
Roles of Phosphate Solubilizing Microorganisms from Managing Soil Phosphorus Deficiency to Mediating Biogeochemical P Cycle


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
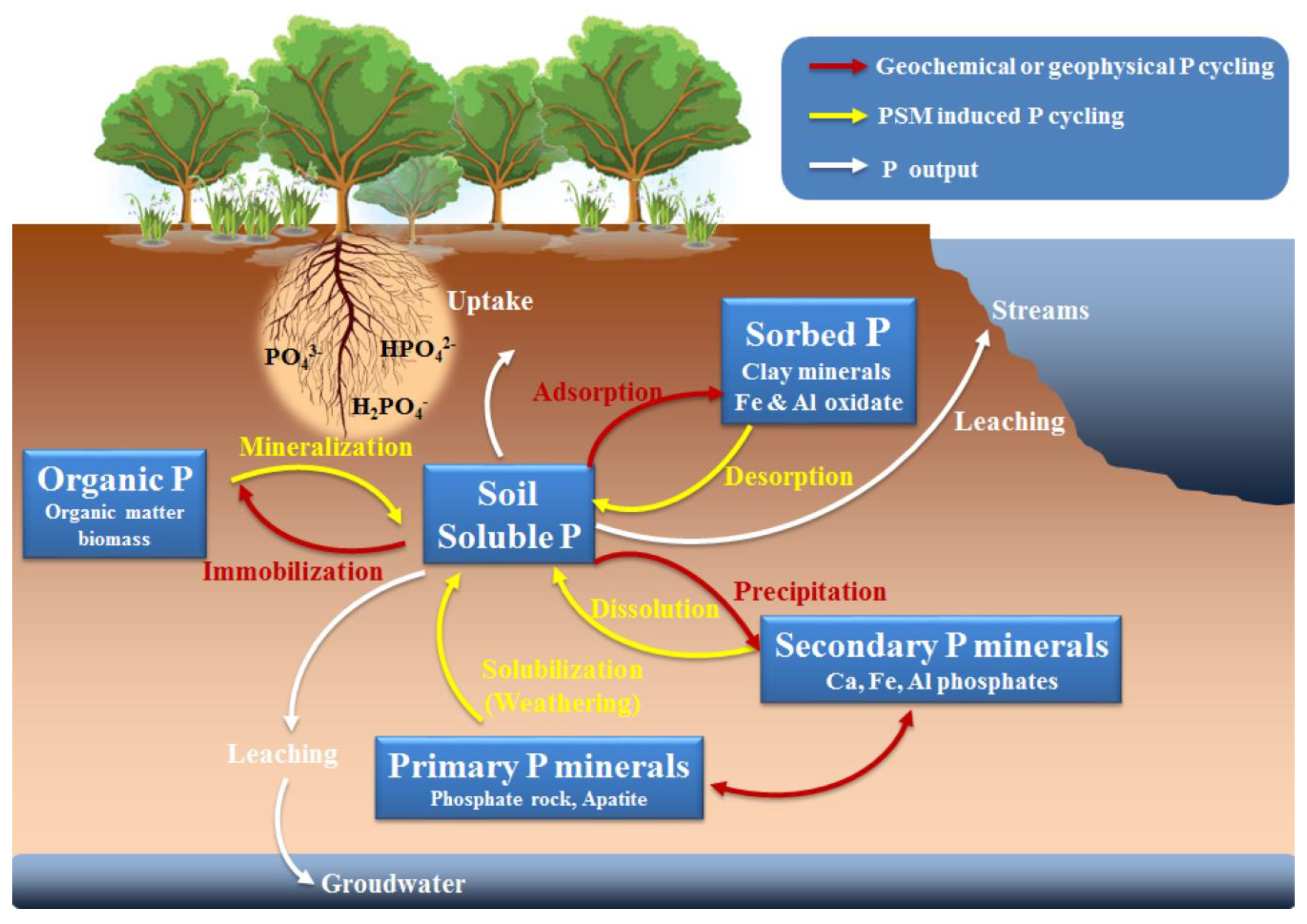
2. Basic P Forms and P Cycling in the Soil
3. PSM Enhance Soil P Cycle through Organic P Mineralization
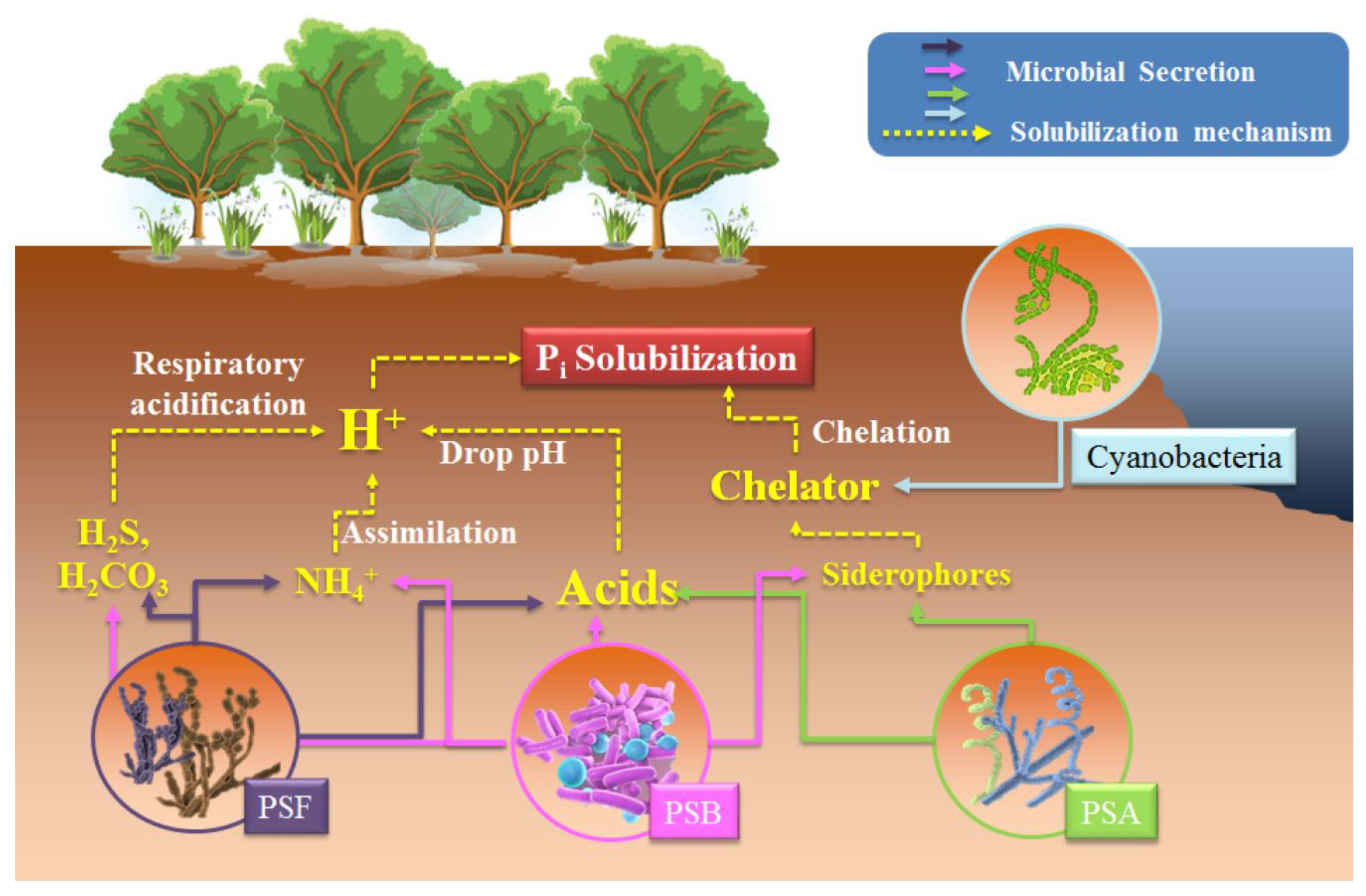
4. PSM-Mediated Inorganic P Solubilization to Enhance Soil Orthophosphate Contents
5. PSM-Derived P Desorption from Clay Minerals
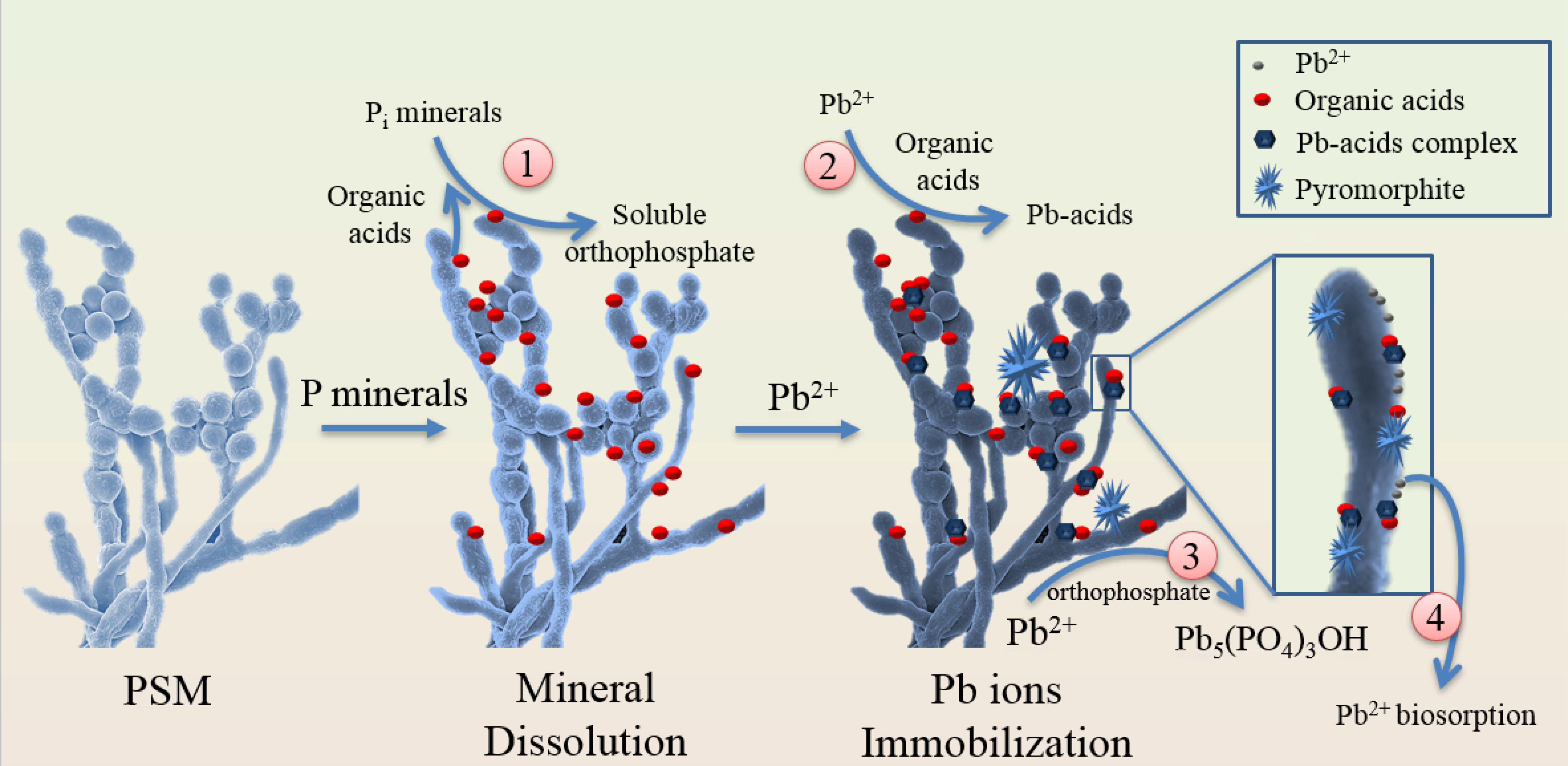
6. PSM-Induced Dissolution in Accelerating Metal Precipitation to form Secondary Pi Minerals
7. Effect of PSM on Pi Mineral Weathering and the Biogeochemical P Cycle
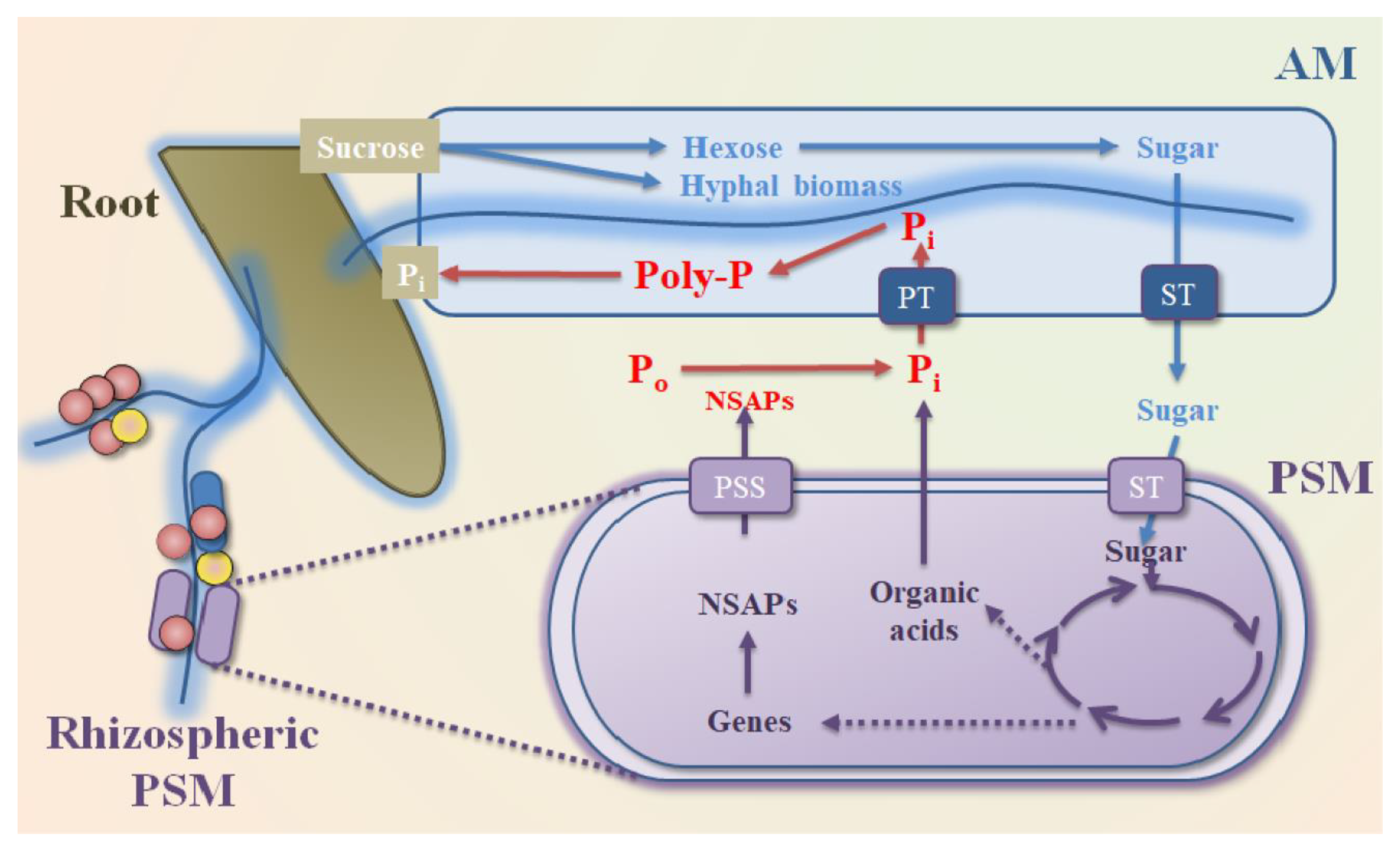
8. PSM Enhance P Uptake from Soil to Plant in the Rhizosphere Environment
9. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Billah, M.; Khan, M.; Bano, A.; Hassan, T.U.; Munir, A.; Gurmani, A.R. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Tamburini, F.; Pfahler, V.; Bunemann, E.K.; Guelland, K.; Bernasconi, S.M.; Frossard, E. Oxygen isotopes unravel the role of microorganisms in phosphate cycling in soils. Environ. Sci. Technol. 2012, 46, 5956–5962. [Google Scholar] [CrossRef]
- Tate, K.R. The biological transformation of P in soil. Plant Soil 1984, 76, 245–256. [Google Scholar] [CrossRef]
- Westheimer, F.H. Why nature chose phosphates. Science 1987, 235, 1173–1178. [Google Scholar] [CrossRef]
- Ma, Y.; Prasad, M.N.; Rajkumar, M.; Freitas, H. Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol. Adv. 2011, 29, 248–258. [Google Scholar] [CrossRef]
- Onodera, S.; Okuda, N.; Ban, S.H.; Saito, M.; Paytan, A.; Iwata, T. Phosphorus cycling in watersheds: From limnology to environmental science. Limnology 2020, 21, 327–328. [Google Scholar] [CrossRef]
- Xiong, C.; Guo, Z.; Chen, S.S.; Gao, Q.; Kishe, M.A.; Shen, Q. Understanding the pathway of phosphorus metabolism in urban household consumption system: A case study of Dar es Salaam, Tanzania. J. Clean. Prod. 2020, 274. [Google Scholar] [CrossRef]
- Liu, X.; Yuan, Z.; Liu, X.; Zhang, Y.; Hua, H.; Jiang, S. Historic Trends and Future Prospects of Waste Generation and Recycling in China’s Phosphorus Cycle. Environ. Sci. Technol. 2020, 54, 5131–5139. [Google Scholar] [CrossRef]
- Lu, G.Y.; Song, X.X.; Yu, Z.M.; Cao, X.H. Application of PAC-modified kaolin to mitigate Prorocentrum donghaiense: Effects on cell removal and phosphorus cycling in a laboratory setting. J. Appl. Phycol. 2017, 29, 917–928. [Google Scholar] [CrossRef]
- McMahon, K.D.; Read, E.K. Microbial Contributions to Phosphorus Cycling in Eutrophic Lakes and Wastewater. Annu. Rev. Microbiol. 2013, 67, 199–219. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [Green Version]
- Percival, L.M.E.; Bond, D.P.G.; Rakocinski, M.; Marynowski, L.; Hood, A.V.S.; Adatte, T.; Spangenberg, J.E.; Follmi, K.B. Phosphorus-cycle disturbances during the Late Devonian anoxic events. Global Planet Change 2020, 184, 103070. [Google Scholar] [CrossRef]
- Nguyen, T.B.; Lee, P.B.; Updyke, K.M.; Bones, D.L.; Laskin, J.; Laskin, A.; Nizkorodov, S.A. Formation of nitrogen- and sulfur-containing light-absorbing compounds accelerated by evaporation of water from secondary organic aerosols. J. Geophys. Res. Atmos. 2012, 117. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Balkanski, Y.; Boucher, O.; Ciais, P.; Penuelas, J.; Tao, S. Significant contribution of combustion-related emissions to the atmospheric phosphorus budget. Nat. Geosci. 2015, 8, 48–54. [Google Scholar] [CrossRef]
- Laakso, T.A.; Sperling, E.A.; Johnston, D.T.; Knoll, A.H. Ediacaran reorganization of the marine phosphorus cycle. Proc. Natl. Acad. Sci. USA 2020, 117, 11961–11967. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Jiang, S.; Sheng, H.; Liu, X.; Hua, H.; Liu, X.; Zhang, Y. Human Perturbation of the Global Phosphorus Cycle: Changes and Consequences. Environ. Sci. Technol. 2018, 52, 2438–2450. [Google Scholar] [CrossRef] [PubMed]
- Chapuis-Lardy, L.; Le Bayon, R.-C.; Brossard, M.; López-Hernández, D.; Blanchart, E. Role of Soil Macrofauna in Phosphorus Cycling. In Phosphorus in Action; Springer: Berlin/Heidelberg, Germany, 2011; pp. 199–213. [Google Scholar] [CrossRef] [Green Version]
- Mooshammer, M.; Hofhansl, F.; Frank, A.H.; Wanek, W.; Hammerle, I.; Leitner, S.; Schnecker, J.; Wild, B.; Watzka, M.; Keiblinger, K.M.; et al. Decoupling of microbial carbon, nitrogen, and phosphorus cycling in response to extreme temperature events. Sci. Adv. 2017, 3, e1602781. [Google Scholar] [CrossRef] [Green Version]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Global Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Elser, J.; Bennett, E. Phosphorus cycle A broken biogeochemical cycle. Nature 2011, 478, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Hébert, M.-P.; Fugère, V.; Gonzalez, A. The overlooked impact of rising glyphosate use on phosphorus loading in agricultural watersheds. Front. Ecol. Environ. 2019, 17, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Sheng, H.; Jiang, S.Y.; Yuan, Z.W.; Zhang, C.S.; Elser, J.J. Intensification of phosphorus cycling in China since the 1600s. Proc. Natl. Acad. Sci. USA 2016, 113, 2609–2614. [Google Scholar] [CrossRef] [Green Version]
- Hou, E.; Chen, C.; Luo, Y.; Zhou, G.; Kuang, Y.; Zhang, Y.; Heenan, M.; Lu, X.; Wen, D. Effects of climate on soil phosphorus cycle and availability in natural terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 3344–3356. [Google Scholar] [CrossRef] [Green Version]
- Gross, A.; Lin, Y.; Weber, P.K.; Pett-Ridge, J.; Silver, W.L. The role of soil redox conditions in microbial phosphorus cycling in humid tropical forests. Ecology 2020, 101, e02928. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.L.; Liu, J.; Jia, P.; Yang, T.T.; Zeng, Q.W.; Zhang, S.C.; Liao, B.; Shu, W.S.; Li, J.T. Novel phosphate-solubilizing bacteria enhance soil phosphorus cycling following ecological restoration of land degraded by mining. ISME J. 2020, 14, 1600–1613. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.E.; Simpson, R.J. Soil Microorganisms Mediating Phosphorus Availability. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.L.; Mao, X.L.; Van Zwieten, L.; Niazi, N.K.; Lu, K.P.; Bolan, N.S.; Wang, H.L. Wetting-drying cycles during a rice-wheat crop rotation rapidly (im)mobilize recalcitrant soil phosphorus. J. Soils Sediment. 2020. [Google Scholar] [CrossRef]
- Cross, A.F.; Schlesinger, W.H. A literature review and evaluation of the Hedley. Geoderma 1995, 64, 197–214. [Google Scholar] [CrossRef]
- Adams, M.A.; Pate, J.S. Availability of organic and inorganic forms of phosphorus to lupins (Lupinus spp.). Plant Soil 1992, 145, 107–113. [Google Scholar] [CrossRef]
- Fabianska, M.J.; Kozielska, B.; Konieczynski, J.; Bielaczyc, P. Occurrence of organic phosphates in particulate matter of the vehicle exhausts and outdoor environment—A case study. Environ. Pollut. 2019, 244, 351–360. [Google Scholar] [CrossRef]
- Hoffman, K.; Butt, C.M.; Webster, T.F.; Preston, E.V.; Hammel, S.C.; Makey, C.; Lorenzo, A.M.; Cooper, E.M.; Carignan, C.; Meeker, J.D.; et al. Temporal Trends in Exposure to Organophosphate Flame Retardants in the United States. Environ. Sci. Technol. Lett. 2017, 4, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Yuan, Z.W.; Zhang, Y.L.; Gao, L.M.; Liu, S.M. Life-cycle phosphorus use efficiency of the farming system in Anhui Province, Central China. Resour. Conserv. Recy. 2014, 83, 1–14. [Google Scholar] [CrossRef]
- Gebrim, F.D.; Novais, R.F.; da Siva, I.R.; Schulthais, F.; Vergutz, L.; Procopio, L.C.; Moreira, F.F.; de Jesus, G.L. Mobility of Inorganic and Organic Phosphorus Forms under Different Levels of Phosphate and Poultry Litter Fertilization in Soils. Rev. Bras. Cienc. Solo. 2010, 34, 1195–1205. [Google Scholar] [CrossRef]
- Maltais-Landry, G.; Scow, K.; Brennan, E. Soil phosphorus mobilization in the rhizosphere of cover crops has little effect on phosphorus cycling in California agricultural soils. Soil Biol. Biochem. 2014, 78, 255–262. [Google Scholar] [CrossRef]
- Hao, J.; Knoll, A.H.; Huang, F.; Schieber, J.; Hazen, R.M.; Daniel, I. Cycling phosphorus on the Archean Earth: Part II. Phosphorus limitation on primary production in Archean ecosystems. Geochim. Cosmochim. Acta 2020, 280, 360–377. [Google Scholar] [CrossRef]
- Prietzel, J.; Harrington, G.; Hausler, W.; Heister, K.; Werner, F.; Klysubun, W. Reference spectra of important adsorbed organic and inorganic phosphate binding forms for soil P speciation using synchrotron-based K-edge XANES spectroscopy. J. Synchrotron. Radiat. 2016, 23, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Hesterberg, D.; Zhou, W.; Hutchison, K.J.; Beauchemin, S.; Syers, D.E. XAFS study of adsorbed and mineral forms of phosphate. J. Synchrotron. Radiat. 1999, 6, 636–638. [Google Scholar] [CrossRef]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 15, 1–19. [Google Scholar] [CrossRef]
- Williams, J.D.H.; Syers, J.K.; Walker, T.W. Fractionation of soil inorganic phosphate by a modification of Chang and Jackson’s procedure. Soil Sci. Soc. Am. J. 1967, 31, 736–739. [Google Scholar] [CrossRef] [Green Version]
- Mathew, D.; Gireeshkumar, T.R.; Balachandran, K.K.; Udayakrishnan, P.B.; Shameem, K.; Deepulal, P.M.; Nair, M.; Madhu, N.V.; Muraleedharan, K.R. Influence of hypoxia on phosphorus cycling in Alappuzha mud banks, southwest coast of India. Reg. Stud. Mar Sci. 2020, 34, 101083. [Google Scholar] [CrossRef]
- Smil, V. Phosphorus in the Environment Natural Flows and Human Interferences. Annu. Rev. Energy Environ. 2000, 25, 53–88. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, K.; Brown, C.T.; Hug, L.A.; Sharon, I.; Castelle, C.J.; Probst, A.J.; Thomas, B.C.; Singh, A.; Wilkins, M.J.; Karaoz, U.; et al. Thousands of microbial genomes shed light on interconnected biogeochemical processes in an aquifer system. Nat. Commun. 2016, 7, 13219. [Google Scholar] [CrossRef] [PubMed]
- Dodd, R.J.; Sharpley, A.N. Recognizing the role of soil organic phosphorus in soil fertility and water quality. Resour. Conserv. Recy. 2015, 105, 282–293. [Google Scholar] [CrossRef]
- Tao, G.-C.; Tian, S.-J.; Cai, M.-Y.; Xie, G.-H. Phosphate-Solubilizing and -Mineralizing Abilities of Bacteria Isolated from Soils. Pedosphere 2008, 18, 515–523. [Google Scholar] [CrossRef]
- Müller, C.; Bünemann, E.K. A 33P tracing model for quantifying gross P transformation rates in soil. Soil Biol. Biochem. 2014, 76, 218–226. [Google Scholar] [CrossRef]
- Bi, Q.F.; Li, K.J.; Zheng, B.X.; Liu, X.P.; Li, H.Z.; Jin, B.J.; Ding, K.; Yang, X.R.; Lin, X.Y.; Zhu, Y.G. Partial replacement of inorganic phosphorus (P) by organic manure reshapes phosphate mobilizing bacterial community and promotes P bioavailability in a paddy soil. Sci. Total Environ. 2020, 703. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Song, C.; Wang, M.; Lai, D.Y.F.; Tariq, A.; Zeng, F.; Zhong, Q.; Wang, F.; Li, Z.; Peng, C. Long-term increase in rainfall decreases soil organic phosphorus decomposition in tropical forests. Soil Biol. Biochem. 2020. [Google Scholar] [CrossRef]
- Bai, J.H.; Yu, L.; Ye, X.F.; Yu, Z.B.; Guan, Y.N.; Li, X.W.; Cui, B.S.; Liu, X.H. Organic phosphorus mineralization characteristics in sediments from the coastal salt marshes of a Chinese delta under simulated tidal cycles. J. Soils Sediment. 2020, 20, 513–523. [Google Scholar] [CrossRef]
- Bi, Q.-F.; Zheng, B.-X.; Lin, X.-Y.; Li, K.-J.; Liu, X.-P.; Hao, X.-L.; Zhang, H.; Zhang, J.-B.; Jaisi, D.P.; Zhu, Y.-G. The microbial cycling of phosphorus on long-term fertilized soil: Insights from phosphate oxygen isotope ratios. Chem. Geol. 2018, 483, 56–64. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil. In Phosphorus in Action; Springer: Berlin/Heidelberg, Germany, 2011; pp. 215–243. [Google Scholar] [CrossRef]
- Zheng, L.; Ren, M.L.; Xie, E.; Ding, A.Z.; Liu, Y.; Deng, S.Q.; Zhang, D.Y. Roles of Phosphorus Sources in Microbial Community Assembly for the Removal of Organic Matters and Ammonia in Activated Sludge. Front. Microbiol. 2019, 10, 1023. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, H.; Fraga, R.; Gonzalez, T.; Bashan, Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil 2006, 287, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [Green Version]
- Passariello, C.; Schippa, S.; Iori, P.; Berlutti, F.; Thaller, M.C.; Rossolini, G.M. The molecular class C acid phosphatase of Chryseobacterium meningosepticum (OlpA) is a broad-spectrum nucleotidase with preferential activity on 5′-nucleotides. Biochim. Biophys. Acta 2003, 1648, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.H.; Liu, X.D.; Yin, H.Q.; Liang, Y.L.; Liu, H.W.; Miao, B.; Peng, Q.Q.; Meng, D.L.; Wang, S.Q.; Yang, J.J.; et al. The utilization of biomineralization technique based on microbial induced phosphate precipitation in remediation of potentially toxic ions contaminated soil: A mini review. Ecotox. Environ. Safety 2020, 191, 110009. [Google Scholar] [CrossRef]
- Xie, E.; Su, Y.P.; Deng, S.Q.; Kontopyrgou, M.; Zhang, D.Y. Significant influence of phosphorus resources on the growth and alkaline phosphatase activities of Microcystis aeruginosa. Environ. Pollut. 2021, 268, 115807. [Google Scholar] [CrossRef]
- Xie, E.; Li, F.F.; Wang, C.Z.; Shi, W.; Huang, C.; Fa, K.Y.; Zhao, X.; Zhang, D.Y. Roles of sulfur compounds in growth and alkaline phosphatase activities of Microcystis aeruginosa under phosphorus deficiency stress. Environ. Sci. Pollut. R. 2020, 27, 21533–21541. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Liang, X.; Csetenyi, L.; Gadd, G.M. Lead Bioprecipitation by Yeasts Utilizing Organic Phosphorus Substrates. Geomicrobiol. J. 2016, 33, 294–307. [Google Scholar] [CrossRef]
- Farias, N.; Almeida, I.; Meneses, C. New Bacterial Phytase through Metagenomic Prospection. Molecules 2018, 23, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kour, D.; Rana, K.L.; Kaur, T.; Yadav, N.; Yadav, A.N.; Kumar, M.; Kumar, V.; Dhaliwal, H.S.; Saxena, A.K. Biodiversity, current developments and potential biotechnological applications of phosphorus-solubilizing and -mobilizing microbes: A review. Pedosphere 2021, 31, 43–75. [Google Scholar] [CrossRef]
- Cong, W.F.; Suriyagoda, L.D.B.; Lambers, H. Tightening the Phosphorus Cycle through Phosphorus-Efficient Crop Genotypes. Trends Plant Sci. 2020. [Google Scholar] [CrossRef]
- Fixen, P.E.; Johnston, A.M. World fertilizer nutrient reserves: A view to the future. J. Sci. Food Agric. 2012, 92, 1001–1005. [Google Scholar] [CrossRef]
- Turan, M.; Ataoğlu, N.; Şahιn, F. Evaluation of the Capacity of Phosphate Solubilizing Bacteria and Fungi on Different Forms of Phosphorus in Liquid Culture. J. Sustain. Agr. 2006, 28, 99–108. [Google Scholar] [CrossRef]
- Kucey, R.M. Increased Phosphorus Uptake by Wheat and Field Beans Inoculated with a Phosphorus-Solubilizing Penicillium bilaji Strain and with Vesicular-Arbuscular Mycorrhizal Fungi. Appl. Environ. Microbiol. 1987, 53, 2699–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.T.; Lee, L.Y.; Tai, C.Y.; Hung, C.H.; Chang, Y.S.; Wolfram, J.H.; Rogers, R.; Goldstein, A.H. Cloning of an Erwinia herbicola gene necessary for gluconic acid production and enhanced mineral phosphate solubilization in Escherichia coli HB101: Nucleotide sequence and probable involvement in biosynthesis of the coenzyme pyrroloquinoline quinone. J. Bacteriol. 1992, 174, 5814–5819. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Xiao, C.Q.; Feng, B.; Chi, R. Phosphate rock solubilization and the potential for lead immobilization by a phosphate-solubilizing bacterium (Pseudomonas sp.). J. Environ. Sci. Heal. A 2020, 55, 411–420. [Google Scholar] [CrossRef]
- Alaylar, B.; Egamberdieva, D.; Gulluce, M.; Karadayi, M.; Arora, N.K. Integration of molecular tools in microbial phosphate solubilization research in agriculture perspective. World J. Microbiol. Biotechnol. 2020, 36, 93. [Google Scholar] [CrossRef]
- Halder, A.K.; Chakrabartty, P.K. Solubilization of inorganic phosphate byRhizobium. Folia Microbiol. 1993, 38, 325–330. [Google Scholar] [CrossRef]
- Sperber, J.I. Solution of mineral phosphate by soil bacteria. Nature 1957, 180, 994–995. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, A.; Ahemad, M.; Oves, M.; Ahmad, E.; Khan, M.S. Role of Phosphate-Solubilizing Bacteria in Legume Improvement. In Microbes for Legume Improvement; Springer: Cham, Switzerland, 2010; pp. 273–292. [Google Scholar] [CrossRef]
- Jiang, Y.F.; Tian, J.; Ge, F. New Insight into Carboxylic Acid Metabolisms and pH Regulations During Insoluble Phosphate Solubilisation Process by Penicillium oxalicum PSF-4. Curr. Microbiol. 2020, 77, 4095–4103. [Google Scholar] [CrossRef]
- Mackay, J.E.; Cavagnaro, T.R.; Müller Stöver, D.S.; Macdonald, L.M.; Grønlund, M.; Jakobsen, I. A key role for arbuscular mycorrhiza in plant acquisition of P from sewage sludge recycled to soil. Soil Biol. Biochem. 2017, 115, 11–20. [Google Scholar] [CrossRef]
- Zai, X.M.; Zhang, H.S.; Hao, Z.P. Effects of Arbuscular Mycorrhizal Fungi and Phosphate-Solubilizing Fungus on the Rooting, Growth and Rhizosphere Niche of Beach Plum (Prunus maritima) Cuttings in a Phosphorus-deficient Soil. J. Am. Pomol. Soc. 2017, 71, 226–235. [Google Scholar]
- Jog, R.; Pandya, M.; Nareshkumar, G.; Rajkumar, S. Mechanism of phosphate solubilization and antifungal activity of Streptomyces spp. isolated from wheat roots and rhizosphere and their application in improving plant growth. Microbiology 2014, 160, 778–788. [Google Scholar] [CrossRef] [Green Version]
- Hamdali, H.; Smirnov, A.; Esnault, C.; Ouhdouch, Y.; Virolle, M.J. Physiological studies and comparative analysis of rock phosphate solubilization abilities of Actinomycetales originating from Moroccan phosphate mines and of Streptomyces lividans. Appl. Soil Ecol. 2010, 44, 24–31. [Google Scholar] [CrossRef]
- Hamdali, H.; Bouizgarne, B.; Hafidi, M.; Lebrihi, A.; Virolle, M.J.; Ouhdouch, Y. Screening for rock phosphate solubilizing Actinomycetes from Moroccan phosphate mines. Appl. Soil. Ecol. 2008, 38, 12–19. [Google Scholar] [CrossRef]
- Chaiharn, M.; Pathom-aree, W.; Sujada, N.; Lumyong, S. Characterization of Phosphate Solubilizing Streptomyces as a Biofertilizer. Chiang Mai. J. Sci. 2018, 45, 701–716. [Google Scholar]
- Farhat, M.B.; Boukhris, I.; Chouayekh, H. Mineral phosphate solubilization by Streptomyces sp. CTM396 involves the excretion of gluconic acid and is stimulated by humic acids. Fems Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yandigeri, M.S.; Yadav, A.K.; Srinivasan, R.; Kashyap, S.; Pabbi, S. Studies on mineral phosphate solubilization by cyanobacteria Westiellopsis and Anabaena. Microbiology 2011, 80, 558–565. [Google Scholar] [CrossRef]
- Baumann, K.; Jung, P.; Samolov, E.; Lehnert, L.W.; Budel, B.; Karsten, U.; Bendix, J.; Achilles, S.; Schermer, M.; Matus, F.; et al. Biological soil crusts along a climatic gradient in Chile: Richness and imprints of phototrophic microorganisms in phosphorus biogeochemical cycling. Soil Biol. Biochem. 2018, 127, 286–300. [Google Scholar] [CrossRef]
- Nahas, E. Factors determining rock phosphate solubilization by microorganisms isolated from soil. World J. Microbiol. Biotechnol. 1996, 12, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, L.; Geerts, S.; Van Caneghem, J. Closing the phosphorus cycle: Multi-criteria techno-economic optimization of phosphorus extraction from wastewater treatment sludge ash. Sci. Total Environ. 2020, 713, 135543. [Google Scholar] [CrossRef]
- Illmer, P.; Schinner, F. Solubilization of inorganic calcium phosphates-solubilization mechanisms. Soil Biol. Biochem. 1995, 27, 257–263. [Google Scholar] [CrossRef]
- Yi, Y.; Huang, W.; Ge, Y. Exopolysaccharide: A novel important factor in the microbial dissolution of tricalcium phosphate. World J. Microbiol. Biotechnol. 2007, 24, 1059–1065. [Google Scholar] [CrossRef]
- Whitton, B.A.; Grainger, S.L.J.; Hawley, G.R.W.; Simon, J.W. Cell-bound and extracellular phosphatase activities of cyanobacterial isolates. Microb. Ecol. 1991, 21, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lin, Q.; Li, B. The solubilization of four insoluble phosphates by some microorganisms. Wei Sheng Wu Xue Bao 2002, 42, 236–241. [Google Scholar]
- Osorno, L.; Osorio, N.W.; Habte, M. Phosphate desorption by a soil fungus in selected Hawaiian soils differing in their mineralogy. Trop. Agric. 2018, 95, 154–166. [Google Scholar]
- Osorio, N.W.; Habte, M. Phosphate desorption from the surface of soil mineral particles by a phosphate-solubilizing fungus. Biol. Fert. Soils 2012, 49, 481–486. [Google Scholar] [CrossRef]
- He, Z.; Zhu, J. Microbial utilization and transformation of phosphate adsorbed by variable charge minerals. Soil Biol. Biochem. 1998, 30, 917–923. [Google Scholar] [CrossRef]
- Eriksson, A.K.; Gustafsson, J.P.; Hesterberg, D. Phosphorus speciation of clay fractions from long-term fertility experiments in Sweden. Geoderma 2015, 241-242, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.J.; Zhao, Y.P.; Chen, Y.Y.; Dong, X.M.; Wang, M.Y.; Wang, G.X. Sulfur cycling in freshwater sediments: A cryptic driving force of iron deposition and phosphorus mobilization. Sci. Total Environ. 2019, 657, 1294–1303. [Google Scholar] [CrossRef]
- de Campos, M.; Antonangelo, J.A.; Alleoni, L.R.F. Phosphorus sorption index in humid tropical soils. Soil Till. Res. 2016, 156, 110–118. [Google Scholar] [CrossRef]
- Qiao, Z.; Hong, J.; Li, L.; Liu, C. Effect of Phosphobacterias on Nutrient, Enzyme Activities and Phosphorus Adsorption—Desorption Characteristics in a Reclaimed Soil. J. Soil Water Conserv. 2017, 31, 166–171. [Google Scholar]
- Osorio, N.W.; Habte, M. Soil Phosphate Desorption Induced by a Phosphate-Solubilizing Fungus. Commun. Soil Sci. Plant 2014, 45, 451–460. [Google Scholar] [CrossRef]
- Hoberg, E.; Marschner, P.; Lieberei, R. Organic acid exudation and pH changes by Gordonia sp. and Pseudomonas fluorescens grown with P adsorbed to goethite. Microbiol. Res. 2005, 160, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Andrade, F.V.; Mendonça, E.S.; Silva, I.R. Organic Acids and Diffusive Flux of Organic and Inorganic Phosphorus in Sandy-Loam and Clayey Latosols. Commun. Soil Sci. Plant 2013, 44, 1211–1223. [Google Scholar] [CrossRef]
- Oburger, E.; Leitner, D.; Jones, D.L.; Zygalakis, K.C.; Schnepf, A.; Roose, T. Adsorption and desorption dynamics of citric acid anions in soil. Eur. J. Soil Sci. 2011, 62, 733–742. [Google Scholar] [CrossRef]
- Suriyagoda, L.B.D.; Tibbett, M.; Edmonds-Tibbett, T.; Cawthray, G.R.; Ryan, M.H. Poor regulation of phosphorus uptake and rhizosphere carboxylates in three phosphorus-hyperaccumulating species of Ptilotus. Plant Soil 2015, 402, 145–158. [Google Scholar] [CrossRef]
- Pastore, G.; Kaiser, K.; Kernchen, S.; Spohn, M. Microbial release of apatite- and goethite-bound phosphate in acidic forest soils. Geoderma 2020, 370, 114360. [Google Scholar] [CrossRef]
- Li, C.K.; Li, Q.S.; Wang, Z.P.; Ji, G.N.; Zhao, H.; Gao, F.; Su, M.; Jiao, J.G.; Li, Z.; Li, H.X. Environmental fungi and bacteria facilitate lecithin decomposition and the transformation of phosphorus to apatite. Sci. Rep. 2019, 9, 15291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Deng, R.; Wan, J.; Zeng, G.; Xue, W.; Wen, X.; Zhou, C.; Hu, L.; Liu, X.; Xu, P.; et al. Remediation of lead-contaminated sediment by biochar-supported nano-chlorapatite: Accompanied with the change of available phosphorus and organic matters. J. Hazard. Mater. 2018, 348, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Remmers, J.C.; Saakes, M.; van der Weijden, R.D.; Buisman, C.J.N. Is There a Precipitation Sequence in Municipal Wastewater Induced by Electrolysis? Environ. Sci. Technol. 2018, 52, 8399–8407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeJong, J.T.; Mortensen, B.M.; Martinez, B.C.; Nelson, D.C. Bio-mediated soil improvement. Ecol. Eng. 2010, 36, 197–210. [Google Scholar] [CrossRef]
- Eighmy, T.T.; Kinner, A.E.; Shaw, E.L.; Eusden, J.D.; Francis, C.A. Chlorapatite (Ca5(PO4)3Cl) Characterization by XPS: An Environmentally Important Secondary Mineral. Surf. Sci. Spectra 1999, 6, 210–218. [Google Scholar] [CrossRef]
- Mutschke, A.; Wylezich, T.; Ritter, C.; Karttunen, A.J.; Kunkel, N. An Unprecedented Fully H-Substituted Phosphate Hydride Sr5(PO4)3H Expanding the Apatite Family. Eur. J. Inorg. Chem. 2019, 2019, 5073–5076. [Google Scholar] [CrossRef] [Green Version]
- Hirschler, A.; Lucas, J.; Hubert, J.C. Apatite genesis: A biologically induced or biologically controlled mineral formation process? Geomicrobiol. J. 1990, 8, 47–56. [Google Scholar] [CrossRef]
- Chang, S.J.; Blake, R.E.; Stout, L.M.; Kim, S.J. Oxygen isotope, micro-textural and molecular evidence for the role of microorganisms in formation of hydroxylapatite in limestone caves, South Korea. Chem. Geol. 2010, 276, 209–224. [Google Scholar] [CrossRef]
- Deng, S.; Zhang, C.; Dang, Y.; Collins, R.N.; Kinsela, A.S.; Tian, J.; Holmes, D.E.; Li, H.; Qiu, B.; Cheng, X.; et al. Iron Transformation and Its Role in Phosphorus Immobilization in a UCT-MBR with Vivianite Formation Enhancement. Environ. Sci. Technol. 2020, 54, 12539–12549. [Google Scholar] [CrossRef]
- Fontaine, L.; Thiffault, N.; Paré, D.; Fortin, J.A.; Piché, Y. Phosphate-solubilizing bacteria isolated from ectomycorrhizal mycelium of Picea glaucaare highly efficient at fluorapatite weathering. Botany 2016, 94, 1183–1193. [Google Scholar] [CrossRef] [Green Version]
- Reyes, I.; Bernier, L.; Simard, R.R.; Tanguay, P.; Antoun, H. Characteristics of phosphate solubilization by an isolate of a tropical Penicillium rugulosum and two UV-induced mutants. FEMS Microbiol. Ecol. 1999, 28, 291–295. [Google Scholar] [CrossRef]
- Iuliano, M.; Ciavatta, L.; De Tommaso, G. On the Solubility Constant of Strengite. Soil Sci. Soc. Am. J. 2007, 71, 1137–1140. [Google Scholar] [CrossRef]
- Ae, N.; Otani, T.; Makino, T.; Tazawa, J. Role of cell wall of groundnut roots in solubilizing sparingly soluble phosphorus in soil. Plant Soil 1996, 186, 197–204. [Google Scholar] [CrossRef]
- Kranzler, C.; Kessler, N.; Keren, N.; Shaked, Y. Enhanced ferrihydrite dissolution by a unicellular, planktonic Cyanobacterium: A biological contribution to particulate iron bioavailability. Environ. Microbiol 2016, 18, 5101–5111. [Google Scholar] [CrossRef]
- Chen, Z.; Pan, X.; Chen, H.; Guan, X.; Lin, Z. Biomineralization of Pb(II) into Pb-hydroxyapatite induced by Bacillus cereus 12-2 isolated from Lead-Zinc mine tailings. J. Hazard. Mater. 2016, 301, 531–537. [Google Scholar] [CrossRef]
- Zhang, K.J.; Xue, Y.W.; Zhang, J.Q.; Hu, X.L. Removal of lead from acidic wastewater by bio-mineralized bacteria with pH self-regulation. Chemosphere 2020, 241, 125041. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Wang, S.L.; Tang, X.C.; Fan, X.P.; Yang, S.; Yao, L.G.; Li, Y.D.; Han, H. Isolation of urease-producing bacteria and their effects on reducing Cd and Pb accumulation in lettuce (Lactuca sativa L.). Environ. Sci. Pollut. R. 2020, 27, 8707–8718. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Bolan, N.; Megharaj, M.; Naidu, R.; Chung, J.W. Bacterial-Assisted Immobilization of Lead in Soils: Implications for Remediation. Pedologist 2011, 54, 162–174. [Google Scholar]
- White, D.A.; Hafsteinsdottir, E.G.; Gore, D.B.; Thorogood, G.; Stark, S.C. Formation and stability of Pb-, Zn- and Cu-PO(4) phases at low temperatures: Implications for heavy metal fixation in polar environments. Environ. Pollut. 2012, 161, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Naik, M.M.; Dubey, S.K. Lead resistant bacteria: Lead resistance mechanisms, their applications in lead bioremediation and biomonitoring. Ecotoxicol. Environ. Saf. 2013, 98, 1–7. [Google Scholar] [CrossRef]
- Zhao, W.-W.; Zhu, G.; Daugulis, A.J.; Chen, Q.; Ma, H.-Y.; Zheng, P.; Liang, J.; Ma, X.-k. Removal and biomineralization of Pb2+ in water by fungus Phanerochaete chrysoporium. J. Clean. Prod. 2020, 260. [Google Scholar] [CrossRef]
- Xu, Y.; Schwartz, F.W. Lead immobilization by hydroxyapatite in aqueous solutions. J. Contam. Hydrol. 1994, 15, 187–206. [Google Scholar] [CrossRef]
- Xu, J.C.; Huang, L.M.; Chen, C.Y.; Wang, J.; Long, X.X. Effective lead immobilization by phosphate rock solubilization mediated by phosphate rock amendment and phosphate solubilizing bacteria. Chemosphere 2019, 237, 124540. [Google Scholar] [CrossRef]
- Schütze, E.; Weist, A.; Klose, M.; Wach, T.; Schumann, M.; Nietzsche, S.; Merten, D.; Baumert, J.; Majzlan, J.; Kothe, E. Taking nature into lab: Biomineralization by heavy metal-resistant Streptomycetes in soil. Biogeosciences 2013, 10, 3605–3614. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Qi, H.-W.; Hu, R.-Z. Element mobilization and redistribution under extreme tropical weathering of basalts from the Hainan Island, South China. J. Asian Earth Sci. 2018, 158, 80–102. [Google Scholar] [CrossRef]
- Ouahmane, L.; Revel, J.C.; Hafidi, M.; Thioulouse, J.; Prin, Y.; Galiana, A.; Dreyfus, B.; Duponnois, R. Responses of Pinus halepensis growth, soil microbial catabolic functions and phosphate-solubilizing bacteria after rock phosphate amendment and ectomycorrhizal inoculation. Plant Soil 2009, 320, 169–179. [Google Scholar] [CrossRef]
- Buss, H.L.; Mathur, R.; White, A.F.; Brantley, S.L. Phosphorus and iron cycling in deep saprolite, Luquillo Mountains, Puerto Rico. Chem. Geol. 2010, 269, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Garland, G.; Bunemann, E.K.; Oberson, A.; Frossard, E.; Snapp, S.; Chikowo, R.; Six, J. Phosphorus cycling within soil aggregate fractions of a highly weathered tropical soil: A conceptual model. Soil Biol. Biochem. 2018, 116, 91–98. [Google Scholar] [CrossRef]
- Rathi, M.; Gaur, N. Phosphate solubilizing bacteria as biofertilizer and its applications. J. Pharm. Res. 2016, 10, 146–148. [Google Scholar]
- Berhe, A.A.; Barnes, R.T.; Six, J.; Marin-Spiotta, E. Role of Soil Erosion in Biogeochemical Cycling of Essential Elements: Carbon, Nitrogen, and Phosphorus. Annu. Rev. Earth Planet. Sci. 2018, 46, 521–548. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, H.; Cravotta, C.A.; Dong, Q.; Xiang, X. Dissolution of Fluorapatite by Pseudomonas fluorescens P35 Resulting in Fluorine Release. Geomicrobiol. J. 2016, 1–13. [Google Scholar] [CrossRef]
- Bashan, Y.; Kamnev, A.A.; de-Bashan, L.E. Tricalcium phosphate is inappropriate as a universal selection factor for isolating and testing phosphate-solubilizing bacteria that enhance plant growth: A proposal for an alternative procedure. Biol. Fert. Soils 2012, 49, 465–479. [Google Scholar] [CrossRef]
- Qiu, S.; Lian, B. Weathering of phosphorus-bearing mineral powder and calcium phosphate by Aspergillus niger. Chinese J. Geochem. 2012, 31, 390–397. [Google Scholar] [CrossRef]
- Hanane, H. Isolation and characterization of rock phosphate solubilizing actinobacteria from a Togolese phosphate mine. Afr. J. Biotechnol. 2011, 11. [Google Scholar] [CrossRef]
- Puente, M.E.; Bashan, Y.; Li, C.Y.; Lebsky, V.K. Microbial populations and activities in the rhizoplane of rock-weathering desert plants. I. Root colonization and weathering of igneous rocks. Plant Biol. 2004, 6, 629–642. [Google Scholar] [CrossRef] [Green Version]
- Mendes, G.d.O.; Galvez, A.; Vassileva, M.; Vassilev, N. Fermentation liquid containing microbially solubilized P significantly improved plant growth and P uptake in both soil and soilless experiments. Appl. Soil. Ecol. 2017, 117, 208–211. [Google Scholar] [CrossRef]
- Ahemad, M. Phosphate-solubilizing bacteria-assisted phytoremediation of metalliferous soils: A review. 3 Biotech 2015, 5, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Bonilla, G.A.; Durrer, A.; Cardoso, E.J.B.N. Use of compost and phosphate-solubilizing bacteria affect sugarcane mineral nutrition, phosphorus availability, and the soil bacterial community. Appl. Soil Ecol. 2021, 157, 103760. [Google Scholar] [CrossRef]
- Raymond, N.S.; Gomez-Munoz, B.; van der Bom, F.J.T.; Nybroe, O.; Jensen, L.S.; Muller-Stover, D.S.; Oberson, A.; Richardson, A.E. Phosphate-solubilising microorganisms for improved crop productivity: A critical assessment. New Phytol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Meyer, G.; Maurhofer, M.; Frossard, E.; Gamper, H.A.; Mäder, P.; Mészáros, É.; Schönholzer-Mauclaire, L.; Symanczik, S.; Oberson, A. Pseudomonas protegens CHA0 does not increase phosphorus uptake from 33P labeled synthetic hydroxyapatite by wheat grown on calcareous soil. Soil Biol. Biochem. 2019, 131, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Romero-Perdomo, F.; Beltrán, I.; Mendoza-Labrador, J.; Estrada-Bonilla, G.; Bonilla, R. Phosphorus Nutrition and Growth of Cotton Plants Inoculated With Growth-Promoting Bacteria Under Low Phosphate Availability. Front. Sustain. Food Syst. 2021, 4. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A.; Oves, M. Role of Plant Growth Promoting Rhizobacteria in the Remediation of Metal Contaminated Soils: A Review. In Organic Farming, Pest Control and Remediation of Soil Pollutants: Organic Farming, Pest Control and Remediation of Soil Pollutants; Lichtfouse, E., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 319–350. [Google Scholar] [CrossRef]
- Yahaghi, Z.; Shirvani, M.; Nourbakhsh, F.; de la Pena, T.C.; Pueyo, J.J.; Talebi, M. Isolation and Characterization of Pb-Solubilizing Bacteria and Their Effects on Pb Uptake by Brassica juncea: Implications for Microbe-Assisted Phytoremediation. J. Microbiol. Biotechnol. 2018, 28, 1156–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.X.; Ding, K.; Yang, X.R.; Wadaan, M.A.M.; Hozzein, W.N.; Penuelas, J.; Zhu, Y.G. Straw biochar increases the abundance of inorganic phosphate solubilizing bacterial community for better rape (Brassica napus) growth and phosphate uptake. Sci. Total Environ. 2019, 647, 1113–1120. [Google Scholar] [CrossRef]
- Kumar, A.; Teja, E.S.; Mathur, V.; Kumari, R. Phosphate-Solubilizing Fungi: Current Perspective, Mechanisms and Potential Agricultural Applications. In Agriculturally Important Fungi for Sustainable Agriculture: Volume 1: Perspective for Diversity and Crop Productivity; Yadav, A.N., Mishra, S., Kour, D., Yadav, N., Kumar, A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 121–141. [Google Scholar] [CrossRef]
- Smith, S.E.; Jakobsen, I.; Gronlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.K.; Tewari, S.; Singh, R. Multifaceted Plant-Associated Microbes and Their Mechanisms Diminish the Concept of Direct and Indirect PGPRs; Springer: New Delhi, India, 2013; pp. 411–449. [Google Scholar] [CrossRef]
- Tisserant, E.; Malbreil, M.; Kuo, A.; Kohler, A.; Symeonidi, A.; Balestrini, R.; Charron, P.; Duensing, N.; Frei dit Frey, N.; Gianinazzi-Pearson, V.; et al. Genome of an arbuscular mycorrhizal fungus provides insight into the oldest plant symbiosis. Proc. Natl. Acad. Sci. USA 2013, 110, 20117–20122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xu, M.; Liu, Y.; Zhang, F.; Hodge, A.; Feng, G. Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytol. 2016, 210, 1022–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Zhang, L.; Zhou, J.; George, T.S.; Feng, G. Arbuscular mycorrhizal fungi enhance mineralisation of organic phosphorus by carrying bacteria along their extraradical hyphae. New Phytol. 2020. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, G.; Declerck, S. Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phytate mineralization by a phosphate solubilizing bacterium. ISME J. 2018, 12, 2339–2351. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Shao, C.; Sun, H.; Liu, Z.B.; Guan, Y.M.; Wu, L.J.; Zhang, L.L.; Pan, X.X.; Zhang, Z.H.; Zhang, Y.Y.; et al. Arbuscular mycorrhizal fungi biofertilizer improves American ginseng (Panax quinquefolius L.) growth under the continuous cropping regime. Geoderma 2020, 363, 114155. [Google Scholar] [CrossRef]
- Hao, X.L.; Zhu, Y.G.; Nybroe, O.; Nicolaisen, M.H. The Composition and Phosphorus Cycling Potential of Bacterial Communities Associated With Hyphae of Penicillium in Soil Are Strongly Affected by Soil Origin. Front. Microbiol. 2020, 10, 2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.K.; Kim, S.B.; Lyou, E.S.; Lee, T.K. Microbial phenomics linking the phenotype to function: The potential of Raman spectroscopy. J. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Z.; Bi, Q.F.; Yang, K.; Zheng, B.X.; Pu, Q.; Cui, L. D2O-Isotope-Labeling Approach to Probing Phosphate-Solubilizing Bacteria in Complex Soil Communities by Single-Cell Raman Spectroscopy. Anal. Chem. 2019, 91, 2239–2246. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, J.; Ge, F.; Zhang, D.; Deng, S.; Liu, X. Roles of Phosphate Solubilizing Microorganisms from Managing Soil Phosphorus Deficiency to Mediating Biogeochemical P Cycle. Biology 2021, 10, 158. https://doi.org/10.3390/biology10020158
Tian J, Ge F, Zhang D, Deng S, Liu X. Roles of Phosphate Solubilizing Microorganisms from Managing Soil Phosphorus Deficiency to Mediating Biogeochemical P Cycle. Biology. 2021; 10(2):158. https://doi.org/10.3390/biology10020158
Chicago/Turabian StyleTian, Jiang, Fei Ge, Dayi Zhang, Songqiang Deng, and Xingwang Liu. 2021. "Roles of Phosphate Solubilizing Microorganisms from Managing Soil Phosphorus Deficiency to Mediating Biogeochemical P Cycle" Biology 10, no. 2: 158. https://doi.org/10.3390/biology10020158