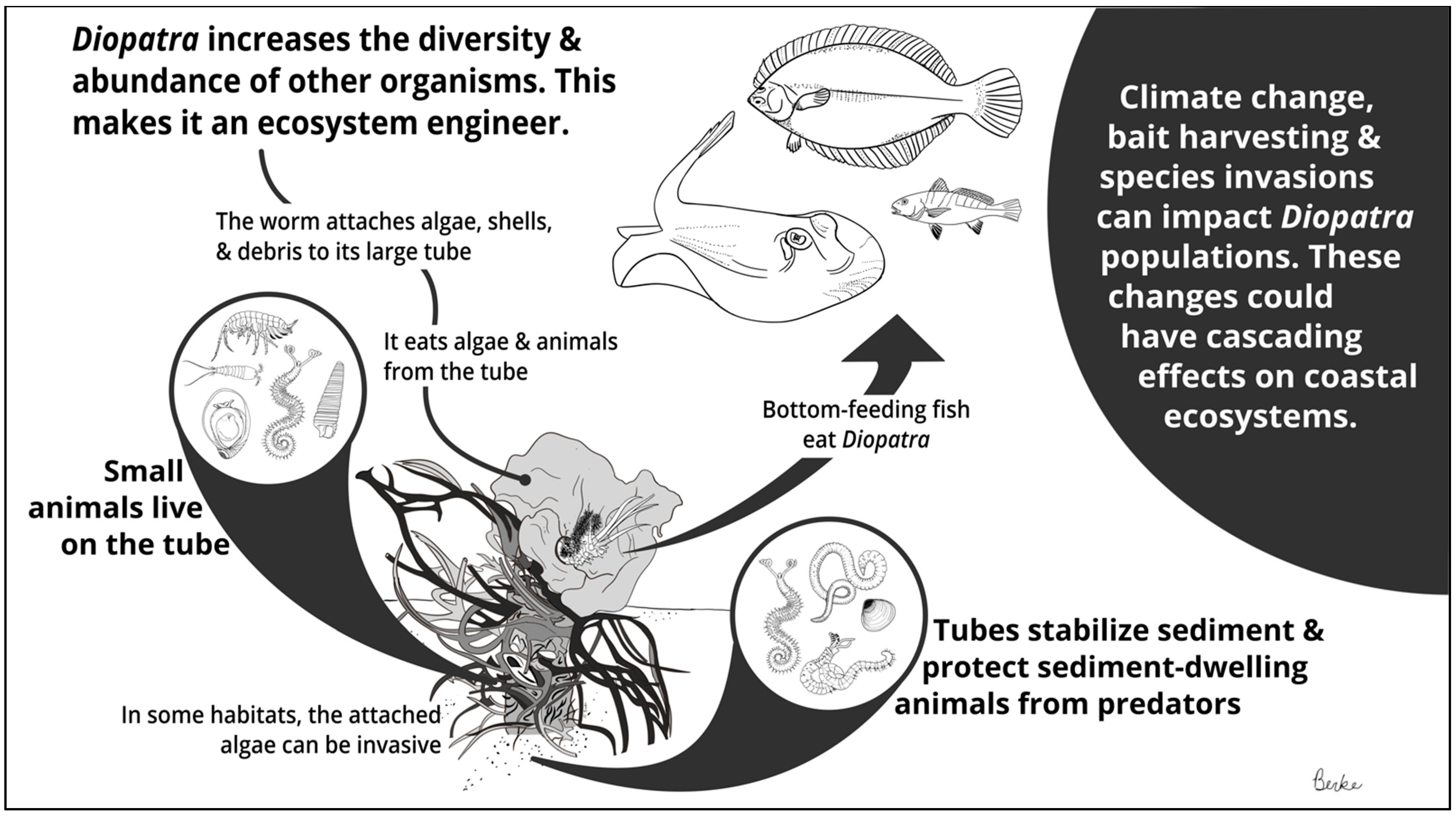
Onuphid polychaetes in the genus Diopatra have captured scientific interest for more than two centuries. The earliest descriptions of Diopatra [1,2,3] remarked on the worm’s size, beauty, and charismatic habit of festooning its tube with fragments of shell, drift algae, and other debris; by the mid-20th century, ecologists increasingly recognized that Diopatra is not only charming but also ecologically important. Some of the earliest examples of facilitation between species were reported for Diopatra [4]. Such effects, now regarded as ecosystem engineering [5,6], are largely due to the physical and biological impacts of the worm’s tube structure (Figure 1). Diopatra’s robust tubes descend deep into the sediment, emerging above the surface in a decorated ‘tube cap’ [7,8]. Diopatra tube caps provide physical structure in habitats where structure is scarce, thereby facilitating a wide variety of epibenthic organisms [9,10,11]. The tubes also stabilize sediments and create predation refugia for infauna [4,12]. Here, I review our current knowledge of Diopatra ecology, including its natural history, role as an ecosystem engineer, and responses to anthropogenic change. My goals are to synthesize existing knowledge and to identify open questions regarding this interesting genus.
Taxonomic Caveat
Polychaete taxonomy in general—and Diopatra taxonomy specifically—is in a period of especially rapid revision [13,14,15,16]. Like many polychaetes, the earliest-recognized Diopatra species were historically thought to show cosmopolitan distributions. The name D. neapolitana has been applied to worms from Europe, the Mediterranean, Africa, and Southeast Asia [8,17,18,19]. Similarly, the name D. cuprea has been applied to worms throughout North America, Central America, Brazil [11,20], the eastern Atlantic [21], and the Indian Ocean [22,23]. We now recognize that such cosmopolitan distributions are quite unlikely [14]. Old species descriptions are being rapidly split and revised as modern systematists, armed with molecular toolkits, generate a cornucopia of new species. This work is revealing enormous diversity within the Diopatra genus [13,15,16,24,25], and, undoubtedly, more species await description.
Many important contributions to Diopatra ecology were made in publications that pre-date this recent burst of novel taxonomic insight, raising questions about which species were actually the focus of any given study. Although this is a real challenge, we should not lose sight of the enormous value provided by these ecological studies. In this review, I will use the species name associated with the original publication. In cases where the original name is clearly not correct (i.e., uses of D. neapolitana for regions far from western Europe and the Mediterranean), I will indicate this by simply using the genus name and including a brief remark about the named used in the study. Anyone working on Diopatra should stay abreast of taxonomic developments in their geographic region. Ecologists would do well to keep voucher specimens and tissue samples for genetic analysis whenever possible. Increased collaboration between ecologists and taxonomists would be most welcome.
2. Natural History
Diopatra species occur in temperate and tropical coastal and estuarine systems worldwide [18,25,26,27,28,29]. Like most estuarine species, Diopatra exhibits wide salinity tolerance from 15 to 40 ppt, and at least some species can survive short periods outside that range [30]. Typical habitats include protected intertidal and shallow subtidal flats of muddy sand or sandy mud—generally not pure sand, pluff mud, or clay [1,31]. Within a geographic region, D. cuprea density is higher in areas with faster currents, but this trend breaks down when drawing comparisons across geographic regions [31]. The best-known Diopatra species occupy a tidal range from the mid intertidal to the shallow subtidal [11,32], but some species occur at greater depths, e.g., D. ornata in the eastern Pacific, D. leuckarti in Hawaii [29], and multiple new species recently described from the continental shelf of Africa [16].
Diopatra is notable for constructing, in Verrill’s words, “a very curious permanent tube in which it dwells very securely” [1]. Most of the worm’s ecosystem engineering effects are tied to this tube-building behavior. The tube extends downward 1 m or more beneath the sediment. The above-sediment portion of the tube varies across species but typically emerges several centimeters above the sediment surface and is decorated with shell fragments, detritus, and algae [13], with the aperture opening either perpendicular to the sediment or pointing down at the sediment [33,34,35]. Exceptions to this typical phenotype include D. budaevae and D. hektoeni, both of which exhibit thinner, more fragile (but still decorated) tubes [13], and D. neapolitana, which constructs tubes either flush with the sediment surface or only slightly emergent, with little to no decoration [13,36]. Whereas most Diopatra occur singly, at least two species, D. leuckarti and D. ornata, have been described as forming mounds or reefs of elevated sediment [29,37]. Tube morphology is a useful field characteristic; whenever possible, descriptions and photographs of tubes should be included in taxonomic descriptions. Tube morphology can also vary within a species; data about the range of decoration and tube size for a given species (as collected by Wethey et al. in their 2016 study [36]) are helpful, particularly if they can help disambiguate co-occurring species in the field.
New recruits build a tube upon settlement, and the tube is expanded (both in diameter and depth) as the animal grows. The tube’s innermost layers consist of a parchment-like material formed by a mucous secretion that polymerizes in seawater. The outermost layer of the tube consists of foreign matter imbricately cemented to the structure, with each object extending outward from the tube wall. To attach an item, the worm holds it in position with its jaws, palps, and anterior-most parapodia (softer materials are sometimes first trimmed with the jaws), then glues it into place by rubbing its anterior ventral surface over the attachment point, secreting mucous from glands beginning at the sixth segment [2]. Myers [34] found that D. cuprea prefers ‘tabular’ materials and appears to utilize progressively smaller items along the axis from the tube opening toward the sediment surface, otherwise selecting material indiscriminately. Preferences have not been assessed for other species.
Why does Diopatra decorate its tube? Algae and invertebrates found on D. cuprea tubes are also found in its gut, suggesting that ‘gardening’ plays a major role (reviewed in [11,38]). Decoration extending from the tube cap may function like ‘whiskers’ to help the worm detect disturbance and avoid predation [39]. Some data for D. cuprea suggest that decoration disrupts solenoidal eddies as current moves past the tube, thereby reducing sediment scour (B. Little and M. LaBarbera, unpublished data). Crypsis has also been suggested, but experimental tests have not supported this hypothesis for D. cuprea [40]. Naturally, species such as D. neapolitana with non-emergent/non-decorated tubes would not be able to feed on the tube cap or enjoy benefits such as scour reduction; this raises questions about how D. neapolitana feeding and environmental tolerances differ from those of other members of the genus and how its ecosystem effects differ from those established for D. cuprea and others with emergent tubes. D. neapolitana appears to occupy lower-energy, muddier habitats than other species (S. Woodin, personal communication), perhaps rendering sediment scour less problematic. It is also sluggish and easier to dig in comparison to D. cuprea (personal observation) and D. biscayensis (S. Woodin, personal communication). This might point to differences in D. neapolitana metabolism and diet. Understanding how tube-worm–sediment interactions vary across the range of tube phenotypes in the genus would be an interesting area for further study.
Diopatra occupies physically dynamic habitats, subject to both erosion and deposition resulting from variable currents and storms. D. cuprea responds to erosion by trimming off excess tube material with the jaws, thus maintaining a tube cap with a height of roughly 2–5 cm. In response to sediment deposition, the worm extends the tube upward through the sediment and rapidly establishes a new tube cap [34]. This response to deposition and erosion has led some researchers to suggest that Diopatra tubes can be used to measure sediment dynamics at a given site [22,33]. Tube caps can be lost during storms or due to the activities of epibenthic organisms. It is common to see Diopatra caps in the wrack zone or accumulated in the bottom of stingray feeding pits (pers obs). During Mogula settlement season, worms may cut off tube caps that have become too overgrown with tunicates (S. Woodin, personal communication). D. cuprea can entirely rebuild a lost tube cap within ~12 hrs. For populations at high densities, D. cuprea tends to orient tubes either perpendicular or parallel to neighboring tubes (presumably allowing the worm to easily feed from its neighbor). At lower population densities, however, tubes are oriented perpendicular to the direction of prevailing currents. This may facilitate passive diffusion from the tube and/or could reduce sediment deposition inside the tube, particularly in oscillatory flow conditions [41]. In very high flows, the tube opening actually collapses, which may further prevent sediment influx [42].
3. Physical Effects of the Tube
Aggregations of polychaete tubes generally stabilize sediment, as evidenced by reduced ripple formation and the development of a diatom layer on the sediment surface within polychaete beds [43,44,45,46]. This is somewhat counterintuitive, given that eddies generated as water flows past tube structures cause upstream sediment scour and downstream deposition [47,48]. However, such destabilizing effects are evidently countered by multiple mechanisms that enhance stability. These include increased sediment binding due to mucous and microbial growth associated with animal activities [45,47,49,50], as well as skimming flow over tube beds [45,51]. Whereas specific conditions at any given site depend on local flow regimes, tube sizes, and tube density, tube mimics can create skimming flow when occupying as little as 8.8% of the available area [51]. This threshold would be met or exceeded in many Diopatra beds according to published field abundances (e.g., [31,52]), particularly when considering that tube decoration extends roughness well beyond the area occupied by the tube itself. Sediment deposition from skimming flow could well account for “mounds” associated with D. leuckarti and D. ornata in the Pacific Ocean [29,37].
It is possible that the tube helps Diopatra colonize mobile sediments, and their stabilizing effect makes the area suitable for other infauna; this hypothesis is supported by observations that a Diopatra species (referred to as D. neapolitana but almost certainly something else) was among the first species to colonize an eroded sand flat in Malaysia following disruption by a typhoon. Their establishment preceded colonization by other organisms [53].
4. Facilitation of Macroalgae and Plants
Diopatra interactions with macroalgae have been best established for D. cuprea in the northwestern Atlantic. By actively attaching drift algae to its tube, D. cuprea facilitates an algal canopy in habitats that would otherwise lack stable, attached algal populations (Table 1); Diopatra thus acts as a foundation species in a facilitation cascade [54,55]. Seagrasses, may also be facilitated when D. cuprea attaches reproductive shoots [56]. A single D. cuprea tube can support more than 300 mg of algal biomass representing multiple species [11]. Whereas the quantitative data for Diopatra facilitation of algae is exclusively from D. cuprea, one would expect similar effects for any species that builds similarly decorated tubes. It is worth noting, however, that even for D. cuprea, the interaction with algae can vary spatially. In habits with fast current, tubes constructed entirely of shell debris are often observed (personal observation), perhaps because drift algae passes by too quickly for worms to catch it and/or because algae would increase drag forces, leading to cap breakage in high flows. There also exists a well-documented latitudinal gradient in which D. cuprea decoration decreases dramatically in the southern portion of its range. This is driven by behavioral variability rather than by algal availability [11]. Whereas geographic patterns in behavior could point to a cryptic species complex, CO1 genetic patterns do not entirely mirror the behavioral patterns (Sotka et al., this issue). The mechanism underlying the behavioral variability remains an open question.
D. cuprea’s decorating behavior has notably been implicated in facilitating the invasive Agarophyton vermiculophyllum (formerly Gracilaria vermiculophylla) in the western Atlantic [52,57,58]. The extent to which D. cuprea has accelerated the invasion is difficult to quantify; A. vermiculophyllum is an aggressive invader in its own right and would undoubtedly have invaded even without D. cuprea (as is happening in many habitats worldwide [59]). However, D. cuprea indisputably anchors and stabilizes extensive A. vermiculophyllum mats. A. vermiculophyllum itself provides habitat for epibenthic invertebrates [58] and nursery habitat for juvenile blue crabs [60]. The D. cuprea–A. vermiculophyllum relationship may therefore be amplifying a habitat cascade, indirectly enhancing secondary productivity in some habitats [61]. However, two important caveats must be made: first, we do not know the extent to which D. cuprea + A. vermiculophyllum functions differently than D. cuprea + native macroalgae. Second, superblooms of A. vermiculophyllum have been associated with sediment anoxia and D. cuprea death [52], suggesting that facilitation of invertebrate communities only occurs below a threshold of A. vermiculophyllum abundance. As the invasion proceeds, it is unclear whether systems will stabilize in an enhanced-functioning state or an anoxic, reduced-functioning state.

{kind=link}
Table 1.
Studies quantifying Diopatra facilitation of algae and plants.
| Study | Species | Location | Effects |
|---|---|---|---|
| Mangum et al., 1968 [31] | D. cuprea | Chesapeake Bay, Virginia, USA | 20 algal species identified from tubes |
| Harwell and Orth 2001 [56] | D. cuprea | Chesapeake Bay, Virginia, USA | Tubes facilitate reproductive seagrass shoots |
| Thomsen 2004 [62] | D. cuprea | Hog Island Bay, Virginia, USA | Tubes facilitate Ulva and A. vermiculophyllum |
| Thomsen & McGlathery 2005; Thomsen et al., 2005 [57,63] | D. cuprea | Hog Island Bay, Virginia, USA | Tubes facilitate invasive alga Agarophyton vermiculophyllum |
| Berke 2012 [11] | D. cuprea | Northwest Atlantic (Massachusetts through Florida, USA) | Tubes support a total of 34 species (as many as 15 species within a single region) |
5. Facilitation of Infauna
Diopatra tubes provide refugia for infauna by physically excluding predators, such as crabs, Limulus, epibenthic fish, skates, rays, and shorebirds (Table 2). This effect appears to be driven by the physical structure of the tube—tube mimics built of soda straws have the same effect [12]. In addition to predator exclusion, tube effects on local flow dynamics may promote entrainment of passively dispersing larvae, increasing recruitment near tubes [64]. Whereas most of the work on Diopatra facilitation of infauna has focused on D. cuprea, similar effects have been described for D. ornata, D. leuckarti, and others (Table 2).
Whereas Diopatra facilitation of infauna appears to be widespread, multiple factors can influence the strength of the interaction. For example, seasonal factors may contribute to variability; Santos and Aviz (2018) found that infauna on a beach with scattered Diopatra (densities of 25–75 m−2) were more diverse and abundant during the rainy season in comparison to a nearby site with no Diopatra. However, this effect was not observed during the dry season. Tube density can also be important; for example, Woodin [4,12] found that a density of 6 · 0.01 m−2 had a clear effect at an inlet site in Virignia, USA, whereas single tubes had no effect. Similarly, Bell and Woodin [65] tested zero, one, three, and six tubes in 0.01 m−2, finding significant differences for only the most extreme zero vs. six tube comparison. This is also consistent with Ban and Nelson’s [66] finding that four tubes in 0.01 m−2 had no effect at a subtropical site in the Indian River Lagoon, FL, USA. In contrast, Thomsen et al. (2011) found an effect of single onuphid tubes (presumed to be a Diopatra species) on a sand flat in Mozambique [23], whereas Santos and Avis (2018) found elevated density/abundance in an area with scattered single Diopatra tubes in Brazil [20] (referred to as D. cuprea, although Diopatra taxonomy in this region has been recently revised). Both of these studies make it somewhat difficult to disambiguate effects of the tube-cap fauna from true infauna; Thomsen et al. [23] compared cores with sediment + tube cap to tube caps alone, making it possible to infer that infauna were affected separately from the tube-cap fauna. In the Santos and Avis study [20], infauna and the tube-cap fauna were analyzed together, so disambiguation is not possible. Some variability in Diopatra effects on infauna might reflect differences in local hydrodynamic regimes and subsequent effects on larval recruitment. Variability could also stem from geographic and seasonal differences in processes such as recruitment and predator abundance. If tubes are physically excluding predators, then one would expect predator size distributions and specific foraging behaviors to play a role. Interactions among smaller infauna may also be important; for example, when facilitated infauna prey on smaller meiofauna and juvenile bivalves, complex patterns can emerge [65,67].
Interestingly, even as Diopatra facilitates other infauna, the worm itself might be sensitive to competition, particularly at the settlement stage. Flamingo exclusion structures in Namibia reduced Diopatra abundance, even as all other infauna increased [68] (referred to as D. neapolitana, although Namibia is far outside its range). This contrasts with Woodin’s (1981) finding of enhanced D. cuprea recruitment in predator-exclusion cages in Virginia, USA. This contrast emphasizes that we have much to learn about the processes influencing Diopatra recruitment in different systems.
Table 2.
Studies quantifying Diopatra facilitation of infauna.
| Study | Species | Location | Densities Tested | Effects |
|---|---|---|---|---|
| Woodin 1978 [4] | D. cuprea | Tom’s Cove, Virginia, USA | 0, 1, 6 · 0.01 m−2 | ↑ infaunal richness and abundance at 6 · 0.01 m−2 |
| Woodin 1981 [12] | D. cuprea | Tom’s Cove, Virginia, USA | 0, 1, 6 · 0.01 m−2 | ↑ infaunal abundance at 6 · 0.01 m−2 |
| Bell and Woodin 1984 [65] | D. cuprea | Tom’s Cove, Virginia, USA | 0, 1, 3, 6 · 0.01 m−2 | ↑ polychaete abundance at 6 · 0.01 m−2; no effect on meiofauna |
| Bailey-Brock 1984 [29] | D. leuckarti | Niu Valley, Hawaii, USA | “mounds” up to 21,800 m−2 | 28 species from 7 phyla are associated with mounds |
| Luckenbach 1984 [69] | D. cuprea | North Inlet, South Carolina, USA | 0, ≥9 · 0.01 m−2 | ↑ infaunal abundance near tubes |
| Luckenbach 1984 [70] | D. cuprea | North Inlet, South Carolina, USA | 0, ≥10 · 0.01 m−2 | ↑ infaunal abundance in areas of high tube density |
| Ban and Nelson 1987 [66] | D. cuprea | Indian River Lagoon, Florida, USA | 0, 1, 4 · 0.01 m−2 | No effect |
| Ambrose & Anderson 1990 [71] | D. ornata | Pendleton Artificial Reef, California, USA | Inside vs. outside “beds” | ↑ richness and abundance of infauna and decapods |
| Thomsen et al., 2011 [23] | Diopatra-like onuphid * | Inhaca Island, Mozambique | 0, 1 · 0.01 m−2 | ↑ richness and abundance around single tubes |
| Santos and Aviz 2018 [20] † | D. cuprea * | Algodoal-Maiandeua, Brazil | 0 vs. 25–75 m−2, but each sample from the Diopatra area included only 1 | Seasonal ↑ richness and abundance |
* Species in this region have been recently revised. † Sampling captured both infauna and the tube-cap, making it impossible to separate infauna from epifauna.
6. Facilitation of Epibenthic Fauna
Diopatra tubes directly provide habitat for smaller epibenthic fauna, including a diverse array of protists, meiofauna, gastropods, bivalves, amphipods, cnidarians, flatworms, bryozoans, and tunicates (Table 3). Tube-cap fauna are most strongly facilitated when tubes are decorated with macroalgae, although caps lacking algae do also support an epifaunal community [58,72,73]. Larger organisms, such as juvenile fish and crabs, also utilize Diopatra tubes. In the northwestern Atlantic, juvenile fish show a diel pattern of habitat use, spending daylight hours sheltering in D. cuprea beds but venturing into open habitats to forage at night [10]. Mounds of D. leuckarti and D. ornata support elevated diversity and abundance of epibenthic crabs and shrimp [29,71]. In habitats where Diopatra facilitates A. vermiculophyllum (e.g., most of the US Atlantic coast), we would also expect it to indirectly facilitate juvenile blue crab, which have higher survivorship in A. vermiculophyllum [55,60].
7. Food Web Connections
Diopatra are omnivores, primarily feeding on organisms living on the tube cap, on neighboring tube caps, and within nearby sediments. D. cuprea is capable of extending 10 cm or more from the tube but appears to spend most of its time grazing within a radius <5 cm (pers. obs.). This species exhibits a strong feeding response to extracts of other polychaetes, bivalves, and Artemia but also shows a weaker feeding response to other arthropods, echinoderms, Fundulus, Codium, and Zostera [75]. Gut contents include a diverse array of invertebrates, protists, Zostera, chlorophytes, rhodophytes, and phaeophytes [31], and I have seen worms eat Ulva in the laboratory.
Diopatra will scavenge to at least some extent; in North Inlet SC, I once encountered a number of dead minnows littering a D. cuprea flat. At least one worm had attached a fish to its tube cap, and another doggedly attempted to pull a fish into its tube, seemingly undaunted by the physical impossibility of this task. That said, D. cuprea held in the lab will ignore shrimp meat that is more than a day old, so true scavenging seems unlikely.
Diopatra are found in the guts of multiple epibenthic fish, including flounder, skates, pigfish, croaker, and turbot [76,77,78]. One report also describes the large gastropod Fasciolaria hunteria feeding on D. cuprea (and the similar Americonuphis magna) by inserting its long proboscis down the tube and rasping at the worm’s tissue [79]. Diopatra avoids predation by withdrawing rapidly to depth; the strength of this response varies within the genus, as indicated by the difficulty of collecting worms. For example, D. cuprea requires skill to dig; the worm is quite vibration-sensitive, so one must step gingerly and strike rapidly with the shovel. In contrast, I was surprised by how easy it was to dig D. neapolitana in Europe. I suspect this difference is at least partly why D. neapolitana is widely harvested for fishing bait [28,80,81], whereas D. cuprea is not.
Because Diopatra can regenerate both anteriorly and posteriorly, predation attempts are not always lethal ([81] and references therein). The worm can cling very tightly to the tube, using hydrostatic pressure to press the chaetae into the textured tube wall [82]. This, together with the tube’s depth, make it generally impossible to pull a worm entirely out of its tube, as the worm will simply break in the attempt. Unsurprisingly, then, worms undergoing anterior regeneration are commonly collected in the field. These represent anywhere from 5–40% of specimens, depending on the species, place, and season (summarized in [81]). Antennae are also frequently nipped off by predators, and antennal regeneration is even more common than head regeneration [83].
8. Human Impacts
Diopatra species are impacted by human activities on multiple fronts; climate change, harvesting for the bait trade, human-assisted transport, and interactions with invasive species are all important aspects of Diopatra biogeography and ecology.
8.1. Range Expansions and Climate Change
Diopatra occupies tropical and temperate waters, with poleward range limits evidently set by cold temperature limits on summer reproduction; D. neapolitana and D. biscayensis do not occur at sites where August sea-surface temperature (SST) remains below 18 °C, suggesting that warmer temperatures are needed for successful reproduction [36,84]. Cold winter temperatures may also play a role by inhibiting feeding and tube maintenance [34,75,85]. At cold temperatures, D. cuprea stops feeding between 5–8 °C [75,85] and stops maintaining its tube below 1.8 °C [34]. We would therefore expect Diopatra ranges to extend poleward with climate change, as has been demonstrated for D. biscayensis in western Europe, where the worm’s progressive northward expansion has matched warm SST anomalies in the Bay of Biscay [84]. The D. biscayensis range shift has been accelerated by human transport, most likely associated with mussel aquaculture [36,86,87].
In the northwestern Atlantic, we have not seen similar poleward movement in D. cuprea [35]—at least, not as of 2022 (Berke unpublished data). This is somewhat surprising, as the Gulf of Maine has been warming rapidly [88]. Whether this reflects different thermal biology, constraints of larval supply, lack of intertidal aquacultural transport, or something else remains an open question. In Europe, the lecithotrophic larvae of D. neapolitana and D. biscayensis both have a short pelagic period that limits their dispersal ability; range shifts in D. biscayensis are therefore human-assisted [86,87]. D. cuprea appears to have a similarly short larval period [89], which may account for its as-yet stable northern range limit.
The extent to which Diopatra will be impacted by higher temperatures in tropical and subtropical habitats remains largely unexplored. Diopatra thermal tolerances vary seasonally and geographically; a 1969 study found that temperatures of 37–38 °C were 100% lethal for D. cuprea from Barnstable, Massachusetts, and 50% lethal for worms from Beaufort, NC, during winter months. However, Beaufort worms acclimatized in summer months to a 50% lethal limit near 42 °C. Linking such laboratory data to field conditions will be complicated by the need to understand what temperatures worms actually experience in situ. Benthic temperatures can depart substantially from SSTs, and intertidal organisms can experience temperatures considerably different from the surrounding habitat [90,91,92,93,94]. Whereas models have been developed to estimate sediment surface temperature for intertidal mud flats [95], infauna will experience progressively lower temperatures as their burrowing depth increases [96]. Diopatra’s tube extends much deeper into the sediment than most infauna, which should buffer it against surface extremes. Furthermore, worms must irrigate the tubes with overlying water and must partly emerge from the tube to forage, so exposure to ambient water temperatures could be substantial. Understanding how worms such as Diopatra experience temperature over the course of a day, season, or year and how this would differ for intertidal vs. subtidal populations are interesting questions. For the lugworm Arenicola marina, temperatures at burrow depths appear to correlate well with SST, allowing biogeographic models to predict likely responses to warming oceans [94]. Whereas lugworm burrows are shallower and more permeable than Diopatra tubes, this modeling approach holds considerable promise for understanding climate responses in Diopatra and infaunal organisms in general.
8.2. Bait Harvesting
Throughout western Europe, intertidal Diopatra are dug for use as fishing bait. In Portugal’s Canal de Mira (Ria de Aveiro estuary), as many as 4.3 million individual D. neapolitana may be harvested each year [28,97,98], with an economic value of more than EUR 325,000/yr [28]. Whereas D. neapolitana can regenerate anterior segments, this ability is limited to the first 15–20 chaetigers, and bait diggers typically collect more than that, suggesting that the activity is generally lethal [81]. Modeling based on catch per unit effort has suggested that D. neapolitana harvesting remains short of maximum sustainable yield for the system [98]. However, Cunha et al. [28] estimated that diggers collected roughly 2.9 worms · m−2 over a period of a few winter days, a number that is alarmingly close to their estimate for the standing population density of 2.8 worms · m−2. Both estimates were based on a fairly limited one-time field survey, so we should not derive too many conclusions (as the authors themselves emphasize). Nonetheless, the disconnect between this field survey and CPUE models highlights the need for a much more complete understanding of D. neapolitana population dynamics and the long-term sustainability of harvesting pressure on this species.
8.3. Interactions with Invasive Species
Given that Diopatra facilitate both algae and invertebrates, the potential for facilitating invasions is high. In the western Atlantic, D. cuprea is facilitating the invasive red alga Agarophyton vermiculophyllum [52,57,59,99]. A. vermiculophyllum is an aggressive invader in its own right and would likely be spreading even without D. cuprea facilitation; it indisputably forms thick mats anchored to D. cuprea tubes. These mats can provide habitat for smaller invertebrates [58,60] which, in turn, increases D. cuprea foraging opportunities [100], so effects of this invader are not universally negative. However, A. vermiculophyllum mats can also create anoxia in the sediment, which has been associated with mass D. cuprea die-offs, possibly contributing to long-term population declines throughout the mid-Atlantic US [52]. Conditions leading to A. vermiculophyllum blooms, their frequency, and effects on invertebrates, including D. cuprea, remain open questions.
Whereas Diopatra species are not generally invasive, the population of D. biscayensis occurring north of the Brittany peninsula in France is evidently the result of human transport from the Bay of Biscay [86,87]. This raises the possibility that other Diopatra species could possibly have been moved through aquaculture and the bait trade. Most Diopatra appear to have a short-lived larval period, making transport in ballast water unlikely, but juveniles could be transported as hitch-hikers in mud and algae associated with aquaculture [87]. As D. biscayensis expands north, it will introduce large tube structures into areas previously dominated by the bioturbator A. marina. This change would be expected to result in an overall increase in sediment stability, productivity, and local diversity [84]. As new Diopatra species are described, researchers should bear in mind the possibility of human-assisted transport.
9. Future Directions
Diopatra occur worldwide and are ecologically important members of coastal sedimentary communities. However, in many ways, we know more about the worm’s effects on other organisms than we know about the worm itself; surprisingly little has been published about Diopatra autecology and basic biology. We generally need better understanding of the worm’s environmental tolerances, feeding biology, reproductive cycles, and population dynamics for all Diopatra species, especially in populations subject to harvesting or existing near geographic range limits. In particular, understanding thermal biology and responses to climate change will require learning more about the relationship between SST, bottom temperatures, and an individual Diopatra’s experienced temperature.
These gaps in our knowledge become especially evident as new species are identified. Taxonomists have made enormous progress in recent years, disentangling species complexes and revealing hitherto unknown diversity. Comparing and contrasting the ecology and ecosystem engineering effects of different Diopatra in different localities should be a priority. This work will not be possible without close collaboration between ecologists and systematists. Funding to support taxonomic work alongside ecological investigations is critically important. Understanding how ecosystem engineering effects vary with Diopatra species, habitat, and season may help us better understand how predation, competition, recruitment, and facilitation structure sedimentary communities writ large.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
I am forever grateful to Sally Woodin for teaching me how to dig Diopatra, how to read a mud flat, and how to be a scientist. Thank you, Sally, for everything. Thanks also to the institutions that have supported my work on Diopatra over the years, including the University of South Carolina, the Smithsonian Environmental Research Center, the Smithsonian Marine Station at Fort Pierce, the Virginia Institute of Marine Science Eastern Shore Laboratory, and Siena College.
Conflicts of Interest
The author declares no conflict of interest.
References
- Verrill, A.E.; Smith, S.I. Report upon the Invertebrate Animals on Vineyard Sound and Adjacent Waters, with an Account of the Physical Features of the Region; U.S. Goverment Printing Office: Washington, DC, USA, 1874.
- Linville, H.R. The Natural History of Some Tube-Forming Annelids. Mark Anniversary Volume; to Edward Laurens Mark, Hersey Professor of Anatomy and Director of the Zoological Laboratory at Harvard University; H. Holt and Company: New York, NY, USA, 1903; pp. 227–237. [Google Scholar]
- Saint-Joseph, B. de Les Annélides Polychètes des Côtes de France (Manche et Océan). Ann. Des Sci. Nat. (Zo-Ologie Paléontol.) 1898, 5, 209–222. [Google Scholar]
- Woodin, S.A. Refuges, Disturbance, and Community Structure: A Marine Soft-Bottom Example. Ecology 1978, 59, 274–284. [Google Scholar] [CrossRef]
- Wright, J.P.; Jones, C. The Concept of Organisms as Ecosystem Engineers Ten Years on: Progress, Limitations, and Challenges. BioScience 2006, 56, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Berke, S.K. Functional Groups of Ecosystem Engineers: A Proposed Classification with Comments on Current Issues. Integr. Comp. Biol. 2010, 50, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, O. Catalogue of the Polychaeta; Allan Hancock Foundation, University of Southern California Press: Los Angeles, CA, USA, 1959. [Google Scholar]
- Day, J.H. Polychaeta of Southern Africa Part 2: Sedentaria; Trustees of the British Museum (Natural History): London, UK, 1967. [Google Scholar]
- Bell, S.S.; Coen, L.D. Investigations on epibenthic meiofauna I. Abundances on and repopulation of the tube-caps of Diopatra cuprea (Polychaeta: Onuphidae) in a subtropical system. Mar. Biol. 1982, 67, 303–309. [Google Scholar] [CrossRef]
- Diaz, R.J.; Cutter, G.R.; Able, K.W. The importance of physical and biogenic structure to juvenile fishes on the shallow inner continental shelf. Estuaries 2003, 26, 12–20. [Google Scholar] [CrossRef]
- Berke, S.K. Biogeographic variability in ecosystem engineering: Patterns in the abundance and behavior of the tube-building polychaete Diopatra cuprea. Mar. Ecol. Prog. Ser. 2012, 447, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Woodin, S. Disturbance and Community Structure in a Shallow Water Sand Flat. Ecology 1981, 62, 1052–1066. [Google Scholar] [CrossRef]
- Paxton, H.; Arias, A. Unveiling a surprising diversity of the genus Diopatra Audouin & Milne Edwards, 1833 (Annelida: Onuphidae) in the Macaronesian region (eastern North Atlantic) with the description of four new species. Zootaxa 2017, 4300, 505–535. [Google Scholar]
- Hutchings, P.; Kupriyanova, E. Cosmopolitan polychaetes—Fact or fiction? Personal and historical perspectives. Invertebr. Syst. 2018, 32, 1–9. [Google Scholar]
- Elgetany, A.H.; Van Rensburg, H.; Hektoen, M.; Matthee, C.; Budaeva, N.; Simon, C.A.; Struck, T.H. Species delineation in the speciation grey zone—The case of Diopatra (Annelida, Onuphidae). Zool. Scr. 2020, 49, 516–534. [Google Scholar] [CrossRef]
- Hektoen, M.M.; Willassen, E.; Budaeva, N. Phylogeny and Cryptic Diversity of Diopatra (Onuphidae, Annelida) in the East Atlantic. Biology 2022, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Choe, S. On the Life History of the Polychaete Worm, Diopatra neapolitana Delle Chiaje. Nippon Suisan Gakkaishi 1960, 26, 430–437. [Google Scholar] [CrossRef]
- Parameswaran, V.S. Distribution of Diopatra neapolitana Delle Chiaje (Polychaeta) in the South West Coast of India. Indian J. Mar. Sci. 1973, 2, 62–63. [Google Scholar]
- Fauchald, K.; Berke, S.K.; Woodin, S.A. Diopatra (Onuphidae: Polychaeta) from Intertidal Sediments in Southwestern Europe. Zootaxa 2012, 3395, 47–58. [Google Scholar] [CrossRef]
- Santos, T.; Aviz, D. Macrobenthic fauna associated with Diopatra cuprea (Onuphidae: Polychaeta) tubes on a macrotidal sandy beach of the Brazilian Amazon Coast. J. Mar. Biol. Assoc. UK 2018, 99, 751–759. [Google Scholar] [CrossRef]
- Monro, C. XVII.—On the post-larval stage in Diopatra cuprea, Bosc, a Polychætous Annelid of the family Eunicidæ. Ann. Mag. Nat. Hist. 1924, 14, 193–199. [Google Scholar] [CrossRef]
- De, C. Application of a Biological Tool for Estimating Current Annual Rates of Erosion and Deposition in Modern Coastal Environments: A Case Study in the Bay of Bengal Coast. Mar. Georesour. Geotechnol. 2002, 20, 209–220. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Muth, M.F.; McGlathery, K.J. Tube-forming polychaetes enhance invertebrate diversity and abundance in sandy sediments of Mozambique, Africa. Afr. J. Mar. Sci. 2011, 33, 327–332. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. The cryptogenic bait worm Diopatra biscayensis Fauchald et al., 2012 (Annelida: Onuphidae)—Revisiting its history, biology and ecology. Estuar. Coast. Shelf Sci. 2015, 163, 22–36. [Google Scholar] [CrossRef]
- Seixas, V.C.; Steiner, T.M.; Solé-Cava, A.M.; Amaral, A.C.Z.; Paiva, P.C. Hidden diversity within the Diopatra cuprea complex (Annelida: Onuphidae): Morphological and genetics analyses reveal four new species in the south-west Atlantic. Zool. J. Linn. Soc. 2020, 191, 637–671. [Google Scholar] [CrossRef]
- Hartman, O. Atlas of the Sedentariate Polychaetous Annelids from California; Allan Hancock Foundation, University of Southern California: Los Angeles, CA, USA, 1969. [Google Scholar]
- Paxton, H.; Fadlaoui, S.; Lechapt, J.P. Diopatra marocensis, a New Brooding Species of Onuphidae (Annelida, Polychaeta). J. Mar. Biol. Assoc. UK 1995, 75, 949–955. [Google Scholar] [CrossRef]
- Cunha, T.; Hall, A.; Queiroga, H. Estimation of the Diopatra neapolitana annual harvest resulting from digging activity in Canal de Mira, Ria de Aveiro. Fish. Res. 2005, 76, 56–66. [Google Scholar] [CrossRef]
- Bailey-Brock, J.H. Ecology of the tube-building polychaete Diopatra leuckarti Kinberg, 1865 (Onuphidae) in Hawaii: Community structure, and sediment stabilizing properties. Zool. J. Linn. Soc. 1984, 80, 191–199. [Google Scholar] [CrossRef]
- Hakkim, V.M.A. Salinity Tolerance of Diopatra neapolitana Delle Chiaje: Annelida-Polychaeta. Indian J. Mar. Sci. 1975, 4, 99–101. [Google Scholar]
- Mangum, C.P.; Santos, S.L.; Rhodes, W.R. Distribution and feeding in the onuphid polychaete, Diopatra cuprea (Bosc). Mar. Biol. 1968, 2, 33–40. [Google Scholar] [CrossRef]
- Peckol, P.; Baxter, D. Population Dynamics of the Onuphid Polychaete Diopatra cuprea (Bosc) along a Tidal Exposure Gradient. Estuar. Coast. Shelf Sci. 1986, 22, 371–377. [Google Scholar] [CrossRef]
- Myers, A.C. Some Palaeoichnological Observations on the Tube of Diopatra cuprea (Bosc): Polychaeta, Onuphidae. In Trace Fossils; Seel House Press: Liverpool, UK, 1970; pp. 331–334. [Google Scholar]
- Myers, A.C. Tube-worm-sediment relationships of Diopatra cuprea (Polychaeta: Onuphidae). Mar. Biol. 1972, 17, 350–356. [Google Scholar] [CrossRef]
- Pettibone, M.H. Polychaete Worms of the New England Region. I. Aphroditidae through Trochochaetidae. Bull. USA Natl. Mus. 1963, 227, 1–361. [Google Scholar] [CrossRef]
- Wethey, D.S.; Woodin, S.A.; Berke, S.K.; Dubois, S.F. Climate hindcasts: Exploring the disjunct distribution of Diopatra biscayensis. Invertebr. Biol. 2016, 135, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.L. The role of drift kelp in the population ecology of a Diopatra ornata Moore (Polychaeta: Onuphidae) ecotone. J. Exp. Mar. Biol. Ecol. 1992, 156, 253–272. [Google Scholar] [CrossRef]
- Berke, S.K.; Miller, M.; Woodin, S.A. Modelling the Energy—Mortality Trade-Offs of Invertebrate Decorating Behaviour. Evol. Ecol. Res. 2006, 8, 1409–1425. [Google Scholar]
- Brenchley, G.A. Predator detection and avoidance: Ornamentation of tube-caps of Diopatra spp. (Polychaeta: Onuphidae). Mar. Biol. 1976, 38, 179–188. [Google Scholar] [CrossRef]
- Berke, S.K.; Woodin, S.A. Tube Decoration May Not Be Cryptic for Diopatra cuprea (Polychaeta: Onuphidae). Biol. Bull. 2008, 214, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenchley, G.A.; Tidball, J.G. Tube-Cap Orientations of Diopatra cuprea (Bosc)(Polychaeta): The Compromise between Physiology and Foraging. Mar. Behav. Physiol. 1980, 7, 1–13. [Google Scholar] [CrossRef]
- Miller, U.C.; Bock, M.J.; Turner, E.J. Deposit and suspension feeding in oscillatory flows and sediment fluxes. J. Mar. Res. 1992, 50, 489–520. [Google Scholar] [CrossRef]
- Fager, E.W. Marine Sediments: Effects of a Tube-Building Polychaete. Science 1964, 143, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Featherstone, R.P.; Risk, M.J. Effect of tube-building polychaetes on intertidal sediments on the Minas Basin, Bay of Fundy. J. Sediment. Res. 1977, 47, 446–450. [Google Scholar] [CrossRef]
- Rhoads, D.C.; Yingst, J.Y.; Ullman, W.J. Seafloor Stability in Central Long Island Sound: Part I. Temporal Changes in Erodibility of Fine-Grained Sediment. In Estuarine Interactions; Academic Press: New York, NY, USA, 1978; pp. 221–244. [Google Scholar]
- Bolam, S.G.; Fernandes, T.F. Dense aggregations of tube-building polychaetes: Response to small-scale disturbances. J. Exp. Mar. Biol. Ecol. 2002, 269, 197–222. [Google Scholar] [CrossRef]
- Eckman, J.E.; Nowell, A.R.M.; Jumars, P.A. Sediment Destabilization by Animal Tubes. J. Mar. Res. 1981, 39, 361–374. [Google Scholar]
- Eckman, J.E.; Nowell, A.R.M. Boundary skin friction and sediment transport about an animal-tube mimic. Sedimentology 1984, 31, 851–862. [Google Scholar] [CrossRef]
- Eckman, J.E. Flow disruption by an animal-tube mimic affects sediment bacterial colonization. J. Mar. Res. 1985, 43, 419–435. [Google Scholar] [CrossRef]
- Eckman, J.E.; Thistle, D. Effects of flow about a biologically produced structure on harpacticoid copepods in San Diego Trough. Deep Sea Res. Part. A Oceanogr. Res. Pap. 1991, 38, 1397–1416. [Google Scholar] [CrossRef]
- Friedrichs, M.; Graf, G.; Springer, B. Skimming flow induced over a simulated polychaete tube lawn at low population densities. Mar. Ecol. Prog. Ser. 2000, 192, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Keller, E.L.; Berke, S.K.; Needham, C.N.; Salerno, C.R. A Double-Edged Sword: Infaunal Responses to Agarophyton vermicu-lophyllum in the Mid-Atlantic United States. Estuaries Coasts J. ERF 2019, 42, 1924–1937. [Google Scholar] [CrossRef]
- Ong, B.; Krishnan, S. Changes in the macrobenthos community of a sand flat after erosion. Estuar. Coast. Shelf Sci. 1995, 40, 21–33. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Altieri, A.; Tuya, F.; Gulbransen, D.; McGlathery, K.J.; Holmer, M.; Silliman, B.R. Habitat Cascades: The Conceptual Context and Global Relevance of Facilitation Cascades via Habitat Formation and Modification. Integr. Comp. Biol. 2010, 50, 158–175. [Google Scholar] [CrossRef]
- Gribben, P.E.; Angelini, C.; Altieri, A.H.; Bishop, M.J.; Thomsen, M.S.; Bulleri, F. Facilitation Cascades in Marine Ecosystems: A Synthesis and Future Directions. Oceanogr. Mar. Biol. 2019, 57, 127–168. [Google Scholar]
- Harwell, M.C.; Orth, R.J. Influence of a tube-dwelling polychaete on the dispersal of fragmented reproductive shoots of eelgrass. Aquat. Bot. 2001, 70, 1–7. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Gurgel, C.F.D.; Fredericq, S.; McGlathery, K.J. Gracilaria vermiculophylla (Rhodophyta, Gracilariales) in Hog Island Bay, Virginia: A Cryptic Alien and Invasive Macroalga and Taxonomic Correction. J. Phycol. 2005, 42, 139–141. [Google Scholar] [CrossRef]
- Wright, J.T.; Byers, J.E.; Devore, J.L.; Sotka, E.E. Engineering or food? mechanisms of facilitation by a habitat-forming invasive seaweed. Ecology 2014, 95, 2699–2706. [Google Scholar]
- Hu, Z.-M.; Juan, L.-B. Adaptation mechanisms and ecological consequences of seaweed invasions: A review case of agarophyte Gracilaria vermiculophylla. Biol. Invasions 2013, 16, 967–976. [Google Scholar] [CrossRef]
- Johnston, C.; Lipcius, R. Exotic macroalga Gracilaria vermiculophylla provides superior nursery habitat for native blue crab in Chesapeake Bay. Mar. Ecol. Prog. Ser. 2012, 467, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, M.S.; Ramus, A.P.; Long, Z.T.; Silliman, B.R. A seaweed increases ecosystem multifunctionality when invading bare mudflats. Biol. Invasions 2018, 21, 27–36. [Google Scholar] [CrossRef]
- Thomsen, M.S. Macroalgal Distribution Patterns and Ecological Performances in a Tidal Coastal Lagoon, with Emphasis on the Non-indigenous Codium fragile. Estuaries Coasts 2006, 29, 465–473. [Google Scholar] [CrossRef]
- Thomsen, M.; McGlathery, K. Facilitation of macroalgae by the sedimentary tube forming polychaete Diopatra cuprea. Estuar. Coast. Shelf Sci. 2005, 62, 63–73. [Google Scholar] [CrossRef]
- Eckman, J.E. Hydrodynamic processes affecting benthic recruitment1. Limnol. Oceanogr. 1983, 28, 241–257. [Google Scholar] [CrossRef]
- Bell, S.S.; Woodin, S. Community unity: Experimental evidence for meiofauna and macrofauna. J. Mar. Res. 1984, 42, 605–632. [Google Scholar] [CrossRef]
- Ban, S.M.; Nelson, W.G. Role of Diopatra cuprea Bosc (Polychaeta: Onuphidae) Tubes in Structuring a Subtropical Infaunal Community. Bull. Mar. Sci. 1987, 40, 11–21. [Google Scholar]
- Luckenbach, M.W. Effects of adult infauna on new recruits: Implications for the role of biogenic refuges. J. Exp. Mar. Biol. Ecol. 1987, 105, 197–206. [Google Scholar] [CrossRef]
- Glassom, D.; Branch, G. Impact of predation by greater flamingos Phoenicopterus ruber on the macrofauna of two southern African lagoons. Mar. Ecol. Prog. Ser. 1997, 149, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Luckenbach, M. Settlement and early post-settlement survival in the recruitment of Mulinia lateralis (Bivalvia). Mar. Ecol. Prog. Ser. 1984, 17, 245–250. [Google Scholar] [CrossRef]
- Luckenbach, M.W. Biogenic structure and foraging by five species of shorebirds (Charadrii). Estuar. Coast. Shelf Sci. 1984, 19, 691–696. [Google Scholar] [CrossRef]
- Ambrose, R.F.; Anderson, T.W. Influence of an artificial reef on the surrounding infaunal community. Mar. Biol. 1990, 107, 41–52. [Google Scholar] [CrossRef]
- Bell, S.S.; Coen, L.D. Investigations on epibenthic meiofauna II: Influence of microhabitat and macroalgae on abundance of small invertebrates on Diopatra cuprea (Bosc) (Polychaeta: Onuphidae) tube-caps in Virginia. J. Exp. Mar. Biol. Ecol. 1982, 61, 175–188. [Google Scholar] [CrossRef]
- Byers, J.E.; Gribben, P.; Yeager, C.; Sotka, E.E. Impacts of an abundant introduced ecosystem engineer within mudflats of the southeastern US coast. Biol. Invasions 2012, 14, 2587–2600. [Google Scholar] [CrossRef]
- Dudley, E.C.; Baker, A.J.; Cariello, A.; Leong, K.M.; Ballantine, A. The Occurrence of Living Mollusks on Diopatra Tube-Caps. Veliger 1989, 32, 229–230. [Google Scholar]
- Mangum, C.P.; Cox, C.D. Analysis of the Feeding Response in the Onuphid Polychaete Diopatra cuprea (Bosc). Biol. Bull. 1971, 140, 215–229. [Google Scholar] [CrossRef]
- Watanabe, K.; Tamai, K.; Numachi, K. Feeding-Habits and Distribution Pattern of 2 Forms of Flatfishes (Genus Pleuron-ich-thys) in Kiisuido Channel Waters Off the Pacific Coast of Central Japan. Nippon Suisan Gakkaishi 1992, 58, 2253–2260. [Google Scholar] [CrossRef]
- Bowman, R.E.; Stillwell, C.E.; Michaels, W.L.; Grosslein, M.D. Food of Northwest Atlantic Fishes and Two Common Species of Squid. NOAA Tech. Memo. 2000, NMFS-NE-155. [Google Scholar]
- Steimle, F. Demersal Fish and American Lobster Diets in the Lower Hudson-Raritan Estuary; US Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Northeast Region, Northeast Fisheries Science Center: Woods Hole, MA, USA, 2000.
- Paine, R.T. Trophic Relationships of 8 Sympatric Predatory Gastropods. Ecology 1963, 44, 63–73. [Google Scholar] [CrossRef]
- Rosado, M.C.; Bruxelas, A.; Castro, J.J. Ecological Effects of the Commercial Use of Diopatra neapolitana Delle Chiaje, 1841 and Solen marginatus Montagu and Structure and Dynamics of the Macrofaunal Community in a Sand Beach of the Sado Estuary (Portugal). Estud. Biol. E Conserv. Da Nat. 1993, 9, 40. [Google Scholar]
- Pires, A.; Freitas, R.; Quintino, V.; Rodrigues, A. Can Diopatra neapolitana (Annelida: Onuphidae) regenerate body damage caused by bait digging or predation? Estuar. Coast. Shelf Sci. 2012, 110, 36–42. [Google Scholar] [CrossRef]
- Merz, R.A. Textures and traction: How tube-dwelling polychaetes get a leg up. Invertebr. Biol. 2015, 134, 61–77. [Google Scholar] [CrossRef]
- Berke, S.K.; Cruz, V.; Osman, R.W. Sublethal Predation and Regeneration in Two Onuphid Polychaetes: Patterns and Impli-cations. Biol Bull. 2009, 217, 242–252. [Google Scholar] [CrossRef]
- Berke, S.K.; Mahon, A.R.; Lima, F.P.; Halanych, K.M.; Wethey, D.S.; Woodin, S.A. Range shifts and species diversity in marine ecosystem engineers: Patterns and predictions for European sedimentary habitats. Glob. Ecol. Biogeogr. 2010, 19, 223–232. [Google Scholar] [CrossRef]
- Mangum, C. Low Temperature Blockage of the Feeding Response in Boreal and Temperate Zone Polychaetes. Estuaries Coasts 1969, 1, 64–65. [Google Scholar] [CrossRef]
- Woodin, S.A.; Wethey, D.; Dubois, S.F. Population structure and spread of the polychaete Diopatra biscayensis along the French Atlantic coast: Human-assisted transport by-passes larval dispersal. Mar. Environ. Res. 2014, 102, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Galaska, M.P.; Wethey, D.S.; Arias, A.; Dubois, S.F.; Halanych, K.M.; Woodin, S.A. The impact of aquaculture on the genetics and distribution of the onuphid annelid Diopatra biscayensis. Ecol. Evol. 2021, 11, 6184–6194. [Google Scholar] [CrossRef]
- Whitney, N.M.; Wanamaker, A.D.; Ummenhofer, C.C.; Johnson, B.J.; Cresswell-Clay, N.; Kreutz, K.J. Rapid 20th century warming reverses 900-year cooling in the Gulf of Maine. Commun. Earth Environ. 2022, 3, 1–15. [Google Scholar] [CrossRef]
- Allen, M.J. Embryological Development of the Polychaetous Annelid, Diopatra cuprea (Bosc). Biol. Bull. 1959, 116, 339–361. [Google Scholar] [CrossRef]
- Lima, F.; Wethey, D. Robolimpets: Measuring intertidal body temperatures using biomimetic loggers. Limnol. Oceanogr. Methods 2009, 7, 347–353. [Google Scholar] [CrossRef]
- Smale, D.; Wernberg, T. Satellite-derived SST data as a proxy for water temperature in nearshore benthic ecology. Mar. Ecol. Prog. Ser. 2009, 387, 27–37. [Google Scholar] [CrossRef]
- Gilman, S.; Hayford, H.; Craig, C.; Carrington, E. Body temperatures of an intertidal barnacle and two whelk predators in relation to shore height, solar aspect, and microhabitat. Mar. Ecol. Prog. Ser. 2015, 536, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Judge, R.; Choi, F.; Helmuth, B. Recent Advances in Data Logging for Intertidal Ecology. Front. Ecol. Evol. 2018, 6, 213. [Google Scholar] [CrossRef] [Green Version]
- Wethey, D.S.; Woodin, S.A. Climate change and Arenicola marina: Heat waves and the southern limit of an ecosystem engineer. Estuar. Coast. Shelf Sci. 2022, 276, 10801. [Google Scholar] [CrossRef]
- Wethey, D.S.; Brin, L.D.; Helmuth, B.; Mislan, K. Predicting intertidal organism temperatures with modified land surface models. Ecol. Model. 2011, 222, 3568–3576. [Google Scholar] [CrossRef]
- Macho, G.; Woodin, S.A.; Wethey, D.S.; Vázquez, E. Impacts of Sublethal and Lethal High Temperatures on Clams Exploited in European Fisheries. J. Shellfish Res. 2016, 35, 405–419. [Google Scholar] [CrossRef]
- Freitas, F.; Cunha, T.; Hall, A.; Queiroga, H. Diopatra Neapolitana, Importância Sócio-Económica e Sustentabilidade Das Capturas, No Canal de Mira, Ria de Aveiro; Actas das Jornadas da Ria de Aveiro: Aveiro, Portugal, 2011; p. 66.
- Aleixo, A.; Queiroga, H.; Xenarios, S.; Lillebø, A. Catch Estimates and Bioecomomic Analysis of Bait Digging: The Case of the Tube Worm Diopatra neapolitana. Bioforsk Rep. 2014, 9, 1–32. [Google Scholar]
- Gurgel, C.F.D.; Norris, J.N.; Schmidt, W.E.; Le, H.N.; Fredericq, S. Systematics of the Gracilariales (Rhodophyta) including new subfamilies, tribes, subgenera, and two new genera, Agarophyton gen. nov. and Crassa gen. nov. Phytotaxa 2018, 374, 1–23. [Google Scholar]
- Kollars, N.; Byers, J.; Sotka, E. Invasive décor: An association between a native decorator worm and a non-native seaweed can be mutualistic. Mar. Ecol. Prog. Ser. 2016, 545, 135–145. [Google Scholar] [CrossRef]
Figure 1.
Summary of Diopatra ecological interactions.

Table 3.
Studies quantifying Diopatra facilitation of epibenthic fauna.
| Study | Species | Location | Community Type | Densities Tested | Effects |
|---|---|---|---|---|---|
| Mangum et al., 1968 [31] | D. cuprea | Chesapeake Bay, Virginia, USA | Epifauna | Single tubes | Tubes support 49 species of Arthropoda, Mollusca, Annelida, and other invertebrate phyla. |
| Bell & Coen 1982 [9] | D. cuprea | Tampa Bay, FL | Meiofauna | Single tubes | Tubes support crustacean nauplii, copepods, and nematodes (tens to hundreds per tube). Polychaetes also found on tubes. |
| Bell & Coen 1982 [72] | D. cuprea | Tom’s Cove, Virginia, USA | Meiofauna | 1 or 4 · 0.01 m−2 | Tubes support nauplii, copepods, amphipods, ostracods, nematodes, and polychaetes. Abundances increase when algae are also present, with no effect of tube density. |
| Dudley et al., 1989 [74] | D. ornata | Venado Beach, Panama | Epifaunal mollusca | Single tubes | Tubes support seven species of gastropod, four bivalves, and a chiton. |
| Dudley et al., 1989 [74] | D. cuprea | Virginia, USA and Woods Hole, Massachusetts, USA | Epifaunal mollusca | Single tubes | Tubes support five species of gastropod and four bivalves. |
| Diaz et al., 2003 [10] | D. cuprea | Mid-Atlantic Bight | Juvenile epibenthic fish | “mat” | Juvenile fish were twice as abundant in tube mats as bare sand during the day (reversed at night). Eight species of fish were associated with D. cuprea tubes. |
| Thomsen et al., 2011 [23] | Diopatra-like onuphid * | Inhaca Island, Mozambique | Epifauna | Single tubes | ↑ richness and abundance on single tubes |
| Santos and Aviz 2018 [20] † | D. cuprea * | Algodoal-Maiandeua, Brazil | Epifauna and Infauna | Single tubes | Seasonal ↑ richness and abundance |
* Species in this region have been recently revised, † Sampling captured both infauna and the tube-cap, making it impossible to separate infauna from epifauna.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Berke, S.K. A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete. Biology 2022, 11, 1485. https://doi.org/10.3390/biology11101485
AMA Style
Berke SK. A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete. Biology. 2022; 11(10):1485. https://doi.org/10.3390/biology11101485
Chicago/Turabian StyleBerke, Sarah K. 2022. "A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete" Biology 11, no. 10: 1485. https://doi.org/10.3390/biology11101485
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.