Zoonotic Visceral Leishmaniasis: New Insights on Innate Immune Response by Blood Macrophages and Liver Kupffer Cells to Leishmania infantum Parasites

 ,
,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Kupffer Cell Purification and Culture
2.2. Separation of Peripheral Blood Monocytes and Differentiation to Macrophages
2.3. L. infantum Parasites
2.4. L. infantum Infection of Macrophage and Kupffer Cells
2.5. Scanning Electron Microscopy (SEM)
2.6. Evaluation of Cell-Parasite Interaction by Microscopy
2.7. Nitrite Detection
2.8. Real Time-PCR Analysis
2.9. Statistical Analysis
2.10. Ethical Considerations
3. Results
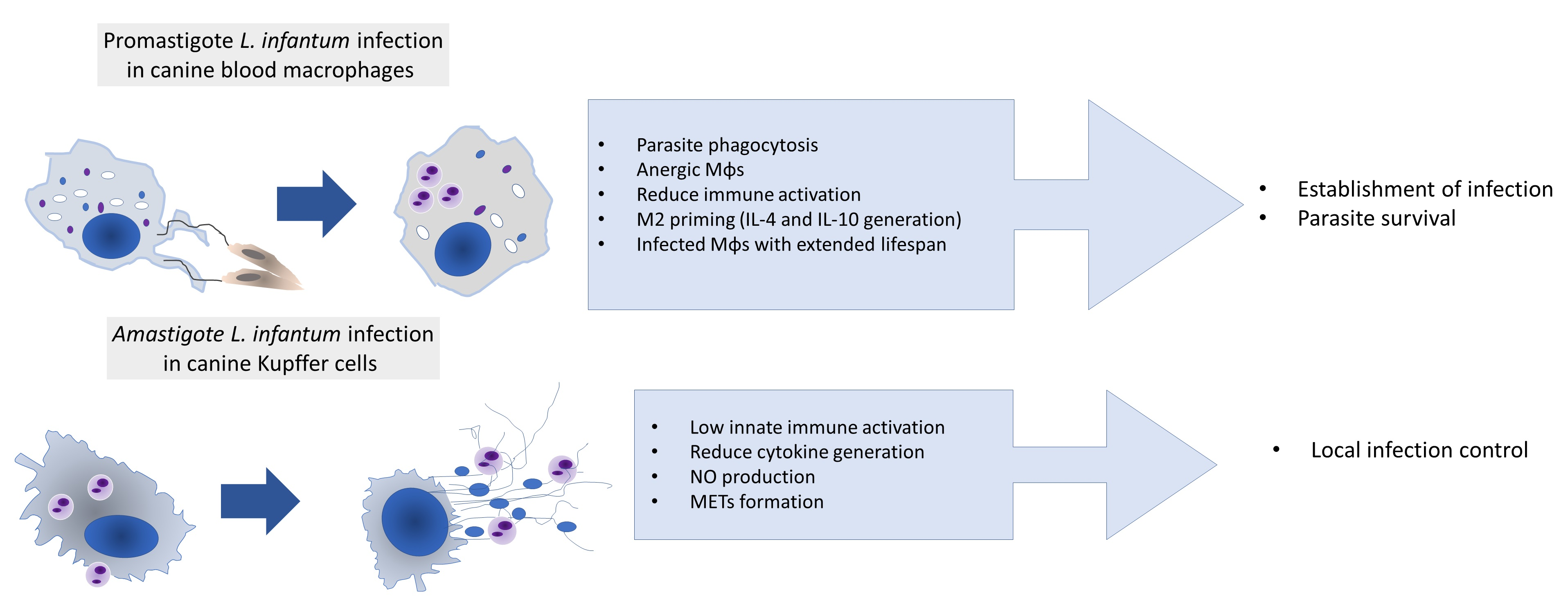
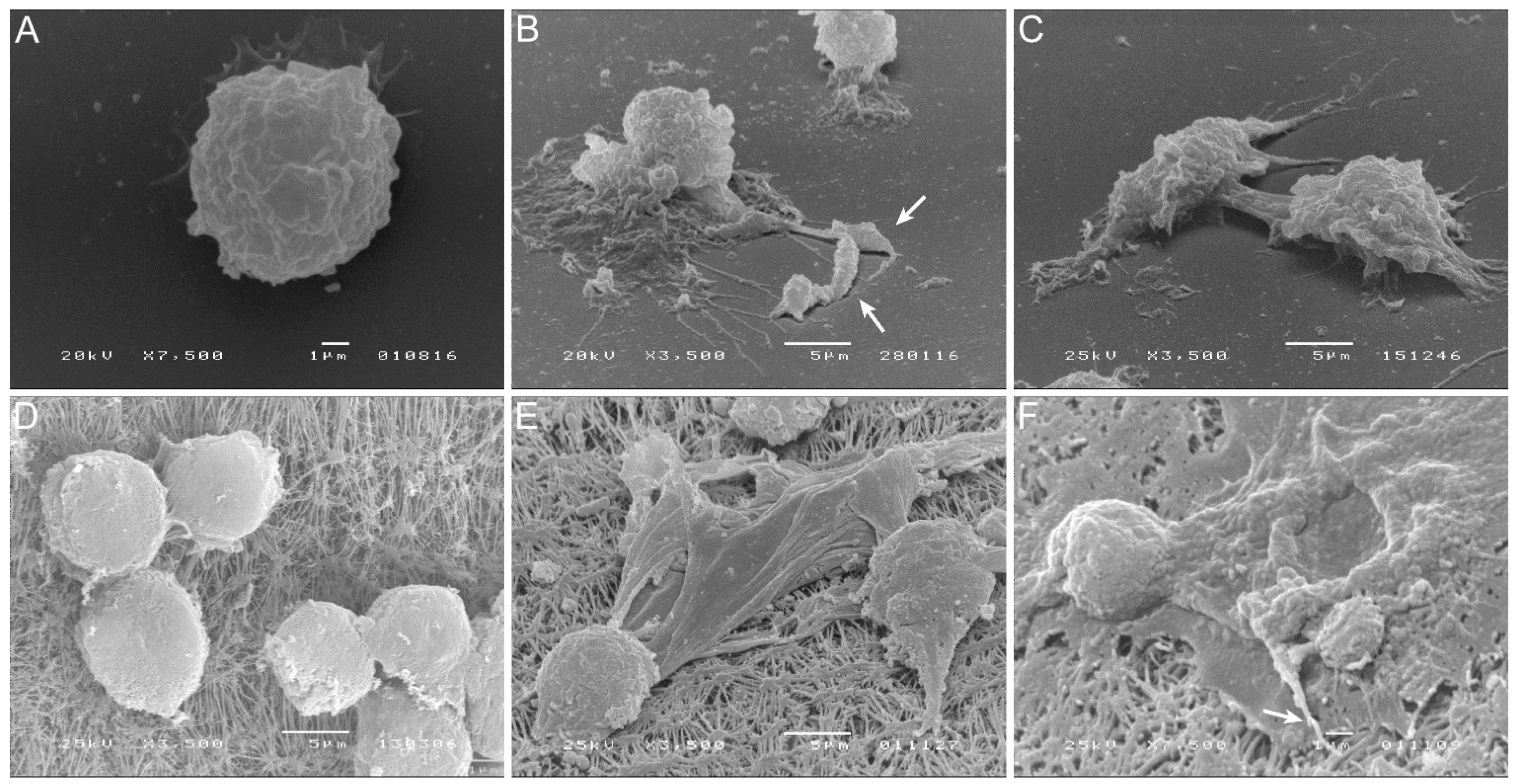
3.1. Axenic L. infantum Amastigotes Can Infect KCs
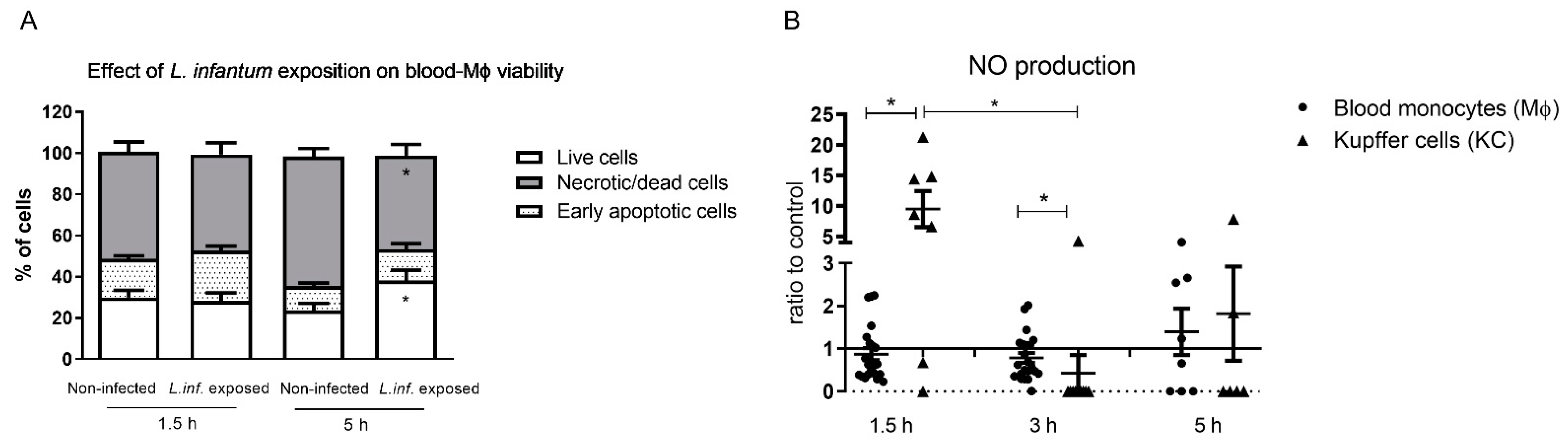
3.2. L. infantum Infected Blood-MØs Have Increased Lifespan
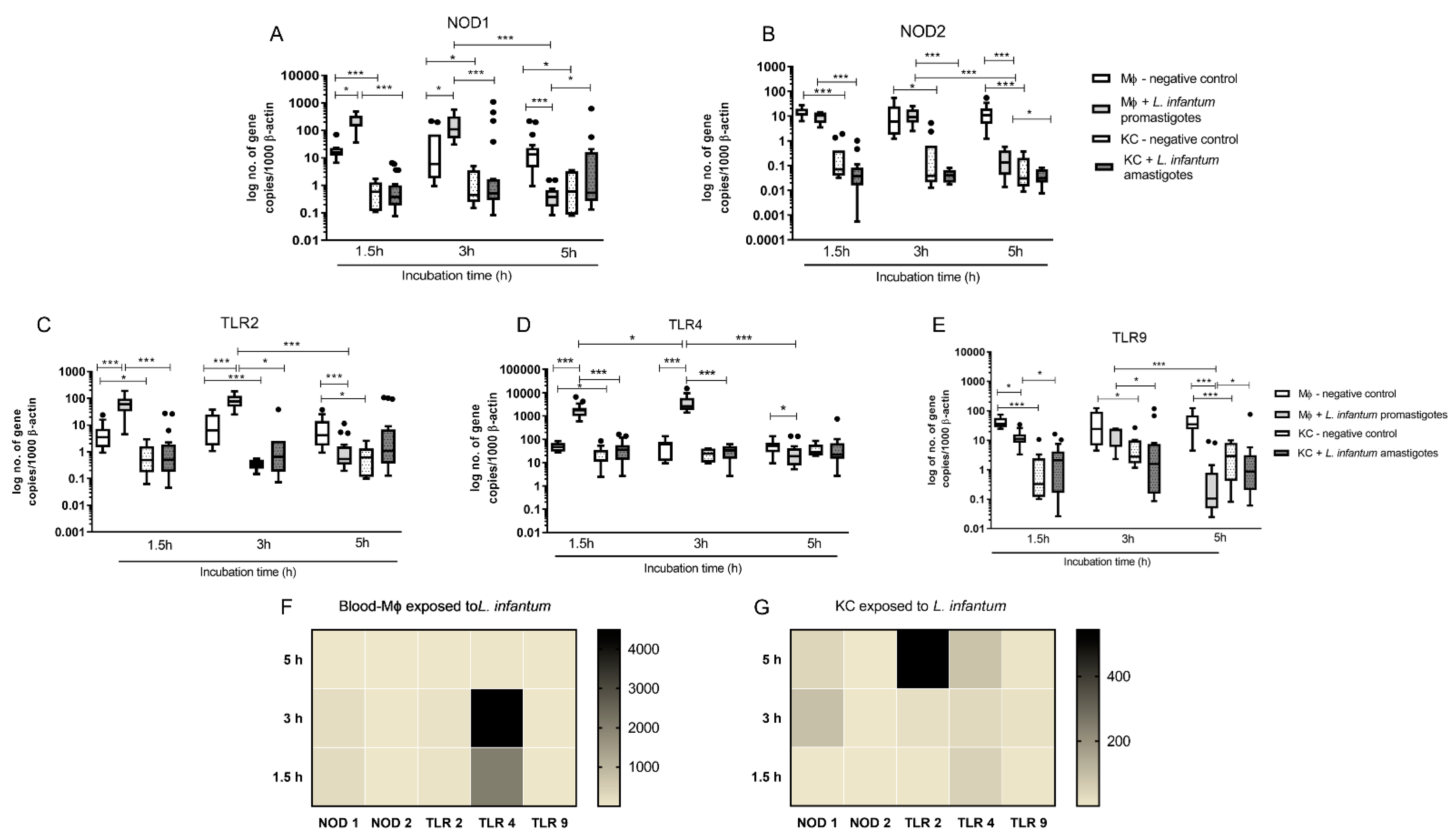
3.3. Infected Blood-MØs Display a Peak of PRR Gene Expression in Response to L. infantum
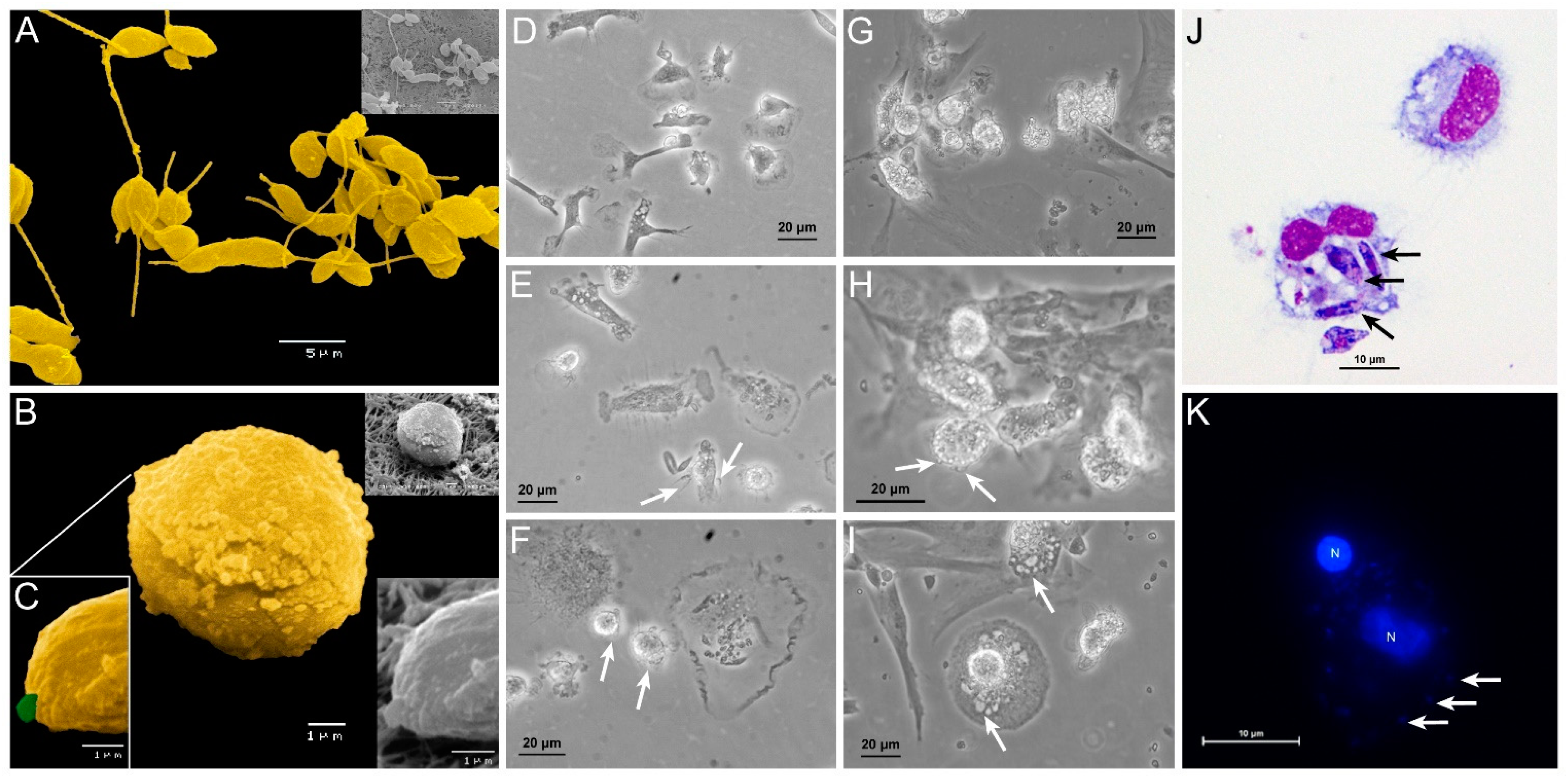
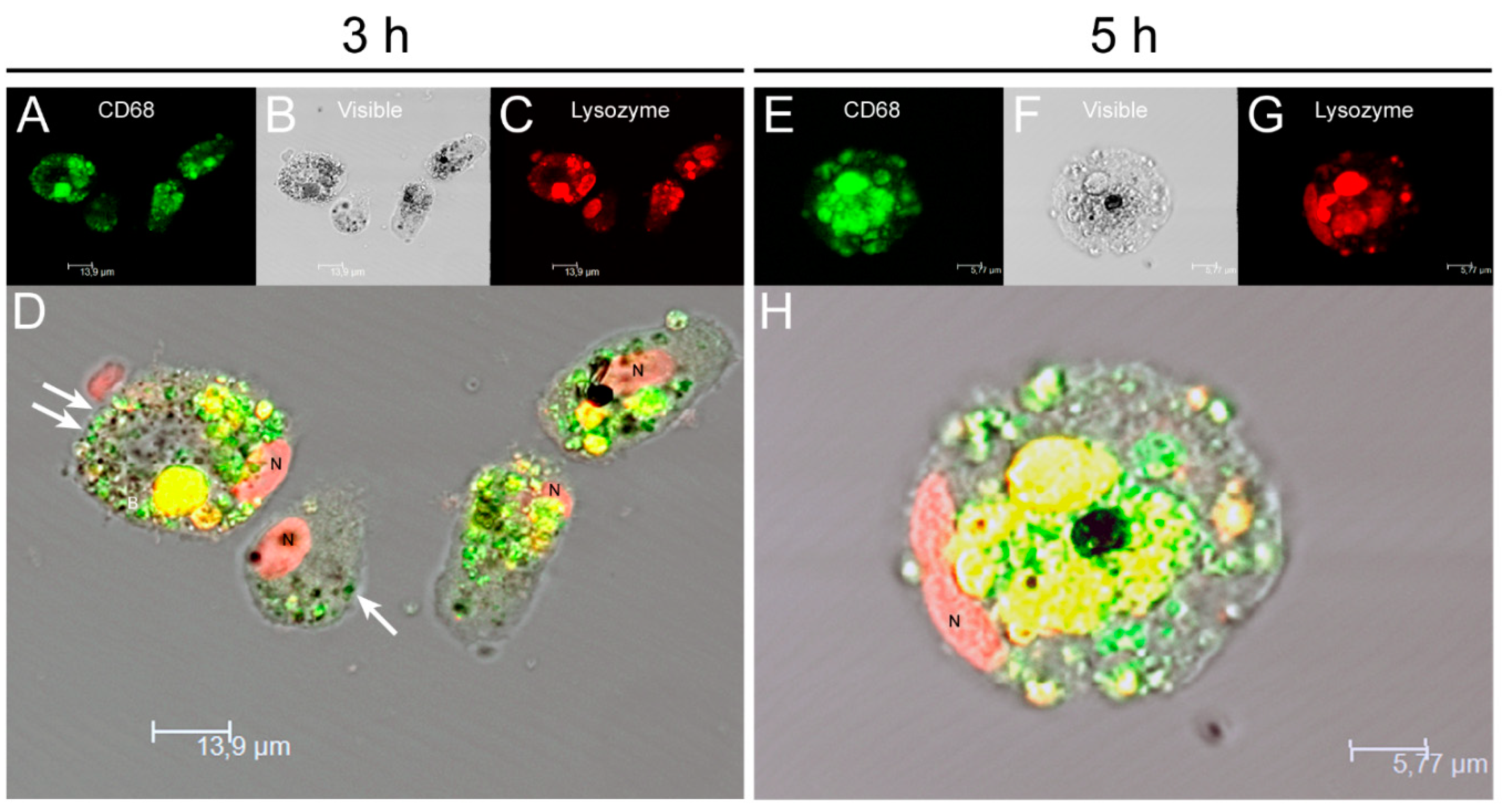
3.4. Blood-MØs Phagocyte Parasites While KCs Release Extracellular Traps
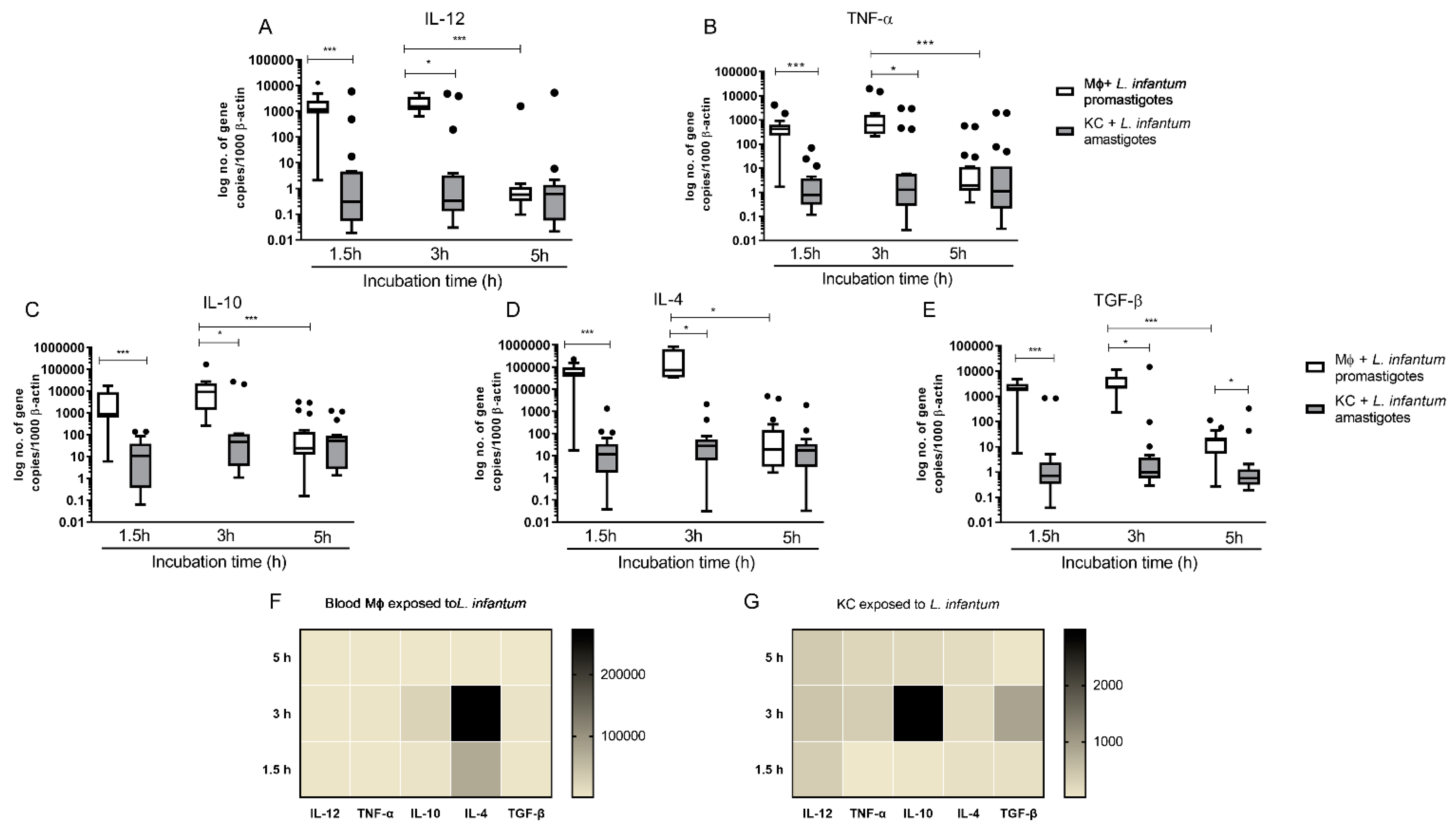
3.5. L. infantum Primes Blood-MØs for M2 Phenotype
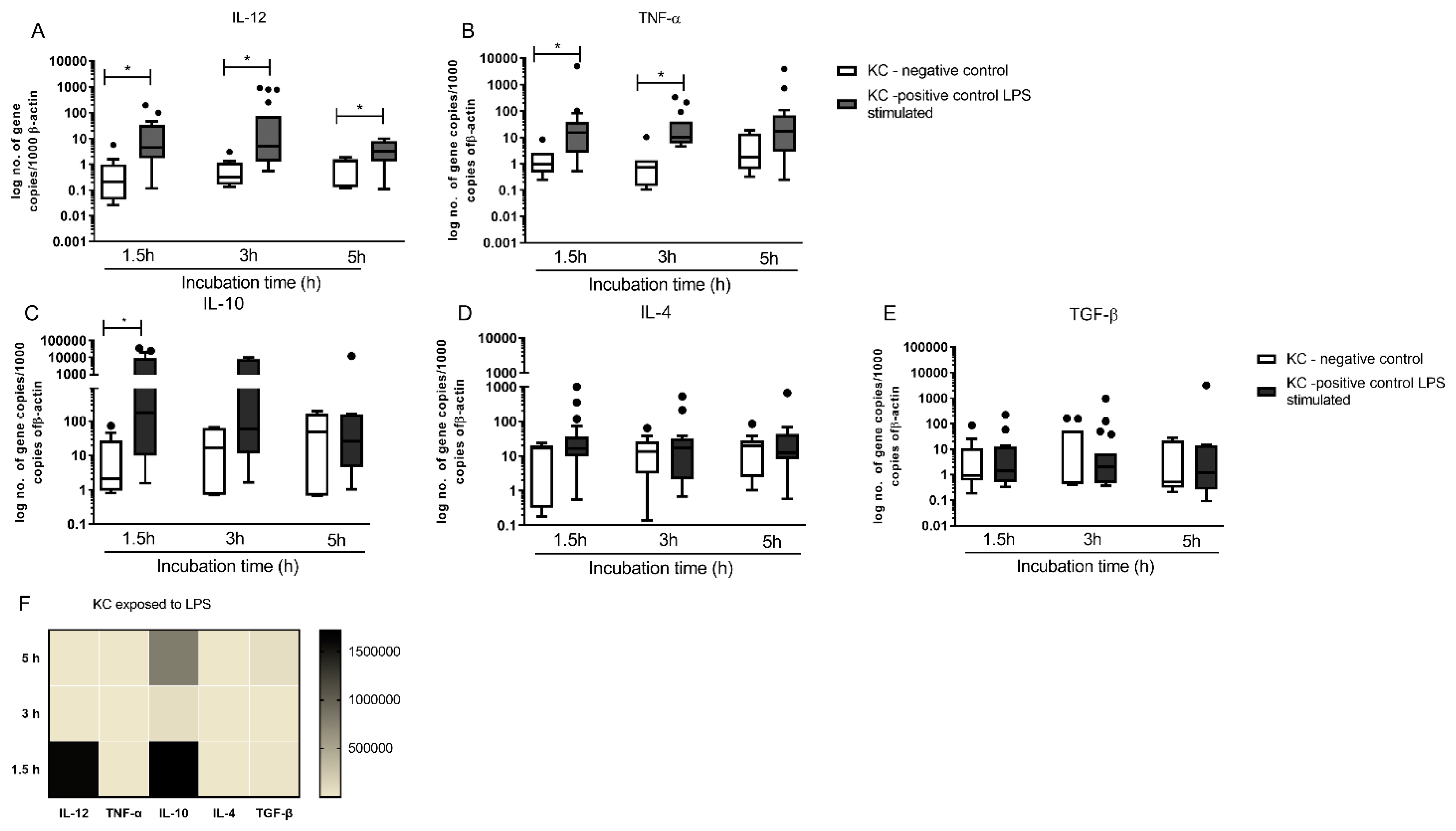
3.6. KCs Are Primed for Tolerance, but Can Generate High Levels of IL-12
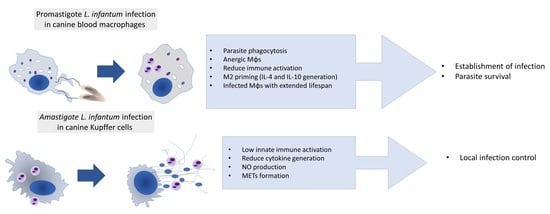
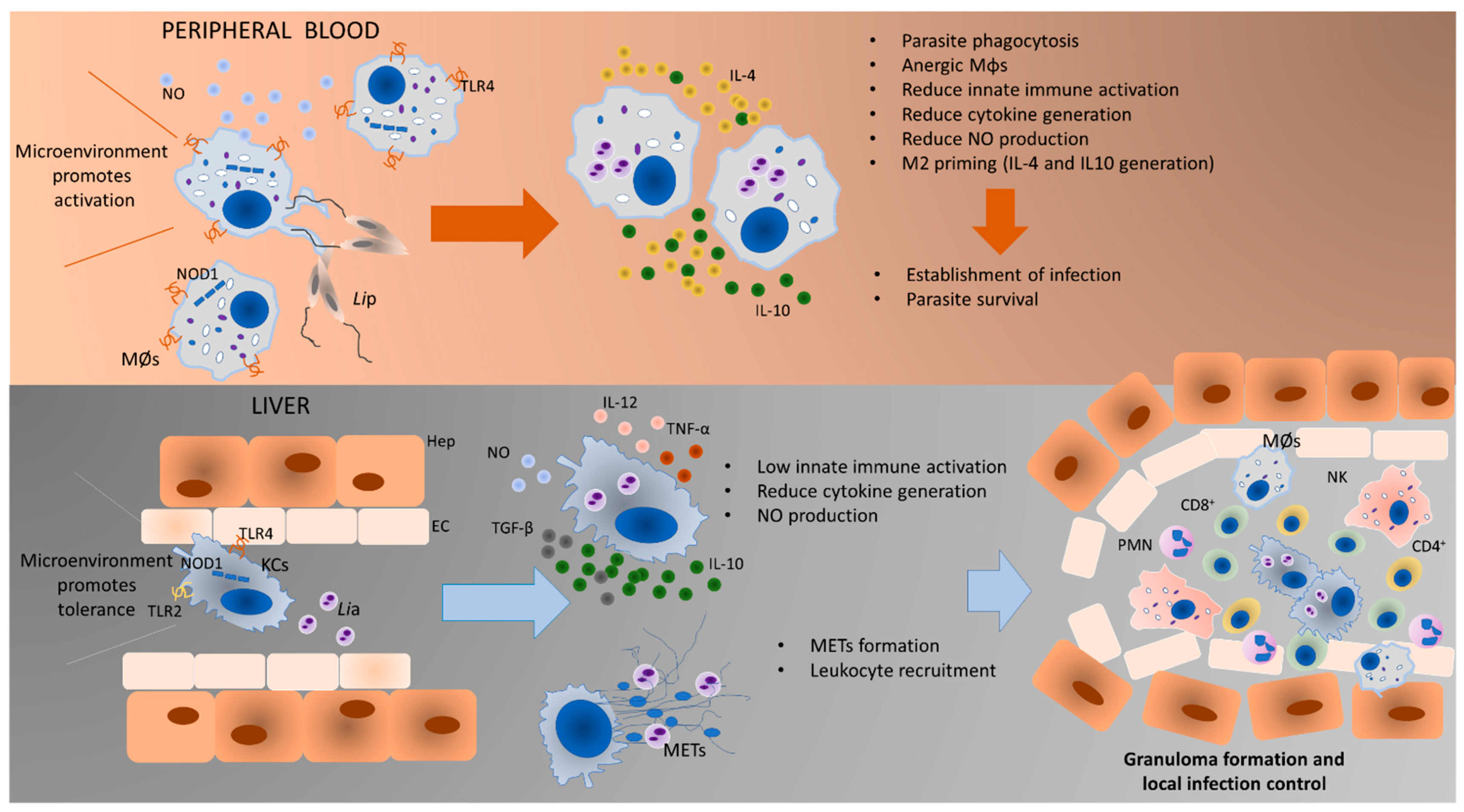
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Furth, R.; Beekhuizen, H. Monocytes. In Encyclopedia of Immunology; Elsevier: Amsterdam, The Netherlands, 1998; pp. 1750–1754. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Cao, X.; Fang, J.; Li, Y.; Fan, M. Macrophages polarization is mediated by the combination of PRR ligands and distinct inflammatory cytokines. Int. J. Clin. Exp. Pathol. 2015, 8, 10964–10974. [Google Scholar] [PubMed]
- Shaw, M.H.; Reimer, T.; Kim, Y.G.; Nuñez, G. NOD-like receptors (NLRs): Bona fide intracellular microbial sensors. Curr. Opin. Immunol. 2008, 20, 377–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yona, S.; Kim, K.W.; Wolf, Y.; Mildner, A.; Varol, D.; Breker, M.; Strauss-Ayali, D.; Viukov, S.; Guilliams, M.; Misharin, A.; et al. Fate Mapping Reveals Origins and Dynamics of Monocytes and Tissue Macrophages under Homeostasis. Immunity 2013, 38, 79–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginhoux, F.; Guilliams, M. Tissue-Resident Macrophage Ontogeny and Homeostasis. Immunity 2016, 44, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and Functions of Tissue Macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenne, C.N.; Kubes, P. Immune surveillance by the liver. Nat. Immunol. 2013, 14, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.J.; Barnes, M.; Tang, H.; Pritchard, M.T.; Nagy, L.E. Kupffer cells in the liver. Compr. Physiol. 2013, 3, 785–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomiotto-Pellissier, F.; Bortoleti, B.T.d.; Assolini, J.P.; Gonçalves, M.D.; Carloto, A.C.M.; Miranda-Sapla, M.M.; Conchon-Costa, I.; Bordignon, J.; Pavanelli, W.R. Macrophage Polarization in Leishmaniasis: Broadening Horizons. Front. Immunol. 2018, 9, 2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, M.A.; Alexandre-Pires, G.; Câmara, M.; Santos, M.; Martins, C.; Rodrigues, A.; Adriana, J.; Passero, L.F.D.; da Fonseca, I.P.; Santos-Gomes, G. Canine neutrophils cooperate with macrophages in the early stages of Leishmania infantum in vitro infection. Parasite Immunol. 2019, 41, e12617. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Gregory, D.J.; Forget, G. Subversion mechanisms by which Leishmania parasites can escape the host immune response: A signaling point of view. Clin. Microbiol. Rev. 2005, 18, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdan, C. Macrophages as host, effector and immunoregulatory cells in leishmaniasis: Impact of tissue micro-environment and metabolism. Cytokine X 2020, 2, 100041. [Google Scholar] [CrossRef]
- Rodrigues, A.; Santos-Mateus, D.; Alexandre-Pires, G.; Valério-Bolas, A.; Rafael-Fernandes, M.; Pereira, M.A.; Ligeiro, D.; de Jesus, J.; Alves-Azevedo, R.; Lopes-Ventura, S.; et al. Leishmania infantum exerts immunomodulation in canine Kupffer cells reverted by meglumine antimoniate. Comp. Immunol. Microbiol. Infect. Dis. 2017, 55, 42–52. [Google Scholar] [CrossRef]
- Santos-Gomes, G.M.; Abranches, P. Comparative study of infectivity caused by promastigotes of Leishmania infantum MON-1, L. infantum MON-24 and L. donovani MON-18. Folia Parasitol. 1996, 43, 7–12. [Google Scholar]
- Marques, C.S.; Passero, L.F.D.; Vale-Gato, I.; Rodrigues, A.; Rodrigues, O.R.; Martins, C.; Correia, I.; Tomás, A.M.; Alexandre-Pires, G.; Ferronha, M.H.; et al. New insights into neutrophil and Leishmania infantum in vitro immune interactions. Comp. Immunol. Microbiol. Infect. Dis. 2015, 40, 19–29. [Google Scholar] [CrossRef]
- Rodrigues, O.R.; Moura, R.A.; Gomes-Pereira, S.; Santos-Gomes, G.M. H-2 complex influences cytokine gene expression in Leishmania infantum-infected macrophages. Cell Immunol. 2006, 243, 118–126. [Google Scholar] [CrossRef]
- Glass, C.K.; Natoli, G. Molecular control of activation and priming in macrophages. Nat. Immunol. 2016, 17, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Gautiar, E.L.; Shay, T.; Miller, J.; Greter, M.; Jakubzick, C.; Ivanov, S.; Helft, J.; Chow, A.; Elpek, K.G.; Gordonov, S.; et al. Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat. Immunol. 2012, 13, 1118–1128. [Google Scholar] [CrossRef] [Green Version]
- Lavin, Y.; Winter, D.; Blecher-Gonen, R.; David, E.; Keren-Shaul, H.; Merad, M.; Jung, S.; Amit, I. Tissue-resident macrophage enhancer landscapes are shaped by the local microenvironment. Cell 2014, 159, 1312–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeksema, M.A.; De Winther, M.P.J. Epigenetic Regulation of Monocyte and Macrophage Function. Antioxid. Redox Signal. 2016, 25, 758–774. [Google Scholar] [CrossRef] [Green Version]
- Mowat, A.M.I.; Scott, C.L.; Bain, C.C. Barrier-tissue macrophages: Functional adaptation to environmental challenges. Nat. Med. 2017, 23, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uribe-Querol, E.; Rosales, C. Phagocytosis: Our Current Understanding of a Universal Biological Process. Front. Immunol. 2020, 11, 1066. [Google Scholar] [CrossRef] [PubMed]
- Mantegazza, A.R.; Magalhaes, J.G.; Amigorena, S.; Marks, M.S. Presentation of Phagocytosed Antigens by MHC Class I and II. Traffic 2013, 14, 135–152. [Google Scholar] [CrossRef] [Green Version]
- Podinovskaia, M.; Descoteaux, A. Leishmania and the macrophage: A multifaceted interaction. Future Microbiol. 2015, 10, 111–129. [Google Scholar] [CrossRef] [PubMed]
- De Menezes, J.P.; Saraiva, E.M.; Da Rocha-Azevedo, B. The site of the bite: Leishmania interaction with macrophages, neutrophils and the extracellular matrix in the dermis. Parasites Vectors 2016, 9, 264. [Google Scholar] [CrossRef] [Green Version]
- Rogers, M.; Kropf, P.; Choi, B.S.; Dillon, R.; Podinovskaia, M.; Bates, P.; Müller, I. Proteophosophoglycans regurgitated by Leishmania-infected sand flies target the L-arginine metabolism of host macrophages to promote parasite survival. PLoS Pathog. 2009, 5, e1000555. [Google Scholar] [CrossRef] [Green Version]
- Rogers, M.E.; Corware, K.; Müller, I.; Bates, P.A. Leishmania infantum proteophosphoglycans regurgitated by the bite of its natural sand fly vector, Lutzomyia longipalpis, promote parasite establishment in mouse skin and skin-distant tissues. Microb. Infect. 2010, 12, 875–879. [Google Scholar] [CrossRef]
- Poulaki, A.; Piperaki, E.T.; Voulgarelis, M. Effects of visceralising Leishmania on the spleen, liver, and bone marrow: A pathophysiological perspective. Microorganisms 2021, 9, 759. [Google Scholar] [CrossRef]
- Tiegs, G.; Lohse, A.W. Immune tolerance: What is unique about the liver. J. Autoimmun. 2010, 34, 1–6. [Google Scholar] [CrossRef]
- Rodrigues, A.; Alexandre-Pires, G.; Valério-Bolas, A.; Santos-Mateus, D.; Rafael-Fernandes, M.; Pereira, M.A.; Ligeiro, D.; Nunes, T.; Alves-Azevedo, R.; Lopes-Ventura, S.; et al. Dog hepatocytes are key effector cells in the liver innate immune response to Leishmania infantum. Parasitology 2019, 146, 753–764. [Google Scholar] [CrossRef]
- Figueiredo, M.M.; Deoti, B.; Amorim, I.F.; Pinto, A.J.W.; Moraes, A.; Carvalho, C.S.; da Silva, S.M.; de Assis, A.C.B.; de Faria, A.M.C.; Tafuri, W.L. Expression of regulatory T cells in jejunum, colon, and cervical and mesenteric lymph nodes of dogs naturally infected with Leishmania infantum. Infect. Immun. 2014, 82, 3704–3712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosein, S.; Blake, D.P.; Solano-Gallego, L. Insights on adaptive and innate immunity in canine leishmaniosis. Parasitology 2017, 144, 95–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchetti, A.P.; da Costa, L.F.; de Lima Romão, E.; Fujiwara, R.T.; da Paixão, T.A.; Santos, R.L. Transcription of innate immunity genes and cytokine secretion by canine macrophages resistant or susceptible to intracellular survival of Leishmania infantum. Vet. Immunol. Immunopathol. 2015, 163, 67–76. [Google Scholar] [CrossRef]
- Sanchez, M.A.; Diaz, N.L.; Zerpa, O.; Negron, E.; Convit, J.; Tapia, F.J. Organ-specific immunity in canine visceral leishmaniasis: Analysis of symptomatic and asymptomatic dogs naturally infected with Leishmania chagasi. Am. J. Trop. Med. Hyg. 2004, 70, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Tafuri, W.L.; Melo, F.A.; Moura, E.P.; Ribeiro, R.R.; Alves, C.F.; Caliari, M.V.; Tafuri, W.L.; Calabrese, K.d.; Tafuri, W.L. Hepatic extracellular matrix alterations in dogs naturally infected with Leishmania (Leishmania) chagasi. Int. J. Exp. Pathol. 2009, 90, 538. [Google Scholar] [CrossRef]
- Kaye, P.M.; Beattie, L. Lessons from other diseases: Granulomatous inflammation in leishmaniasis. Semin. Immunopathol. 2016, 38, 249. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.F.; Mosser, D.M. A novel phenotype for an activated macrophage: The type 2 activated macrophage. J. Leukoc. Biol. 2002, 72, 101–106. [Google Scholar] [CrossRef]
- Moreira, P.R.R.; Fernando, F.S.; Montassier, H.J.; André, M.R.; de Oliveira Vasconcelos, R. Polarized M2 macrophages in dogs with visceral leishmaniasis. Vet. Parasitol. 2016, 226, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doster, R.S.; Rogers, L.M.; Gaddy, J.A.; Aronoff, D.M. Macrophage Extracellular Traps: A Scoping Review. J. Innate Immun. 2018, 10, 3–13. [Google Scholar] [CrossRef]
- Brinkmann, V. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Guimarães-Costa, A.B.; Nascimento, M.T.C.; Froment, G.S.; Soares, R.P.P.; Morgado, F.N.; Conceição-Silva, F.; Saraiva, E.M. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc. Natl. Acad. Sci. USA 2009, 106, 6748–6753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valério-Bolas, A.; Pereira, M.; Alexandre-Pires, G.; Santos-Mateus, D.; Rodrigues, A.; Rafael-Fernandes, M.; Gabriela, A.; Passerod, F.; Santos-Gomesa, G. Intracellular and extracellular effector activity of mouse neutrophils in response to cutaneous and visceral Leishmania parasites. Cell Immunol. 2019, 335, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.M.A.; Carneiro, M.B.H.; Resende, M.E.; Martins, L.S.; dos Santos, L.M.; Vaz, L.G.; Mello, P.S.; Mosser, D.M.; Oliveira, M.A.P.; Vieira, L.Q. Neutrophils have a protective role during early stages of Leishmania amazonensis infection in BALB/c mice. Parasite Immunol. 2014, 36, 13–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, C.L.; Svensson, M.; Kaye, P.M. at the Single-Cell Level Macrophage Antigen Presentation Analyzed-Induced Inhibition of Leishmania. J. Immunol. Ref. 2003, 171, 6706–6713. [Google Scholar] [CrossRef] [Green Version]
- Stanley, A.C.; Engwerda, C.R. Balancing immunity and pathology in visceral leishmaniasis. Immunol. Cell Biol. 2007, 85, 138–147. [Google Scholar] [CrossRef]
- Rodrigues, A.V.; Alexandre-Pires, G.; Valério-Bolas, A.; Santos-Mateus, D.; Rafael-Fernandes, M.; Pereira, M.A.; Ligeiro, D.; Nunes, T.; Alves-Azevedo, R.; Santos, M.; et al. 3D-hepatocyte culture applied to parasitology: Immune activation of canine hepatic spheroids exposed to Leishmania infantum. Biomedicines 2020, 8, 628. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, A.V.; Valério-Bolas, A.; Alexandre-Pires, G.; Aires Pereira, M.; Nunes, T.; Ligeiro, D.; Pereira da Fonseca, I.; Santos-Gomes, G. Zoonotic Visceral Leishmaniasis: New Insights on Innate Immune Response by Blood Macrophages and Liver Kupffer Cells to Leishmania infantum Parasites. Biology 2022, 11, 100. https://doi.org/10.3390/biology11010100
Rodrigues AV, Valério-Bolas A, Alexandre-Pires G, Aires Pereira M, Nunes T, Ligeiro D, Pereira da Fonseca I, Santos-Gomes G. Zoonotic Visceral Leishmaniasis: New Insights on Innate Immune Response by Blood Macrophages and Liver Kupffer Cells to Leishmania infantum Parasites. Biology. 2022; 11(1):100. https://doi.org/10.3390/biology11010100
Chicago/Turabian StyleRodrigues, Armanda Viana, Ana Valério-Bolas, Graça Alexandre-Pires, Maria Aires Pereira, Telmo Nunes, Dário Ligeiro, Isabel Pereira da Fonseca, and Gabriela Santos-Gomes. 2022. "Zoonotic Visceral Leishmaniasis: New Insights on Innate Immune Response by Blood Macrophages and Liver Kupffer Cells to Leishmania infantum Parasites" Biology 11, no. 1: 100. https://doi.org/10.3390/biology11010100