
Interplay between Hypoxia and Extracellular Vesicles in Cancer and Inflammation

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Extracellular Vesicles
1.2. Diagnostic Value of EVs
1.3. Hypoxia
1.4. Purpose of the Review
2. Effects of Hypoxia on EVs in Inflammatory Diseases
3. Effects of Hypoxia on EVs in Cancer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [Green Version]
- De Toro, J.; Herschlik, L.; Waldner, C.; Mongini, C. Emerging Roles of Exosomes in Normal and Pathological Conditions: New Insights for Diagnosis and Therapeutic Applications. Front. Immunol. 2015, 6, 203. [Google Scholar] [CrossRef] [Green Version]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Gould, S.J.; Raposo, G. As We Wait: Coping with an Imperfect Nomenclature for Extracellular Vesicles. J. Extracell. Vesicles 2013, 2, 20389. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular Vesicles in Cancer: Exosomes, Microvesicles and the Emerging Role of Large Oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of Distinct Nanoparticles and Subsets of Extracellular Vesicles by Asymmetric Flow Field-Flow Fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef]
- Muralidharan-Chari, V.; Clancy, J.W.; Sedgwick, A.; D’Souza-Schorey, C. Microvesicles: Mediators of Extra-cellular Communication during Cancer Progression. J. Cell Sci. 2010, 123, 1603. [Google Scholar] [CrossRef] [Green Version]
- Schorey, J.S.; Bhatnagar, S. Exosome Function: From Tumor Immunology to Pathogen Biology. Traffic 2008, 9, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver MiR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res. 2016, 76, 1770. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.J.; Kalra, H.; Mathivanan, S. ExoCarta as a Resource for Exosomal Research. J. Extracell. Vesicle 2012, 1, 18374. [Google Scholar] [CrossRef]
- Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; Bertier, L.; et al. EV-TRACK: Transparent Reporting and Centralizing Knowledge in Extracellular Vesicle Research. Nat. Methods 2017, 14, 228–232. [Google Scholar] [CrossRef]
- Jia, S.; Zocco, D.; Samuels, M.L.; Chou, M.F.; Chammas, R.; Skog, J.; Zarovni, N.; Momen-Heravi, F.; Kuo, W.P. Emerging Technologies in Extracellular Vesicle-Based Molecular Diagnostics. Expert Rev. Mol. Diagn. 2014, 14, 307–321. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Baixauli, F.; Gutiérrez-Vázquez, C.; Sánchez-Madrid, F.; Mittelbrunn, M. Sorting It out: Regulation of Exosome Loading. Semin. Cancer Biol. 2014, 28, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An Integrated Database of High-Throughput Data for Systemic Analyses of Extracellular Vesicles. J. Extracell. Vesicles 2013, 2, 20384. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinado, H.; Lavotshkin, S.; Lyden, D. The Secreted Factors Responsible for Pre-Metastatic Niche Formation: Old Sayings and New Thoughts. Semin. Cancer Biol. 2011, 21, 139–146. [Google Scholar] [CrossRef]
- Van Niel, G.; Porto-Carreiro, I.; Simoes, S.; Raposo, G. Exosomes: A Common Pathway for a Specialized Func-tion. J. Biochem. 2006, 140, 13–21. [Google Scholar] [CrossRef]
- Milane, L.; Singh, A.; Mattheolabakis, G.; Suresh, M.; Amiji, M.M. Exosome Mediated Communication within the Tumor Microenvironment. J. Control. Release 2015, 219, 278–294. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of Secretion and Uptake of Exosomes and Other Extracellular Vesicles for Cell-to-Cell Communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Wang, T.; Gilkes, D.M.; Takano, N.; Xiang, L.; Luo, W.; Bishop, C.J.; Chaturvedi, P.; Green, J.J.; Semenza, G.L. Hypoxia-Inducible Factors and RAB22A Mediate Formation of Microvesicles That Stimulate Breast Cancer Invasion and Metastasis. Proc. Natl. Acad. Sci. USA 2014, 111, E3234. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, C.; Kalluri, R. Exosomes in Tumor Microenvironment Influence Cancer Progression and Metastasis. J. Mol. Med. 2013, 91, 431–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quail, D.F.; Joyce, J.A. Microenvironmental Regulation of Tumor Progression and Metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Zocco, D.; Ferruzzi, P.; Cappello, F.; Kuo, W.P.; Fais, S. Extracellular Vesicles as Shuttles of Tumor Biomarkers and Anti-Tumor Drugs. Front. Oncol. 2014, 4, 267. [Google Scholar] [CrossRef] [Green Version]
- Park, J.E.; Tan, H.S.; Datta, A.; Lai, R.C.; Zhang, H.; Meng, W.; Lim, S.K.; Sze, S.K. Hypoxic Tumor Cell Modu-lates Its Microenvironment to Enhance Angiogenic and Metastatic Potential by Secretion of Proteins and Ex-osomes *. Mol. Cell. Proteom. 2010, 9, 1085–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-B.; Zhang, Z.-R.; Schluesener, H.J.; Xu, S.-Q. Role of Exosomes in Immune Regulation. J. Cell. Mol. Med. 2006, 10, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current Knowledge of Their Composition, Biological Functions, and Diagnostic and Therapeutic Potentials. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, C.; Zhou, T.; Liu, X.; Liu, X.; Li, X.; Chen, D. Role of Exosomal Proteins in Cancer Diagnosis. Mol. Cancer 2017, 16, 145. [Google Scholar] [CrossRef] [PubMed]
- Torrano, V.; Royo, F.; Peinado, H.; Loizaga-Iriarte, A.; Unda, M.; Falcón-Perez, J.M.; Carracedo, A. Vesi-cle-MaNiA: Extracellular Vesicles in Liquid Biopsy and Cancer. Curr. Opin. Pharmacol. 2016, 29, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturella, M.; Carpi, F.M.; Zocco, D. Standardization of Blood Collection and Processing for the Diagnostic Use of Extracellular Vesicles. Curr. Pathobiol. Rep. 2019, 7, 1–8. [Google Scholar] [CrossRef]
- Pant, S.; Hilton, H.; Burczynski, M.E. The Multifaceted Exosome: Biogenesis, Role in Normal and Aberrant Cellular Function, and Frontiers for Pharmacological and Biomarker Opportunities. Biochem. Pharmacol. 2012, 83, 1484–1494. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, B.N.; Ding, W.-Q. Intercellular Communication by Exosome-Derived MicroRNAs in Cancer. Int. J. Mol. Sci. 2013, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, J.; D’Souza-Schorey, C. Extracellular Vesicles in Cancer: Purpose and Promise. Cancer J. 2018, 24, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal MiRNAs as Biomarkers for Prostate Cancer. Front. Genet. 2013, 4, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaka, N.; Kogure, A.; Yamamoto, T.; Urabe, F.; Usuba, W.; Prieto-Vila, M.; Ochiya, T. Exploiting the Message from Cancer: The Diagnostic Value of Extracellular Vesicles for Clinical Applications. Exp. Mol. Med. 2019, 51, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, T.; Wong, D.T.W. Liquid Biopsy in Head and Neck Cancer: Promises and Challenges. J. Dent. Res. 2018, 97, 701–708. [Google Scholar] [CrossRef]
- Taylor, C.T.; Colgan, S.P. Regulation of Immunity and Inflammation by Hypoxia in Immunological Niches. Nat. Rev. Immunol. 2017, 17, 774–785. [Google Scholar] [CrossRef]
- Vanderkooi, J.M.; Erecinska, M.; Silver, I.A. Oxygen in Mammalian Tissue: Methods of Measurement and Af-finities of Various Reactions. Am. J. Physiol. Cell Physiol. 1991, 260, C1131–C1150. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and Inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.P.; Keogh, C.E.; Crean, D.; Taylor, C.T. The Role of HIF in Immunity and Inflammation. Mol. Asp. Med. 2016, 47–48, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Monaci, S.; Aldinucci, C.; Rossi, D.; Giuntini, G.; Filippi, I.; Ulivieri, C.; Marotta, G.; Sozzani, S.; Carraro, F.; Naldini, A. Hypoxia Shapes Autophagy in LPS-Activated Dendritic Cells. Front. Immunol. 2020, 11, 3071. [Google Scholar] [CrossRef] [PubMed]
- Naldini, A.; Morena, E.; Pucci, A.; Pellegrini, M.; Baldari, C.T.; Pelicci, P.G.; Presta, M.; Ribatti, D.; Carraro, F. The Adaptor Protein P66shc Is a Positive Regulator in the Angiogenic Response Induced by Hypoxic T Cells. J. Leukoc. Biol. 2010, 87, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Bernards, R.; Weinberg, R.A. Metastasis Genes: A Progression Puzzle. Nature 2002, 418, 823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, A.L. Hypoxia—A Key Regulatory Factor in Tumour Growth. Nat. Rev. Cancer 2002, 2, 38–47. [Google Scholar] [CrossRef]
- Vaupel, P.; Mayer, A. Hypoxia in Cancer: Significance and Impact on Clinical Outcome. Cancer Metastasis Rev. 2007, 26, 225–239. [Google Scholar] [CrossRef]
- Meehan, J.; Ward, C.; Turnbull, A.; Bukowski-Wills, J.; Finch, A.J.; Jarman, E.J.; Xintaropoulou, C.; Mar-tinez-Perez, C.; Gray, M.; Pearson, M.; et al. Inhibition of PH Regulation as a Therapeutic Strategy in Hypoxic Human Breast Cancer Cells. Oncotarget 2017, 8, 42857–42875. [Google Scholar] [CrossRef]
- Finger, E.C.; Giaccia, A.J. Hypoxia, Inflammation, and the Tumor Microenvironment in Metastatic Disease. Cancer Metastasis Rev. 2010, 29, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Yang, F.; Miao, S.; Liu, W.; Wang, C.; Shu, Y.; Shen, H. Role of Hypoxia-Induced Exosomes in Tumor Biology. Mol. Cancer 2018, 17, 120. [Google Scholar] [CrossRef]
- Deep, G.; Panigrahi, G.K. Hypoxia-Induced Signaling Promotes Prostate Cancer Progression: Exosomes Role as Messenger of Hypoxic Response in Tumor Microenvironment. Crit. Rev. Oncog. 2015, 20, 419–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Shi, R.; Zhang, Q. Hypoxia and Oxygen-Sensing Signaling in Gene Regulation and Cancer Progres-sion. Int. J. Mol. Sci. 2020, 21, 8162. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.W.; Shakir, D.; Batie, M.; Frost, M.; Rocha, S. Oxygen-Sensing Mechanisms in Cells. FEBS J. 2020, 287, 3888–3906. [Google Scholar] [CrossRef] [PubMed]
- Ortmann, B.; Druker, J.; Rocha, S. Cell Cycle Progression in Response to Oxygen Levels. Cell. Mol. Life Sci. 2014, 71, 3569–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Kang, Y. Hypoxia and Hypoxia-Inducible Factors: Master Regulators of Metastasis. Clin. Cancer Res. 2010, 16, 5928. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-Inducible Factors in Physiology and Medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Aga, M.; Bentz, G.L.; Raffa, S.; Torrisi, M.R.; Kondo, S.; Wakisaka, N.; Yoshizaki, T.; Pagano, J.S.; Shackelford, J. Exosomal HIF1α Supports Invasive Potential of Nasopharyngeal Carcinoma-Associated LMP1-Positive Exo-somes. Oncogene 2014, 33, 4613–4622. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Life with Oxygen. Science 2007, 318, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen Sensing by Metazoans: The Central Role of the HIF Hydroxylase Path-way. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef]
- Smith, T.G.; Robbins, P.A.; Ratcliffe, P.J. The Human Side of Hypoxia-Inducible Factor. Br. J. Haematol. 2008, 141, 325–334. [Google Scholar] [CrossRef]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yang, Y.; Huang, Y.; Zhang, L.; Ling, Z.; Zhu, Y.; Wang, F.; Zou, X.; Chen, M. Hypoxia-Induced Ex-tracellular Vesicles Mediate Protection of Remote Ischemic Preconditioning for Renal Ischemia-Reperfusion Injury. Biomed. Pharmacother. 2017, 90, 473–478. [Google Scholar] [CrossRef] [PubMed]
- King, H.W.; Michael, M.Z.; Gleadle, J.M. Hypoxic Enhancement of Exosome Release by Breast Cancer Cells. BMC Cancer 2012, 12, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dengler, V.L.; Galbraith, M.; Espinosa, J.M. Transcriptional Regulation by Hypoxia Inducible Factors. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Chen, M.; Jiang, R.; Guo, Y.; Wu, M.; Zhang, X. Exosome-Related Tumor Microenvironment. J. Cancer 2018, 9, 3084–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.E.; Dutta, B.; Tse, S.W.; Gupta, N.; Tan, C.F.; Low, J.K.; Yeoh, K.W.; Kon, O.L.; Tam, J.P.; Sze, S.K. Hypox-ia-Induced Tumor Exosomes Promote M2-like Macrophage Polarization of Infiltrating Myeloid Cells and Mi-croRNA-Mediated Metabolic Shift. Oncogene 2019, 38, 5158–5173. [Google Scholar] [CrossRef]
- Belting, M.; Christianson, H.C. Role of Exosomes and Microvesicles in Hypoxia-Associated Tumour Develop-ment and Cardiovascular Disease. J. Intern. Med. 2015, 278, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Yaghoubi, S.; Najminejad, H.; Dabaghian, M.; Karimi, M.H.; Abdollahpour-Alitappeh, M.; Rad, F.; Mahi-Birjand, M.; Mohammadi, S.; Mohseni, F.; Sobhani Lari, M.; et al. How Hypoxia Regulate Exosomes in Ischemic Diseases and Cancer Microenvironment? IUBMB Life 2020, 72, 1286–1305. [Google Scholar] [CrossRef]
- Ramteke, A.; Ting, H.; Agarwal, C.; Mateen, S.; Somasagara, R.; Hussain, A.; Graner, M.; Frederick, B.; Agarwal, R.; Deep, G. Exosomes Secreted under Hypoxia Enhance Invasiveness and Stemness of Prostate Cancer Cells by Targeting Adherens Junction Molecules. Mol. Carcinog. 2015, 54, 554–565. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Luo, H.; Li, X.; Li, T.; He, J.; Qi, Q.; Liu, Y.; Yu, Z. Exosomes Derived from Human Pulmonary Artery Endothelial Cells Shift the Balance between Proliferation and Apoptosis of Smooth Muscle Cells. Cardiology 2017, 137, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.T.; Melo, S.; Özdemir, B.C.; Kato, N.; Revuelta, I.; Miller, C.A.; Ii, V.H.G.; LeBleu, V.S.; Kalluri, R. TGF-β1–Containing Exosomes from Injured Epithelial Cells Activate Fibroblasts to Initiate Tissue Regenerative Responses and Fibrosis. J. Am. Soc. Nephrol. 2012, 24, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Sano, S.; Izumi, Y.; Yamaguchi, T.; Yamazaki, T.; Tanaka, M.; Shiota, M.; Osada-Oka, M.; Nakamura, Y.; Wei, M.; Wanibuchi, H.; et al. Lipid Synthesis Is Promoted by Hypoxic Adipocyte-Derived Exosomes in 3T3-L1 Cells. Biochem. Biophys. Res. Commun. 2014, 445, 327–333. [Google Scholar] [CrossRef]
- Khalyfa, A.; Zhang, C.; Khalyfa, A.A.; Foster, G.E.; Beaudin, A.E.; Andrade, J.; Hanly, P.J.; Poulin, M.J.; Gozal, D. Effect on Intermittent Hypoxia on Plasma Exosomal Micro RNA Signature and Endothelial Function in Healthy Adults. Sleep 2016, 39, 2077–2090. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhou, X.; Yao, Q.; Liu, Y.; Zhang, H.; Dong, Z. HIF-1-Mediated Production of Exosomes during Hy-poxia Is Protective in Renal Tubular Cells. Am. J. Physiol. Ren. Physiol. 2017, 313, F906–F913. [Google Scholar] [CrossRef] [PubMed]
- Khalyfa, A.; Almendros, I.; Gileles-Hillel, A.; Akbarpour, M.; Trzepizur, W.; Mokhlesi, B.; Huang, L.; Andrade, J.; Farré, R.; Gozal, D. Circulating Exosomes Potentiate Tumor Malignant Properties in a Mouse Model of Chronic Sleep Fragmentation. Oncotarget 2016, 7, 54676–54690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almendros, I.; Khalyfa, A.; Trzepizur, W.; Gileles-Hillel, A.; Huang, L.; Akbarpour, M.; Andrade, J.; Farré, R.; Gozal, D. Tumor Cell Malignant Properties Are Enhanced by Circulating Exosomes in Sleep Apnea. CHEST 2016, 150, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, H.; Xu, K.; Ling, Z.; Huang, Y.; Hu, Q.; Lu, K.; Liu, C.; Wang, Y.; Liu, N.; et al. Hypoxia Precondi-tioned Renal Tubular Epithelial Cell-Derived Extracellular Vesicles Alleviate Renal Ischaemia-Reperfusion Injury Mediated by the HIF-1α/Rab22 Pathway and Potentially Affected by MicroRNAs. Int. J. Biol. Sci. 2019, 15, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.D.; French, K.M.; Ghosh-Choudhary, S.; Maxwell, J.T.; Brown, M.E.; Platt, M.O.; Searles, C.D.; Davis, M.E. Identification of Therapeutic Covariant MicroRNA Clusters in Hypoxia-Treated Cardiac Progenitor Cell Exosomes Using Systems Biology. Circ. Res. 2015, 116, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.-P.; Tian, T.; Wang, J.-Y.; He, J.-N.; Chen, T.; Pan, M.; Xu, L.; Zhang, H.; Qiu, X.-T.; Li, C.-C.; et al. Hypox-ia-Elicited Mesenchymal Stem Cell-Derived Exosomes Facilitates Cardiac Repair through MiR-125b-Mediated Prevention of Cell Death in Myocardial Infarction. Theranostics 2018, 8, 6163–6177. [Google Scholar] [CrossRef]
- Gonzalez-King, H.; García, N.A.; Ontoria-Oviedo, I.; Ciria, M.; Montero, J.A.; Sepúlveda, P. Hypoxia Inducible Factor-1α Potentiates Jagged 1-Mediated Angiogenesis by Mesenchymal Stem Cell-Derived Exosomes. Stem Cells 2017, 35, 1747–1759. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Deng, L.; Wang, D.; Li, N.; Chen, X.; Cheng, X.; Yuan, J.; Gao, X.; Liao, M.; Wang, M.; et al. Mechanism of TNF-α Autocrine Effects in Hypoxic Cardiomyocytes: Initiated by Hypoxia Inducible Factor 1α, Presented by Exosomes. J. Mol. Cellular Cardiol. 2012, 53, 848–857. [Google Scholar] [CrossRef]
- Duan, P.; Tan, J.; Miao, Y.; Zhang, Q. Potential Role of Exosomes in the Pathophysiology, Diagnosis, and Treatment of Hypoxic Diseases. Am. J. Transl. Res. 2019, 11, 1184–1201. [Google Scholar] [PubMed]
- Wysoczynski, M.; Ratajczak, M.Z. Lung Cancer Secreted Microvesicles: Underappreciated Modulators of Mi-croenvironment in Expanding Tumors. Int. J. Cancer 2009, 125, 1595–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadokoro, H.; Umezu, T.; Ohyashiki, K.; Hirano, T.; Ohyashiki, J.H. Exosomes Derived from Hypoxic Leukemia Cells Enhance Tube Formation in Endothelial Cells *. J. Biol. Chem. 2013, 288, 34343–34351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezu, T.; Tadokoro, H.; Azuma, K.; Yoshizawa, S.; Ohyashiki, K.; Ohyashiki, J.H. Exosomal MiR-135b Shed from Hypoxic Multiple Myeloma Cells Enhances Angiogenesis by Targeting Factor-Inhibiting HIF-1. Blood 2014, 124, 3748–3757. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-L.; Hung, J.-Y.; Chang, W.-A.; Lin, Y.-S.; Pan, Y.-C.; Tsai, P.-H.; Wu, C.-Y.; Kuo, P.-L. Hypoxic Lung Cancer-Secreted Exosomal MiR-23a Increased Angiogenesis and Vascular Permeability by Targeting Prolyl Hydroxylase and Tight Junction Protein ZO-1. Oncogene 2017, 36, 4929–4942. [Google Scholar] [CrossRef] [PubMed]
- Dorayappan, K.D.P.; Wanner, R.; Wallbillich, J.J.; Saini, U.; Zingarelli, R.; Suarez, A.A.; Cohn, D.E.; Selvendiran, K. Hypoxia-Induced Exosomes Contribute to a More Aggressive and Chemoresistant Ovarian Cancer Pheno-type: A Novel Mechanism Linking STAT3/Rab Proteins. Oncogene 2018, 37, 3806–3821. [Google Scholar] [CrossRef]
- Yu, Y.; Min, Z.; Zhou, Z.; Linhong, M.; Tao, R.; Yan, L.; Song, H. Hypoxia-Induced Exosomes Promote Hepatocellular Carcinoma Proliferation and Metastasis via MiR-1273f Transfer. Experimental Cell Res. 2019, 385, 111649. [Google Scholar] [CrossRef] [PubMed]
- Patton, M.C.; Zubair, H.; Khan, M.A.; Singh, S.; Singh, A.P. Hypoxia Alters the Release and Size Distribution of Extracellular Vesicles in Pancreatic Cancer Cells to Support Their Adaptive Survival. J. Cell Biochem. 2020, 121, 828–839. [Google Scholar] [CrossRef]
- Jung, K.O.; Youn, H.; Lee, C.-H.; Kang, K.W.; Chung, J.-K. Visualization of Exosome-Mediated MiR-210 Trans-fer from Hypoxic Tumor Cells. Oncotarget 2016, 8, 9899–9910. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xu, R.; Xia, J.; Huang, J.; Su, B.; Wang, S. Aspirin Inhibits Hypoxia-Mediated Lung Cancer Cell Stem-ness and Exosome Function. Pathol. Res. Pract. 2019, 215, 152379. [Google Scholar] [CrossRef] [PubMed]
- Svensson, K.J.; Kucharzewska, P.; Christianson, H.C.; Sköld, S.; Löfstedt, T.; Johansson, M.C.; Mörgelin, M.; Bengzon, J.; Ruf, W.; Belting, M. Hypoxia Triggers a Proangiogenic Pathway Involving Cancer Cell Microvesi-cles and PAR-2–Mediated Heparin-Binding EGF Signaling in Endothelial Cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharzewska, P.; Christianson, H.C.; Welch, J.E.; Svensson, K.J.; Fredlund, E.; Ringnér, M.; Mörgelin, M.; Bourseau-Guilmain, E.; Bengzon, J.; Belting, M. Exosomes Reflect the Hypoxic Status of Glioma Cells and Me-diate Hypoxia-Dependent Activation of Vascular Cells during Tumor Development. Proc. Natl. Acad. Sci. USA 2013, 110, 7312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaepfer, I.R.; Nambiar, D.K.; Ramteke, A.; Kumar, R.; Dhar, D.; Agarwal, C.; Bergman, B.; Graner, M.; Maro-ni, P.; Singh, R.P.; et al. Hypoxia Induces Triglycerides Accumulation in Prostate Cancer Cells and Extracellu-lar Vesicles Supporting Growth and Invasiveness Following Reoxygenation. Oncotarget 2015, 6, 22836–22856. [Google Scholar] [CrossRef]
- Wang, Y.; Yi, J.; Chen, X.; Zhang, Y.; Xu, M.; Yang, Z. The Regulation of Cancer Cell Migration by Lung Cancer Cell-Derived Exosomes through TGF-β and IL-10. Oncol. Lett. 2016, 11, 1527–1530. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Luo, G.; Zhang, K.; Cao, J.; Huang, C.; Jiang, T.; Liu, B.; Su, L.; Qiu, Z. Correction: Hypoxic Tu-mor-Derived Exosomal MiR-301a Mediates M2 Macrophage Polarization via PTEN/PI3Kγ to Promote Pan-creatic Cancer Metastasis. Cancer Res. 2020, 80, 922. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Feng, Y. Exosomes Derived From Hypoxic Colorectal Cancer Cells Promote Angiogenesis Through Wnt4-Induced β-Catenin Signaling in Endothelial Cells. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2017, 25, 651–661. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Diagnostic Values of EVs |
|---|
| Multiple biomarker candidates (proteins, nucleic acids) |
| Biomarkers protected from degradation |
| Different sources (almost all body fluids) |
| Plasma and serum easily accessible, abundant and biobanked |
| Sample representative of the patient status |
| Minimally invasive sampling |
| Type of Inflammatory Disease | Source of EVs | Functions of Released EVs | Biomolecules Carried by Hypoxic EVs | References |
|---|---|---|---|---|
| Pulmonary arterial hypertension (PAH) | Human pulmonary artery endothelial cells (PAECs) | Pulmonary vascular remodeling | miR-17, miR-20a, miR-21, miR145 (to be investigated) | [70] |
| Kidney fibrosis | Mouse and human tubular epithelial cells (TECs) | Repair in injured parenchyma; fibroblasts’ activation and proliferation | TGF-β1 mRNA | [71] |
| Obesity | Mouse 3T3-L1 adipocytes | Delivery of proteins involved in metabolic processes; accumulation of lipids in normoxic cells | G6PD, FASN, ACC enzymes | [72] |
| AKI (Acute Kidney Injury) | Rat serum; human kidney cells HK-2 | Renoprotective effects | not described | [62] |
| Human renal tubular epithelial cells (RTEC) | Therapeutic effects in renal ischaemia-reperfusion (I/R) injury | miR-129-5p, miR-138-5p, miR-127-3p, miR-9-5p, miR-125b-5p, miR-129a-2-3p, miR-124-3p, miR-136-3p, miR-135a-5p, miR-411-5p, miR-129-2-3p, miR-9-3p, miR-330-5p, miR-128-3p, miR-218-5p, miR-148a-3p | [77] | |
| Rat renal proximal tubular cells (RPTC) | Cytoprotective role | to be investigated | [74] | |
| Obstructive sleep apnea (OSA) | Human plasma (sources of exosomes: endothelial cells, progenitor cells, monocytes, lymphocytes, and platelets) | Cardiovascular dysfunction; regulation of immune and atherosclerosis pathways | hsa-miR-4649-3p, hsa-miR-4436b-5p, hsa-miR-483-3p, hsa-miR-1202, hsa-miR-4505 | [73] |
| Plasma of mice and patients | Regulation of cancer pathways | mmu-miR-5128, mmu-miR-5112, mmu-miR-6366 | [75] | |
| Plasma of mice and patients | In vitro proliferation and migration of cells; metastatic behavior with disruption of the endothelial monolayer barrier; regulation of cancer pathways and inflammation | mmu-miR-671-5p, mmu-miR-6418-5p, mmu-miR-709, mmu-miR-6366, mmu-miR-5100, mmu-miR-2137, mmu-miR-882, mmu-miR-92a-3p, mmu-miR-451a, mmu-miR-3082-5p, mmu-miR-5113 | [76] | |
| Myocardial infarction (MI) and acute myocardial infarction (AMI) | Cardiac progenitor cells (CPCs) | Therapeutic potential | miR-17, miR-199a, miR-210, miR-292, miR-103, miR-15b, miR-20a | [78] |
| Cardiomyocytes | Regulation of inflammatory responses | TNF-α | [81] | |
| Murine MSCs | Ischemic myocardium repair | miR-125b-5p | [79] | |
| Ischemic-tissue related diseases | MSCs | Angiogenesis, therapeutic potential | Jagged1 protein, miR15, miR16, miR-17, miR31, miR126, miR145, miR221, miR222, miR320a, miR424 | [80] |
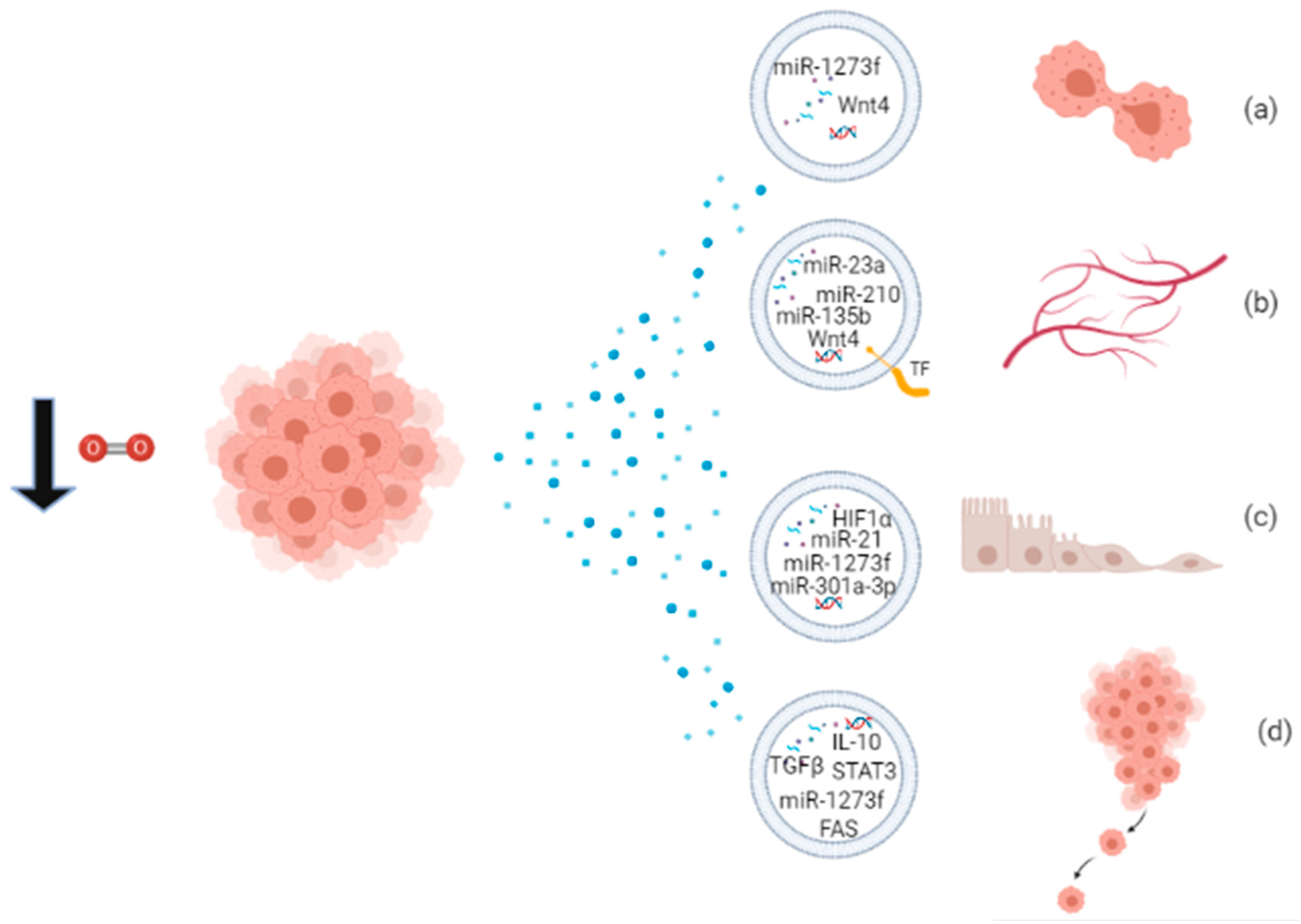
| Cancer Disease | Source of EVs | Functions of Released EVs | Biomolecules Carried by Hypoxic EVs | References |
|---|---|---|---|---|
| Prostate cancer (PCA) | LNCaP and PC3 human PCA cells | Invasiveness, motility and stemness of naïve PCA cells; metalloproteinase activity; remodeling of epithelial adherens junction pathways; induction of CAF-type phenotype in prostate fibroblasts; PCA growth and invasiveness | Proteins: MMP-9, MMP-2, TGF-β2, TNF1α, IL6, TSG101, Akt, ILK1, β-catenin; triglycerides | [69,94] |
| Epidermoid carcinoma | A431 human squamous carcinoma cells | Angiogenesis; metastasis | not described | [26] |
| Glioblastoma multiforme (GBM) | U87-MG human GBM cells; plasma of tumor-bearing mice; plasma of GBM patients | Pro-angiogenic effects | mRNAs: ADM, LOX, IGFPB, BCL, BNIP3, NDRG1, PLOD2, PAI1; proteins: IL8, IGFBP1, IGFBP3; MMP9, PTX3, PDGF-AB/AA, CD26, PAI1, CAV1 | [93] |
| Breast cancer | U87-MG human GBM cells | Tumor development; angiogenesis | TF | [92] |
| MCF-7, MDA-MB-231, MDA-MB-435 human breast cancer cells | Endothelial cell tubulogenesis; mechanisms of repressing DNA repair | miR-210 | [63] | |
| Focal adhesion formation, invasion and metastasis | not described | [22] | ||
| 4T1 mouse breast cancer cells | Angiogenesis | miR-210 | [90] | |
| Leukemia | K562 human leukemia cells | HUVECs tube formation | miR-210 | [84] |
| Multiple myeloma | RPMI8226, KMS-11, U266 human multiple myeloma cells | Tube formation; angiogenesis; regulation of FIH molecular pathway | miR-210, miR135b | [85] |
| Lung cancer | CL1-5 human lung cancer cells | Local and distant angiogenesis; increased vascular permeability; cancer transendothelial migration | miR-23a | [86] |
| NCI-H1688 human small cell lung cancer and NCI-H2228 human non-small cell lung cancer | Migration of endothelial and cancer cells; metastasis | TGF-β and IL-10 | [95] | |
| A549 human lung adenocarcinoma cells | Angiogenesis, metastasis, cancer cell survival, migration and tube formation | miR-135b, miR-210 | [91] | |
| Ovarian cancer | OVCAR-8, A2780, TR127, TR182 human ovarian cancer cells; patient-derived ascites | Tumor progression, metastasis and chemoresistance | STAT3, FAS proteins | [87] |
| Oral squamous cell carcinoma (OSCC) | SCC-9, CAL-27 human OSCC cells; serum of OSCC patients | EMT, migration and invasion of target normoxic cells | miR-21; miR-205, miR-148b (to be investigated) | [11] |
| Nasopharyngeal carcinoma (NPC) | NP69 and AdAH human NPC cells, transfected with latent membrane protein 1 (LMP1) | EMT; cancer progression and invasive potential | HIF1α | [57] |
| Colorectal cancer (CRC) | HT29 and HCT116 human CRC cells | Endothelial cells proliferation and migration; tumor growth and angiogenesis | Wnt4 protein | [97] |
| Melanoma | B16-F0 mouse melanoma cells, A375 human melanoma cells, A431 squamous skin carcinoma cells, A549 lung adenocarcinoma cells | Monocyte/macrophage recruitment in vitro and in vivo and host immunosuppression; tumor cell proliferation | chemokines and growth factors (CSF-1, CCL2, EMAP2, TGFβ, FTH, FTL); miR-let-7a, miR-21a | [66] |
| Hepatocellular carcinoma (HCC) | Huh7 and MHCC-97H human HCC cells | Proliferation, migration, invasiveness, EMT in normoxic HCC cells | miR-1273f, miR-93-5p, miR-221-3p | [88] |
| Pancreatic cancer (PC) | MiaPaCa and AsPC1 PC cell lines | Adaptive survival of PC hypoxic cancer cells | not described | |
| PANC-1, BxPC-3 cell lines; serum of PC patients | M2 polarization of macrophages, metastatic behavior of PC cells in vitro and in vivo | miR-301a-3p | [96] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venturella, M.; Criscuoli, M.; Carraro, F.; Naldini, A.; Zocco, D. Interplay between Hypoxia and Extracellular Vesicles in Cancer and Inflammation. Biology 2021, 10, 606. https://doi.org/10.3390/biology10070606
Venturella M, Criscuoli M, Carraro F, Naldini A, Zocco D. Interplay between Hypoxia and Extracellular Vesicles in Cancer and Inflammation. Biology. 2021; 10(7):606. https://doi.org/10.3390/biology10070606
Chicago/Turabian StyleVenturella, Marta, Mattia Criscuoli, Fabio Carraro, Antonella Naldini, and Davide Zocco. 2021. "Interplay between Hypoxia and Extracellular Vesicles in Cancer and Inflammation" Biology 10, no. 7: 606. https://doi.org/10.3390/biology10070606