
Preliminary Data about Habitat Use of Subadult and Adult White Sharks (Carcharodon carcharias) in Eastern Australian Waters


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
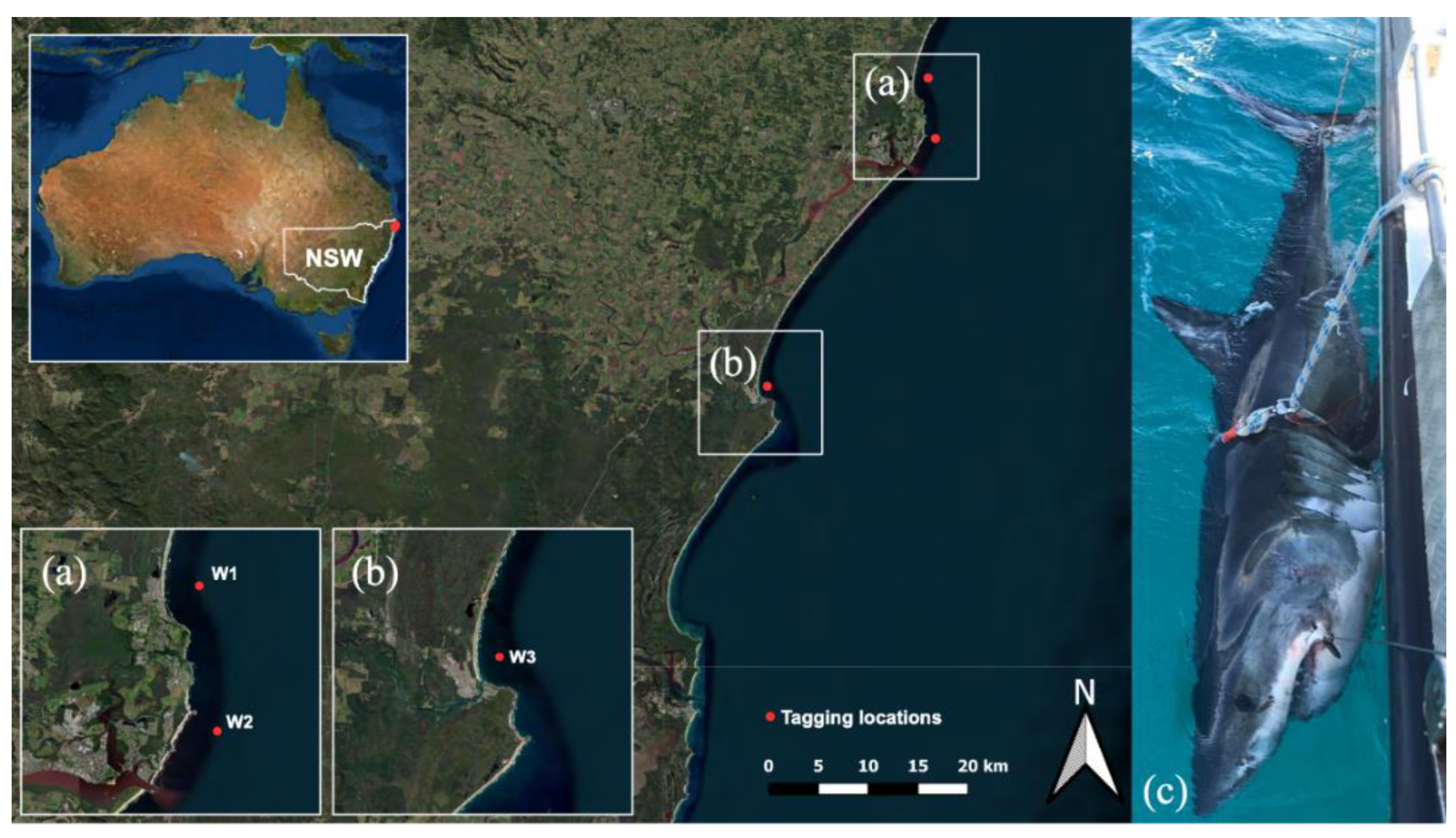
2.1. Shark Capture and Fishing Gear
2.2. Tagging Method
2.3. Tag Details and Programming
2.4. Track Reconstruction and Data Analysis
3. Results
3.1. Tag Success and Deployment Summarys
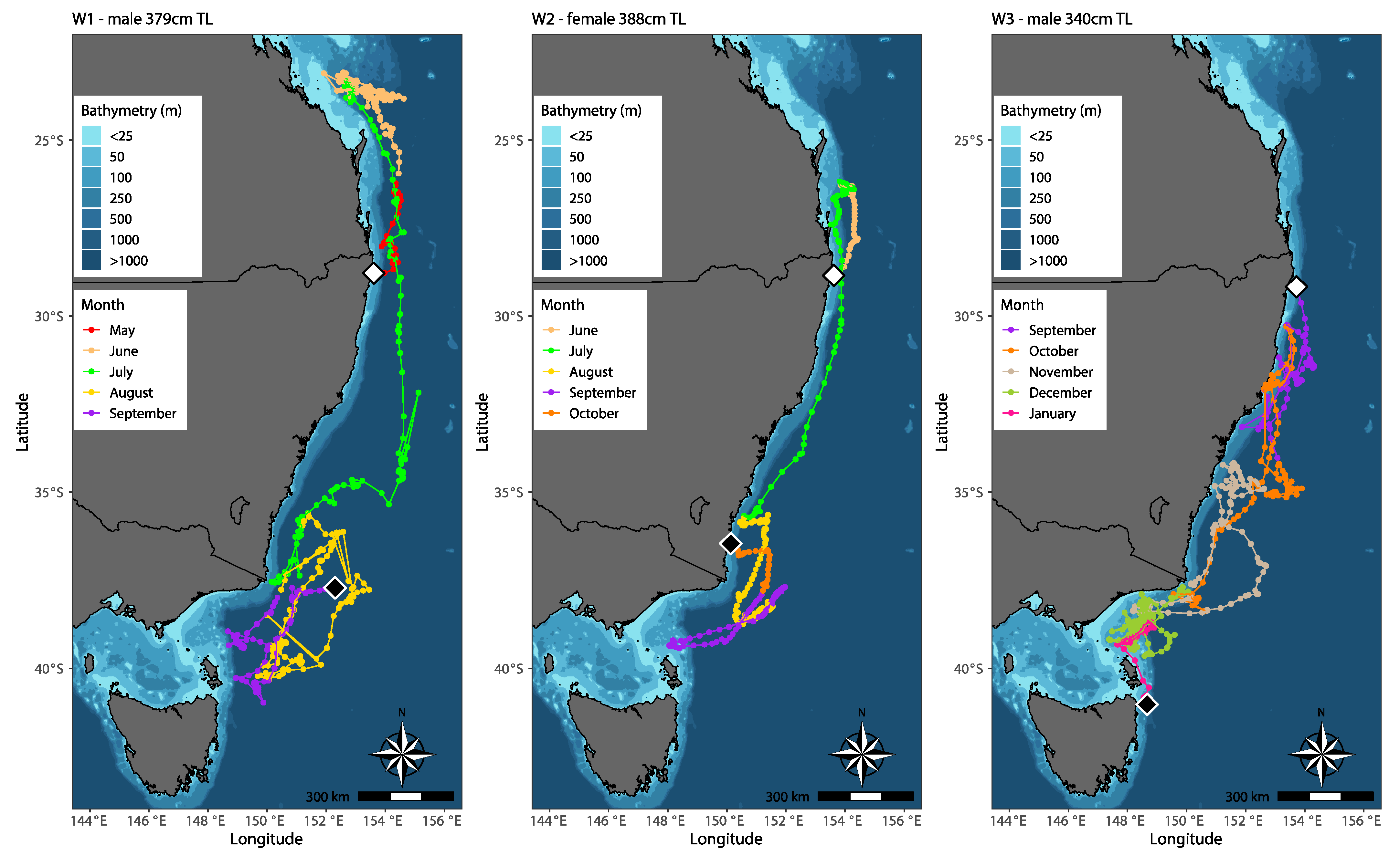
3.2. Horizontal Movement and Behaviour
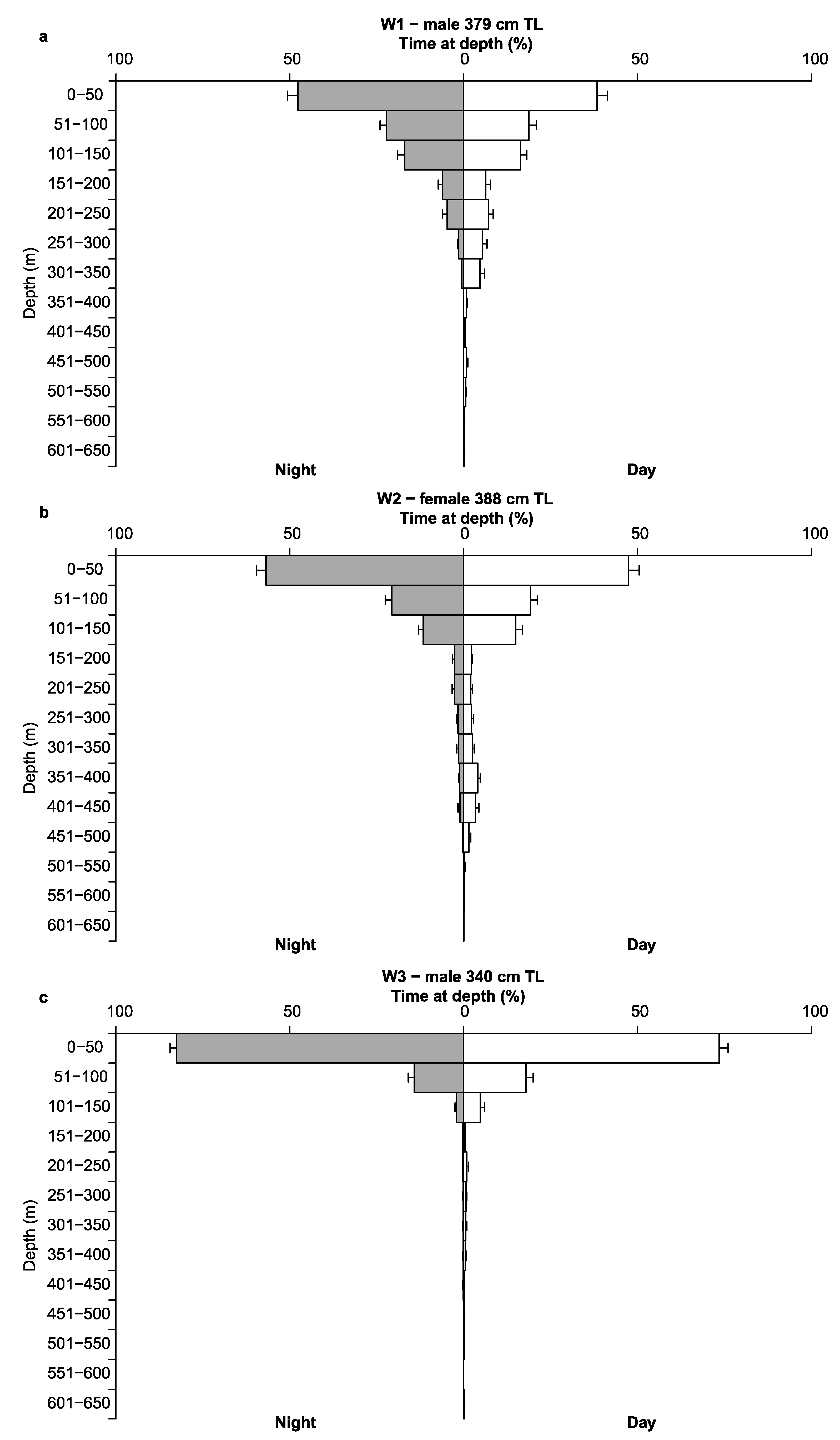
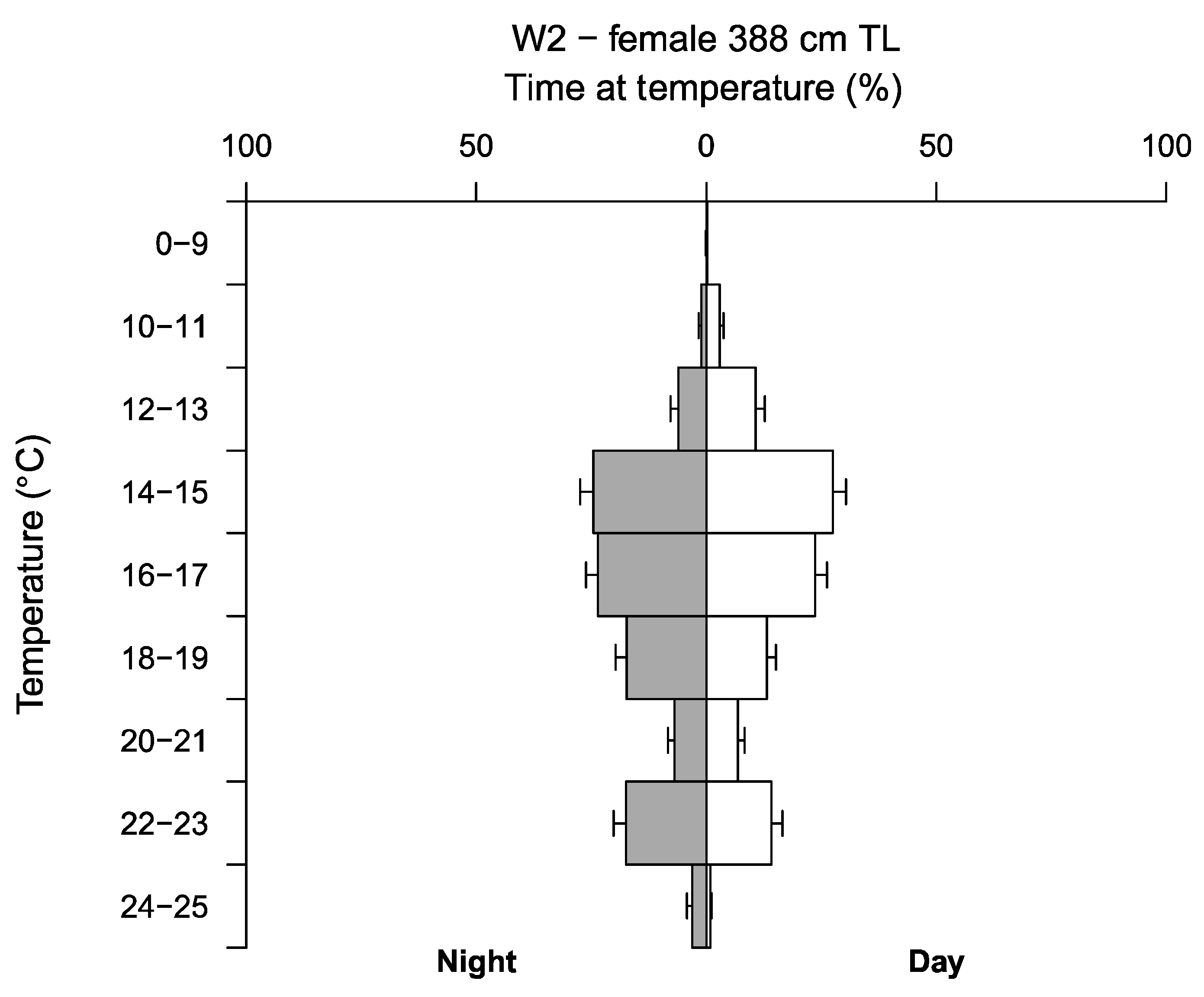
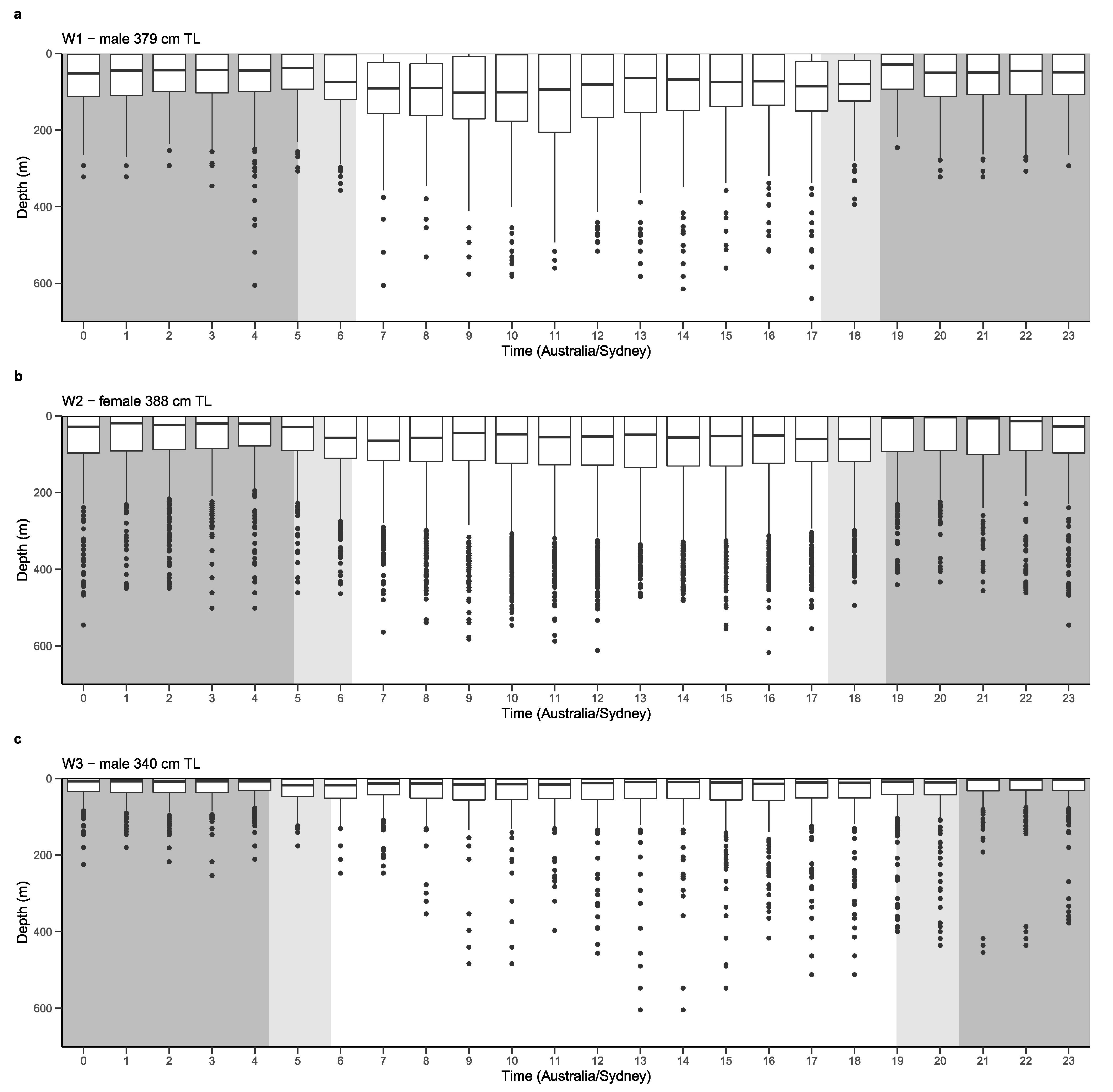
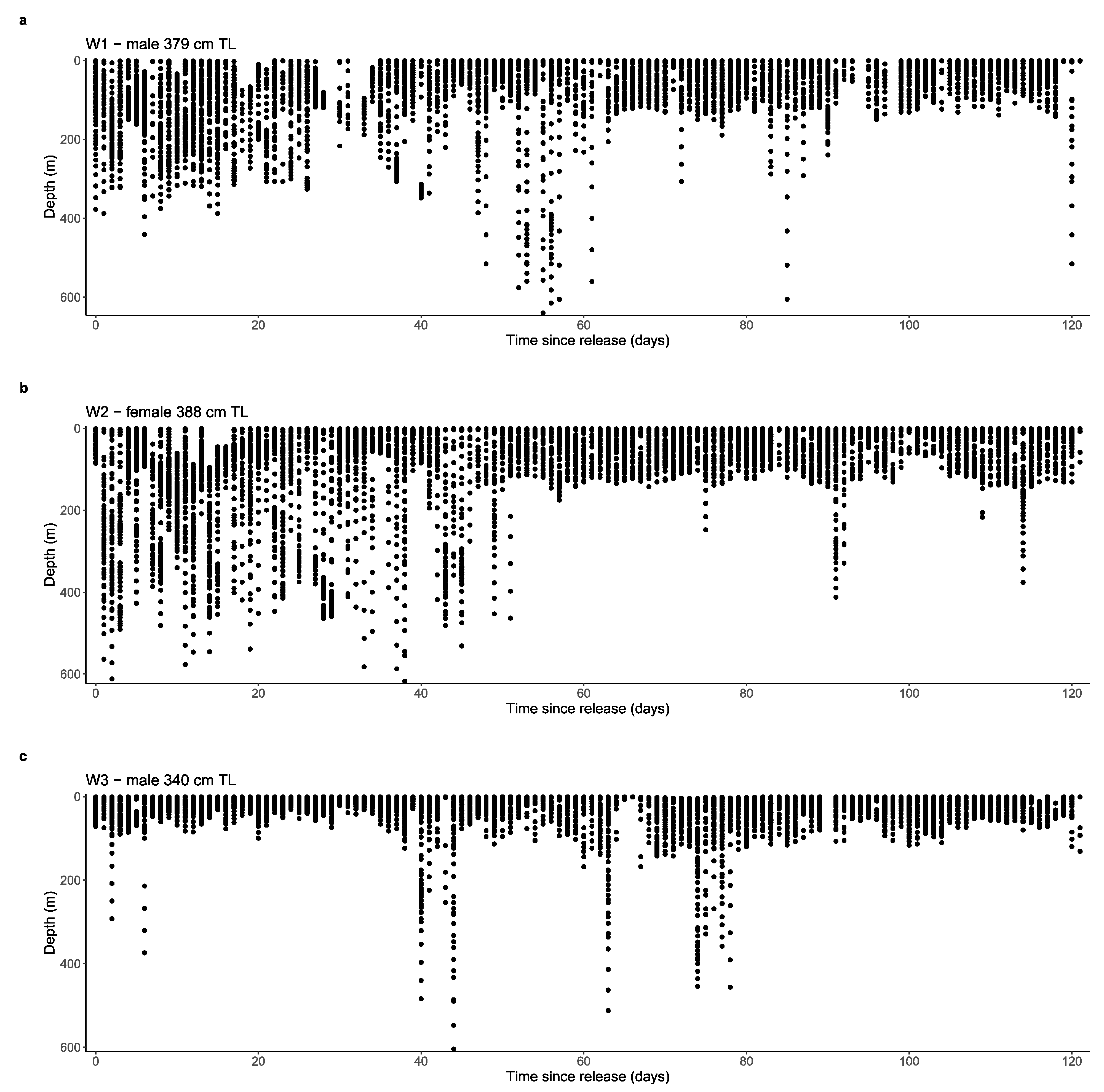
3.3. Depth, Temperature and Vertical Diel Habitat Use
4. Discussion
4.1. Horizontal Movement
4.1.1. Longitudinal Movement
4.1.2. Latitudinal Movement
4.2. Resident Behavior
4.3. Depth and Temperature Preference
4.4. Vertical Habitat Use and Diving Behaviour
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruce, B.D.; Stevens, J.D.; Malcolm, H. Movements and swimming behaviour of white sharks (Carcharodon carcharias) in Australian waters. Mar. Biol. 2006, 150, 161–172. [Google Scholar] [CrossRef]
- Weng, K.C.; Boustany, A.M.; Pyle, P.; Anderson, S.D.; Brown, A.; Block, B.A. Migration and habitat of white sharks (Carcharodon carcharias) in the eastern Pacific Ocean. Mar. Biol. 2007, 152, 877–894. [Google Scholar] [CrossRef]
- Ferretti, F.; Worm, B.; Britten, G.L.; Heithaus, M.R.; Lotze, H.K. Patterns and ecosystem consequences of shark declines in the ocean. Ecol. Lett. 2010, 13, 1055–1071. [Google Scholar] [CrossRef] [PubMed]
- Skomal, G.B.; Braun, C.D.; Chisholm, J.H.; Thorrold, S.R. Movements of the white shark Carcharodon carcharias in the North Atlantic Ocean. Mar. Ecol. Prog. Ser. 2017, 580, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hammerschlag, N.; Williams, L.; Fallows, M.; Fallows, C. Disappearance of white sharks leads to novel emergence of and allopatric apex predator, the sevengill shark. Sci. Reps. 2018, 9, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Springer, V.G.; Last, P.R.; Stevens, J.D. Sharks and Rays of Australia. Copeia 1994, 1055. [Google Scholar] [CrossRef] [Green Version]
- Bradford, R.; Patterson, T.A.; Rogers, P.J.; McAuley, R.; Mountford, S.; Huveneers, C.; Robbins, R.; Fox, A.; Bruce, B.D. Evidence of diverse movement strategies and habitat use by white sharks, Carcharodon carcharias, off southern Australia. Mar. Biol. 2020, 167, 1–12. [Google Scholar] [CrossRef]
- Spaet, J.L.Y.; Manica, A.; Brand, C.P.; Gallen, C.; Butcher, P.A. Environmental conditions are poor predictors of im-mature white shark Carchardon carcharias occurrences on coastal beaches of eastern Australia. Mar. Ecol. Prog. Series 2020, 653, 167–179. [Google Scholar] [CrossRef]
- Blower, D.C.; Pandolfi, J.M.; Bruce, B.D.; Gomez-Cabrera, M.D.C.; Ovenden, J.R. Population genetics of Australian white sharks reveals fine-scale spatial structure, transoceanic dispersal events and low effective population sizes. Mar. Ecol. Prog. Ser. 2012, 455, 229–244. [Google Scholar] [CrossRef] [Green Version]
- McAuley, R.B.; Bruce, B.D.; Keay, I.S.; Mountford, S.; Pinnell, T.; Whoriskey, F.G. Broad-scale coastal movements of white sharks off Western Australia described by passive acoustic telemetry data. Mar. Freshw. Res. 2017, 68, 1518. [Google Scholar] [CrossRef]
- Hillary, R.M.; Bravington, M.V.; Patterson, T.A.; Grewe, P.; Bradford, R.; Feutry, P.; Gunasekera, R.; Peddemors, V.; Werry, J.; Francis, M.P.; et al. Genetic relatedness reveals total population size of white sharks in eastern Australia and New Zealand. Sci. Rep. 2018, 8, 2661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, B.; Harasti, D.; Lee, K.; Gallen, C.; Bradford, R. Broad-scale movements of juvenile white sharks Carcharodon carcharias in eastern Australia from acoustic and satellite telemetry. Mar. Ecol. Prog. Ser. 2019, 619, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Spaet, J.L.Y.; Patterson, T.A.; Bradford, R.W.; Butcher, P.A. Spatiotemporal distribution patterns of immature Australasian white sharks (Carcharodon carcharias). Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Bonfil, R.; Meyer, M.; Scholl, M.C.; Johnson, R.; O’Brien, S.; Oosthuizen, H.; Swanson, S.; Kotze, D.; Paterson, M. Transoceanic Migration, Spatial Dynamics, and Population Linkages of White Sharks. Science 2005, 310, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Bruce, B.D.; Bradford, R.W. Habitat Use and Spatial Dynamic of Juvenile White Sharks, Carcharodon Carcharias, in Eastern Australia. In Global Perspectives on the Biology and Life History of the White Shark, 1st ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 250–279. [Google Scholar]
- Bradford, R.W.; Hobday, A.J.; Bruce, B.D. Identifying juvenile white shark behaviour from electronic tag data. In Global Perspectives on the Biology and Life History of the White Shark, 1st ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 255–270. [Google Scholar]
- Domeier, M.L.; Nasby-Lucas, N. Two-year migration of adult female white sharks (Carcharodon carcharias) reveals widely separated nursery areas and conservation concerns. Anim. Biotelemetry 2013, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Huveneers, C.; Apps, K.; Becerril-García, E.; Bruce, B.; Butcher, P.; Carlisle, A.; Chapple, T.; Christiansen, H.; Cliff, G.; Curtis, T.; et al. Future research directions on the “Elusive” White Shark. Front. Mar. Science. 2018, 5, 108–115. [Google Scholar] [CrossRef]
- Werry, J.M.; Bruce, B.D.; Sumpton, W.; Reid, D.; Mayer, D.G. Beach areas used by juvenile white sharks, Carcharodon carcharias, in eastern Australia. In Global Perspectives on the Biology and Life History of the White Shark, 1st ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 271–287. [Google Scholar]
- Reid, D.D.; Krogh, M. Assessment of catches from protective shark meshing off New South Wales beaches between 1950 and 1990. Aust. J. Mar. Freshw. Research. 1992, 43, 283–296. [Google Scholar] [CrossRef]
- Butcher, P.A.; Piddocke, T.P.; Colefax, A.P.; Hoade, B.; Peddemors, V.M.; Borg, L.; Cullis, B.R. Beach safety: Can drones provide a platform for sighting sharks? Wildl. Res. 2019, 46, 701. [Google Scholar] [CrossRef] [Green Version]
- Tate, R.D.; Cullis, B.R.; Smith, S.D.A.; Kelaher, B.P.; Brand, C.P.; Gallen, C.R.; Mandelman, J.W.; Butcher, P.A. The acute physiological status of white sharks (Carcharodon carcharias) exhibits minimal variation after capture on SMART drum-lines. Conserv. Physiol. 2019, 7, 1–9. [Google Scholar]
- Colefax, A.P.; Kelaher, B.P.; Pagendam, D.E.; Butcher, P.A. Assessing white shark (Carcharodon carcharias) behaviour along coastal beaches for conservation-focused shark mitigation. Front. Mar. Sci. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Tate, R.D.; Kelaher, B.P.; Brand, C.P.; Cullis, B.R.; Gallen, C.R.; Smith, S.D.A.; Butcher, P.A. The effective of Shark-Management-Alert-in-Real-Time (SMART) drumlines as a tool for catching white sharks, Carcharodon carcharias, off coastal New South Wales, Australia. Fish. Manag. Ecology. 2021, 28, 496–506. [Google Scholar] [CrossRef]
- Bruce, B.; Bradford, R.W.; Bravington, M.; Feutry, P.; Grewe, P.; Gunasekera, R.; Harasti, D.; Hillary, R.M.; Patterson, T.A. A National Assessment of the Status of White Sharks; National Environmental Science Program Marine Biodiversity Hub: Hobart, Australia, 2018; Available online: https://www.nespmarine.edu.au/document/national-assessment-status-white-sharks (accessed on 1 December 2021).
- Guyomard, D.; Perry, C.; Tournoux, P.U.; Cliff, G.; Peddemors, V.; Jaquemet, S. An innovative fishing gear to enhance the release of non-target species in coastal shark-control programs: The SMART (shark management alert in real-time) drumline. Fish. Res. 2019, 216, 6–17. [Google Scholar] [CrossRef]
- Domeier, M.L.; Kiefer, D.; Nasby-Lucas, N.; Wagschal, A.; O’Brien, F. Tracking Pacific bluefin tuna (Thunnus thynnus orientalis) in the northeastern Pacific with an automated algorithm hat estimates latitudes by matching sea-surface-temperature data from satellites with temperature data from tags on fish. Fish. Bulletin. 2005, 2, 292–306. [Google Scholar]
- Wildlife Computers. Location Processing (GPE3 and Fastloc GPS®) in the Wildlife Computers Data Portal User Guide. 2020. Available online: https://static.wildlifecomputers.com/Location-Processing-UserGuide.pdf (accessed on 12 December 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 29 June 2021).
- Bauer, R.K.; Forget, F.; Fromentin, J.M. Optimising PAT data transmission: Assessing the accuracy of temperature summary data to estimate environmental conditions. Fish. Oceanograph. 2015, 24, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; London, J. Crawl: An R Package for Fitting Continuous-Cime Correlated Random Walk Models to Animal Movement Data. Zenodo; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar] [CrossRef]
- Pante, E.; Simon-Bouhet, B. Marmap: A Package for Importing, Plotting and Analyzing Bathymetric and Topographic Data in R. PLoS ONE 2013, 8, e73051. [Google Scholar] [CrossRef]
- Wickham, H. Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Spaet, J.L.Y.; Butcher, P.A.; Manica, A.; Lam, C.H. Spatial dynamics and fine-scale vertical behaviour of immature eastern Australasian white sharks (Carcharodon carcharias). J. Biol. (submitted).
- Lipscombe, R.S.; Spaet, J.L.Y.; Scott, A.; Lam, C.H.; Brand, C.P.; A Butcher, P. Habitat use and movement patterns of tiger sharks (Galeocerdo cuvier) in eastern Australian waters. ICES J. Mar. Sci. 2020, 77, 3127–3137. [Google Scholar] [CrossRef]
- Barnes, C.; Butcher, P.; Macbeth, W.; Mandelman, J.; Smith, S.; Peddemors, V. Movements and mortality of two commercially exploited carcharhinid sharks following longline capture and release off eastern Australia. Endanger. Species Res. 2016, 30, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Boustany, A.M.; Davis, S.F.; Pyle, P.; Anderson, S.D.; Le Boeuf, B.J.; Block, B.A. Expanded niche for white sharks. Nature 2002, 415, 35–36. [Google Scholar] [CrossRef]
- Gallagher, A.J.; Meyer, L.; Pethybridge, H.R.; Huveneers, C.; Butcher, P.A. Effects of short-term capture on the physi-ology of white sharks Carcharodon carcharias: Amino acids and fatty acids. Endanger. Species Res. 2019, 40, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Domeier, M.L.; Nasby-Lucas, N. Sex-specific migration patterns and sexual segregation of adult white sharks, Car-charodon carcharias, in the northeastern Pacific. In Global Perspectives on the Biology and Life History of the White Shark, 1st ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 133–146. [Google Scholar]
- Duffy, C.A.J.; Francis, M.P.; Manning, M.J.; Bonfil, R. Regional population connectivity, oceanic habitat, and return migra-tion revealed by satellite tagging of white sharks, Carcharodon carcharias, at New Zealand aggregation sites. In Global Perspectives on the Biology and Life History of the White Shark, 1st ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 326–343. [Google Scholar]
- Ridgeway, K.; Hill, K.L. The east Australian current. In A Marine Climate Change Impacts and Adaptation Report Card for Australia; Poloczanska, E.S., Hobday, A.J., Richardson, A.J., Eds.; NCCARF Publication: Southport, Australia, 2009; pp. 1–16. [Google Scholar]
- Everett, J.D.; Baird, M.E.; Oke, P.R.; Suthers, I.M. An avenue of eddies: Quantifying the biophysical properties of mesoscale eddies in the Tasman Sea. Geophys. Res. Lett. 2012, 39, 3091. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Butcher, P.; Harcourt, R.; Patterson, T.; Peddemors, V.; Roughan, M.; Harasti, D.; Smoothey, A.; Bradford, R. Oceanographic conditions associated with white shark (Carcharodon carcharias) habitat use along eastern Australia. Mar. Ecol. Prog. Ser. 2021, 659, 143–159. [Google Scholar] [CrossRef]
- Braun, C.D.; Arostegui, M.C.; Thorrold, S.R.; Papastamatiou, Y.P.; Gaube, P.; Fontes, J.; Afonso, P. The Functional and Ecological Significance of Deep Diving by Large Marine Predators. Annu. Rev. Mar. Sci. 2022, 14, 129–159. [Google Scholar] [CrossRef]
- Klimley, A.P. Highly directional swimming by scalloped hammerhead sharks, Sphyrna lewini, and subsurface irradiance, temperature, bathymetry, and geomagnetic field. Mar. Biol. 1993, 117, 1–22. [Google Scholar] [CrossRef]
- Malcolm, H.; Bruce, B.D.; Stevens, J.D. A Review of the Biology and Status of White Sharks in Australian Waters; CSIRO Division of Marine Research: Hobart, Australia, 2001. [Google Scholar]
- Clark, Z.; Fish, J.; Butcher, P.; Sherman, C.; Weeks, A.; Miller, A. DNA metabacoding provides new insights into white shark diet and feeding behaviours in eastern Australia. J. Biol. (submitted).
- Francis, M.P.; Duffy, C.A.J.; Bonfil, R.; Manning, M.J. The third dimension: Vertical habitat use by white sharks, Carcharodon carcharias, in New Zealand and in oceanic tropical waters of the southwest Pacific Ocean. In Global Perspectives on the Biology and Life History of the White Shark, 1st ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 319–342. [Google Scholar]
- Weltz, K.; Kock, A.A.; Winker, H.; Attwood, C.; Sikweyiya, M. The influence of environmental variables on the presence of white sharks, Carcharodon carcharias at two popular Cape Town bathing beaches: A generalised additive mixed model. PLoS ONE 2013, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, K.J. Regulation of body temperature in the white shark, Carcharodon carcharias. J. Comp. Physiol. B 1997, 167, 423–429. [Google Scholar] [CrossRef]
- Arrowsmith, L.M.; Sequeria, A.M.M.; Pattiaratchi, C.B.; Meekan, M.G. Water temperature is a key driver of horizontal and vertical movements of an ocean giant, the whale shark Rhicodon typus. Mar. Ecol. Prog. Ser. 2021, 679, 101–114. [Google Scholar] [CrossRef]
- Santos, C.C.; Domingo, A.; Carlson, J.; Natanson, L.; Travassos, P.; Macias, D.; Cortés, E.; Miller, P.; Hazin, F.; Mas, F.; et al. Updates on habitat use and migrations of shortfin mako in the Atlantic using satellite telemetry. Collect. Vol. Sci. Papers. 2020, 75, 445–456. [Google Scholar]
- Huang, B.; Thorne, P.W.; Banzon, V.F.; Boyer, T.; Chepurin, G.; Lawrimore, J.H.; Menne, M.J.; Smith, T.M.; Vose, R.S.; Zhang, H.-M. Extended Reconstructed Sea Surface Temperature, Version 5 (ERSSTv5): Upgrades, Validations, and Intercomparisons. J. Clim. 2017, 30, 8179–8205. [Google Scholar] [CrossRef]
- Bonfil, R.; Francis, M.P.; Duffy, C.; Manning, M.J.; O’Brien, S. Large-scale tropical movements and diving behaviour of white sharks Carcharodon carcharias tagged off New Zealand. Aquat. Biol. 2009, 8, 115–123. [Google Scholar] [CrossRef]
- Kim, S.L.; Tinker, M.T.; Estes, J.A.; Koch, P.L. Ontogenetic and Among-Individual Variation in Foraging Strategies of Northeast Pacific White Sharks Based on Stable Isotope Analysis. PLoS ONE 2012, 7, e45068. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, P.J.; Huveneers, C.; Page, B.; Goldsworthy, S.D.; Coyne, M.; Lowther, A.D.; Mitchell, J.G.; Seuront, L. Living on the continental shelf edge: Habitat use of juvenile shortfin mako Isurus oxyrinchus in the Great Australian Bight, southern Australia. Fish. Oceanography 2015, 24, 205–218. [Google Scholar] [CrossRef]
- Carey, F.G.; Kanwisher, J.W.; Brazier, O.; Gabrielson, G.; Casey, J.G.; Pratt, H.L. Temperature and Activities of a White Shark, Carcharodon carcharias. Copeia 1982, 1982, 254. [Google Scholar] [CrossRef]
- McPhee, D. Unprovoked shark bites: Are they becoming more prevalent? Coast. Manag. 2014, 42, 478–492. [Google Scholar] [CrossRef]
- Chapman, B.K.; McPhee, D. Global shark attack hotspots: Identifying underlying factors behind increased unprovoked shark bite incidence. Ocean Coast. Manag. 2016, 133, 72–84. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sex | Total Length (cm) | Tagging Date | Tagging Location Lat. (°S), Long. (°E) | Date Tag Detached | Pop-Up Location Lat. (°S), Long. (°E) | Days Tracked | Estimated Track Length (km) | Estimated Average Movement (km day−1) |
|---|---|---|---|---|---|---|---|---|---|
| W1 | M | 379 | 23 May 2021 | 28.783, 153.610 | 20 September 2021 | 37.721, 152.300 | 120 | 10,420 | 86.83 |
| W2 * | F | 388 | 10 June 2021 | 28.845, 153.617 | 8 October 2021 | 36.468, 150.133 | 120 | 6243 | 52.02 |
| W3 | M | 340 | 10 September 2021 | 29.167, 153.717 | 9 January 2022 | 41.026, 148.666 | 120 | 9620 | 80.16 |
| Depth (m) | Temperature (°C) | |||
|---|---|---|---|---|
| Shark ID | Min-Max | Mean ± SE | Min-Max | Mean ± SE |
| W1 | 0.50–640.00 | 84.77 ± 0.60 | - | - |
| W2 * | 0.00–644.50 | 79.20 ± 0.60 | 7.70–24.05 | 16.79 ± 0.002 |
| W3 | 0.50–604.50 | 32.59 ± 0.38 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coxon, J.L.; Butcher, P.A.; Spaet, J.L.Y.; Rizzari, J.R. Preliminary Data about Habitat Use of Subadult and Adult White Sharks (Carcharodon carcharias) in Eastern Australian Waters. Biology 2022, 11, 1443. https://doi.org/10.3390/biology11101443
Coxon JL, Butcher PA, Spaet JLY, Rizzari JR. Preliminary Data about Habitat Use of Subadult and Adult White Sharks (Carcharodon carcharias) in Eastern Australian Waters. Biology. 2022; 11(10):1443. https://doi.org/10.3390/biology11101443
Chicago/Turabian StyleCoxon, Jessica L., Paul A. Butcher, Julia L. Y. Spaet, and Justin R. Rizzari. 2022. "Preliminary Data about Habitat Use of Subadult and Adult White Sharks (Carcharodon carcharias) in Eastern Australian Waters" Biology 11, no. 10: 1443. https://doi.org/10.3390/biology11101443