
Plasticity in Cervical Motor Circuits following Spinal Cord Injury and Rehabilitation


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
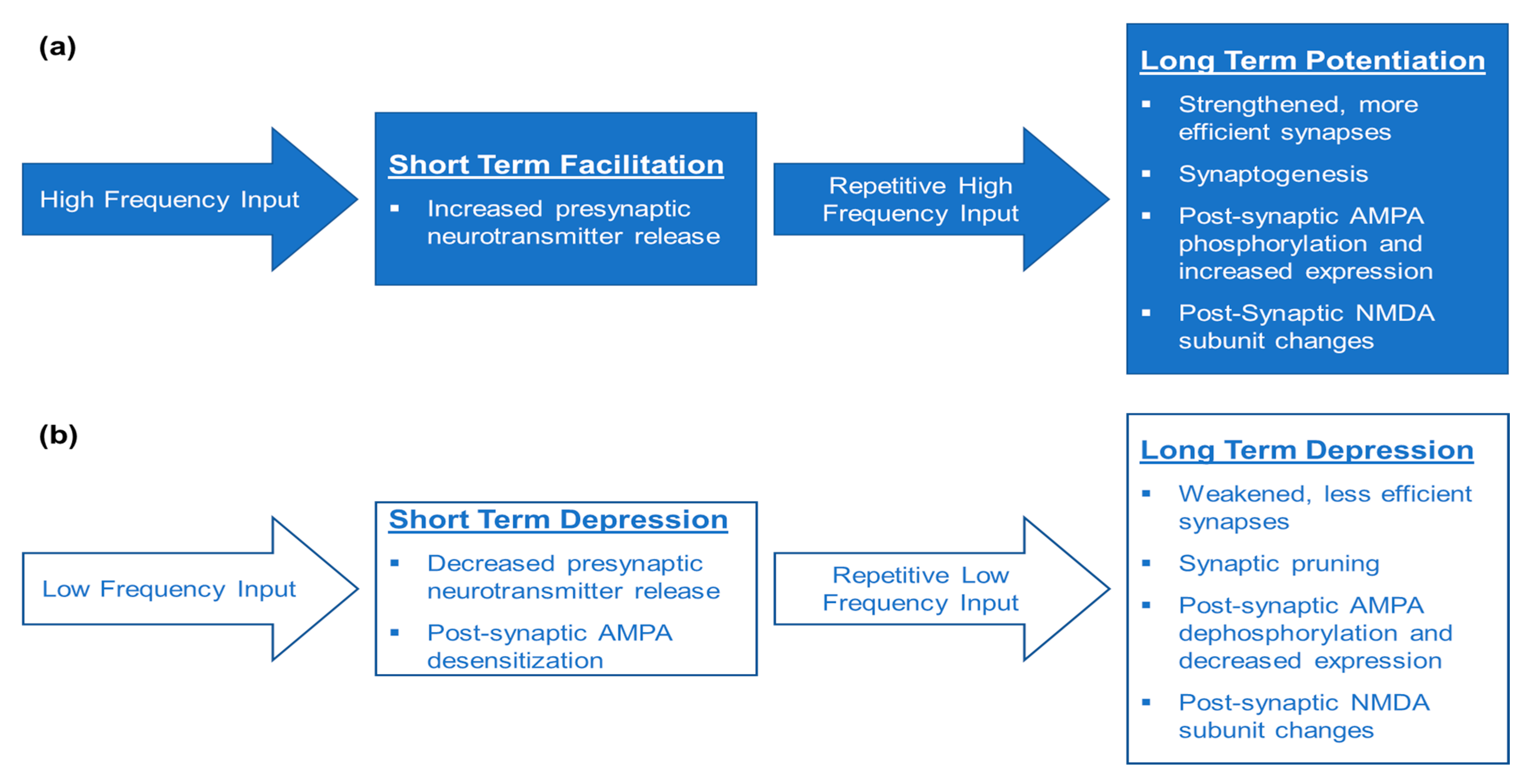
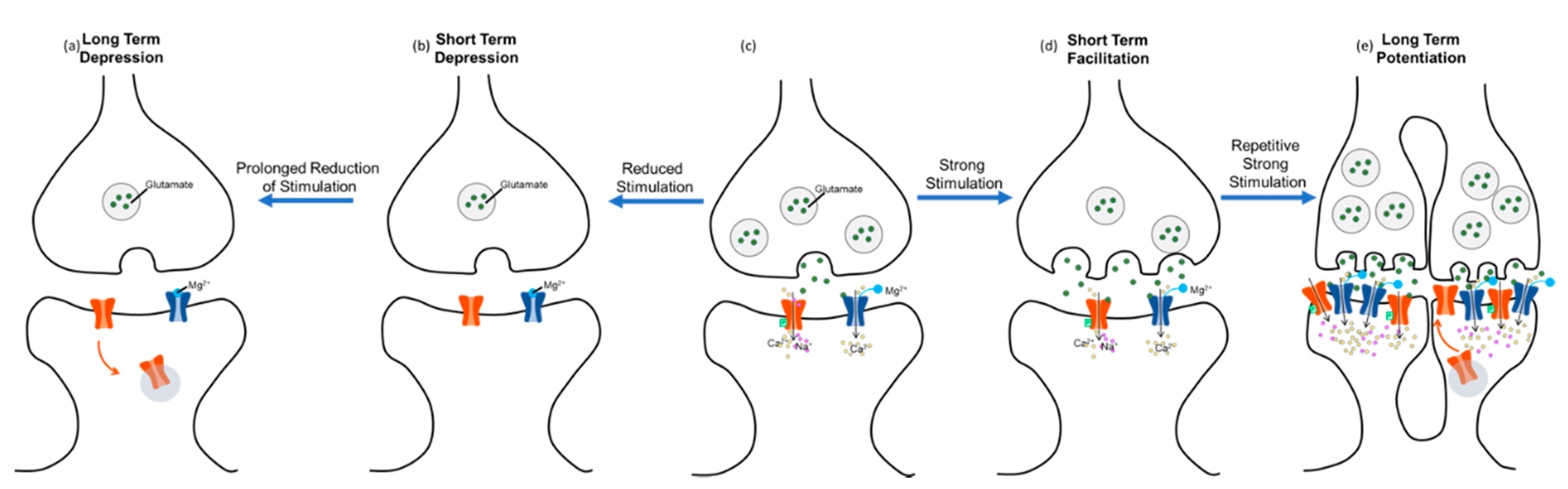
2. What Is Neuroplasticity?
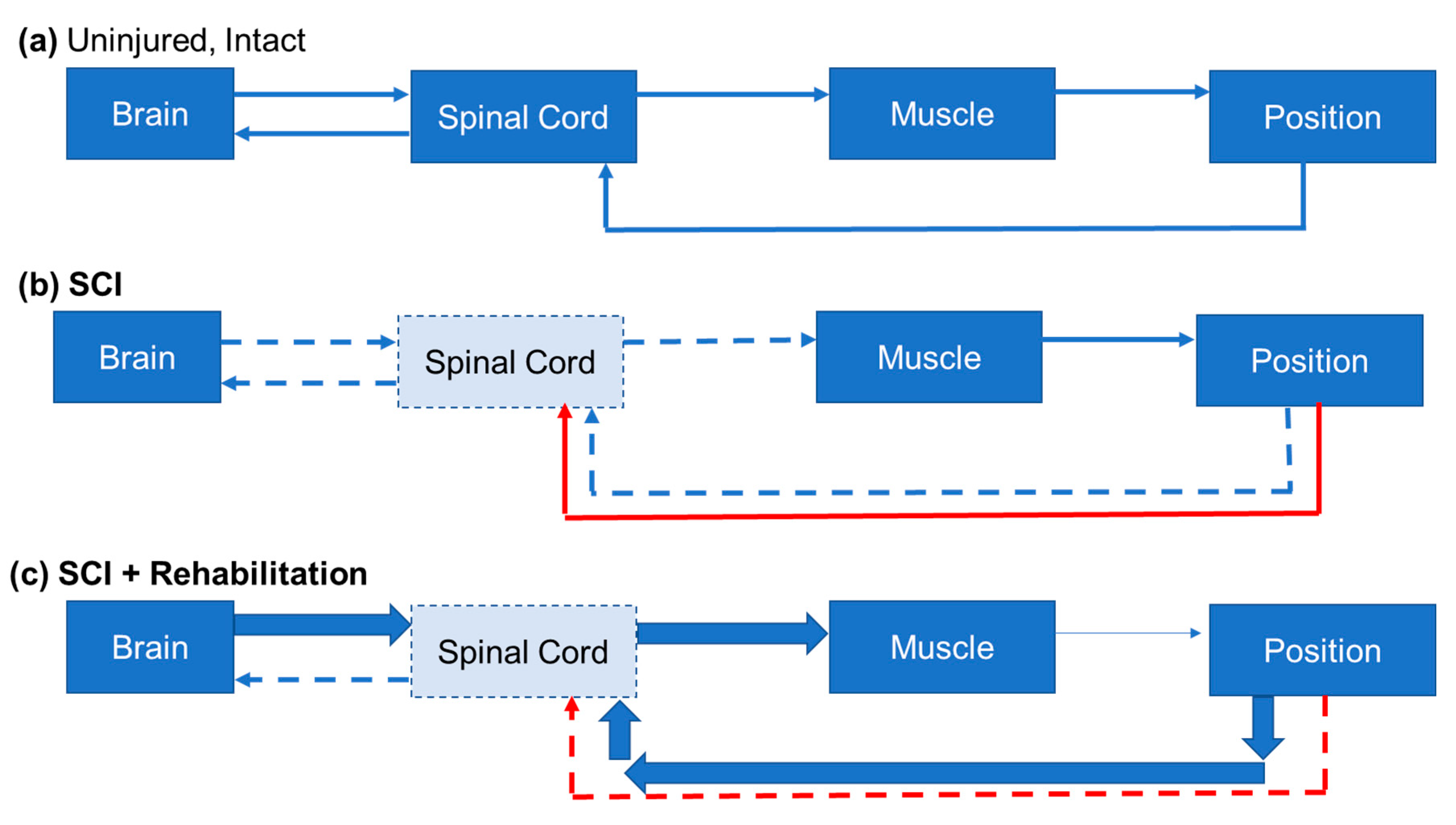
3. Neural Plasticity Associated with Reaching and Grasping after SCI
3.1. Plasticity in Spared Descending Systems
3.2. Neuromodulation to Drive Descending Plasticity after SCI
3.3. Primary Afferent Plasticity
4. Rehabilitation, and Afferent Driven Plasticity for Reaching and Grasping
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Donnelly, D.J.; Popovich, P.G. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Detloff, M.R.; Fisher, L.C.; McGaughy, V.; Longbrake, E.E.; Popovich, P.G.; Basso, D.M. Remote activation of microglia and pro-inflammatory cytokines predict the onset and severity of below-level neuropathic pain after spinal cord injury in rats. Exp. Neurol. 2008, 212, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Gensel, J.C.; Zhang, B. Macrophage activation and its role in repair and pathology after spinal cord injury. Brain Res. 2015, 1619, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kopper, T.J.; Gensel, J.C. Myelin as an inflammatory mediator: Myelin interactions with complement, macrophages, and microglia in spinal cord injury. J. Neurosci. Res. 2018, 96, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Chhaya, S.J.; Quiros-Molina, D.; Tamashiro-Orrego, A.D.; Houle, J.D.; Detloff, M.R. Exercise-Induced Changes to the Macrophage Response in the Dorsal Root Ganglia Prevent Neuropathic Pain after Spinal Cord Injury. J. Neurotraum. 2019, 36, 877–890. [Google Scholar] [CrossRef]
- Kroner, A.; Almanza, J.R. Role of microglia in spinal cord injury. Neurosci. Lett. 2019, 709, 134370. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, T.M.; Burda, J.E.; Sofroniew, M.V. Cell biology of spinal cord injury and repair. J. Clin. Investig. 2017, 127, 3259–3270. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.P.; Warren, P.M.; Silver, J. New insights into glial scar formation after spinal cord injury. Cell Tissue Res. 2021. [Google Scholar] [CrossRef]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Reier, P.J.; Houle, J.D. The glial scar: Its bearing on axonal elongation and transplantation approaches to CNS repair. Adv. Neurol. 1988, 47, 87–138. [Google Scholar] [PubMed]
- Snow, D.M.; Brown, E.M.; Letourneau, P.C. Growth cone behavior in the presence of soluble chondroitin sulfate proteoglycan (CSPG), compared to behavior on CSPG bound to laminin or fibronectin. Int. J. Dev. Neurosci. 1996, 14, 331–349. [Google Scholar] [CrossRef]
- Fawcett, J.W.; Asher, R.A. The glial scar and central nervous system repair. Brain Res. Bull. 1999, 49, 377–391. [Google Scholar] [CrossRef]
- Herrmann, J.E.; Imura, T.; Song, B.; Qi, J.; Ao, Y.; Nguyen, T.K.; Korsak, R.A.; Takeda, K.; Akira, S.; Sofroniew, M.V. STAT3 is a critical regulator of astrogliosis and scar formation after spinal cord injury. J. Neurosci. 2008, 28, 7231–7243. [Google Scholar] [CrossRef]
- Tom, V.J.; Houle, J.D. Intraspinal microinjection of chondroitinase ABC following injury promotes axonal regeneration out of a peripheral nerve graft bridge. Exp. Neurol. 2008, 211, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, A.P.; Warren, P.M.; Silver, J. The Biology of Regeneration Failure and Success After Spinal Cord Injury. Physiol. Rev. 2018, 98, 881–917. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, M.L.; Tom, V.J. Neuroimmune System as a Driving Force for Plasticity Following CNS Injury. Front. Cell Neurosci. 2020, 14, 187. [Google Scholar] [CrossRef]
- Anderson, M.A.; Burda, J.E.; Ren, Y.; Ao, Y.; O’Shea, T.M.; Kawaguchi, R.; Coppola, G.; Khakh, B.S.; Deming, T.J.; Sofroniew, M.V. Astrocyte scar formation aids central nervous system axon regeneration. Nature 2016, 532, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Geoffroy, C.G.; Chan, A.F.; Tolentino, K.E.; Crawford, M.J.; Leal, M.A.; Kang, B.; Zheng, B. Assessing spinal axon regeneration and sprouting in Nogo-, MAG-, and OMgp-deficient mice. Neuron 2010, 66, 663–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Spinal Cord Injury Statistical Center. Facts and Figures at a Glance; University of Alabama at Birmingham: Birmingham, AL, USA, 2021. [Google Scholar]
- Xiao, Z.; Tang, F.; Zhao, Y.; Han, G.; Yin, N.; Li, X.; Chen, B.; Han, S.; Jiang, X.; Yun, C.; et al. Significant Improvement of Acute Complete Spinal Cord Injury Patients Diagnosed by a Combined Criteria Implanted with NeuroRegen Scaffolds and Mesenchymal Stem Cells. Cell Transplant. 2018, 27, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Flesher, S.N.; Downey, J.E.; Weiss, J.M.; Hughes, C.L.; Herrera, A.J.; Tyler-Kabara, E.C.; Boninger, M.L.; Collinger, J.L.; Gaunt, R.A. A brain-computer interface that evokes tactile sensations improves robotic arm control. Science 2021, 372, 831–836. [Google Scholar] [CrossRef]
- Goshgarian, H.G. Developmental plasticity in the respiratory pathway of the adult rat. Exp. Neurol. 1979, 66, 547–555. [Google Scholar] [CrossRef]
- Zimmer, M.B.; Goshgarian, H.G. GABA, not glycine, mediates inhibition of latent respiratory motor pathways after spinal cord injury. Exp. Neurol. 2007, 203, 493–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.Z.; Dougherty, B.J.; Sandhu, M.S.; Lane, M.A.; Reier, P.J.; Fuller, D.D. Phrenic motoneuron discharge patterns following chronic cervical spinal cord injury. Exp. Neurol. 2013, 249, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hachem, L.D.; Ahuja, C.S.; Fehlings, M.G. Assessment and management of acute spinal cord injury: From point of injury to rehabilitation. J. Spinal Cord Med. 2017, 40, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Wolpaw, J.R.; Lee, C.L.; Carp, J.S. Operantly conditioned plasticity in spinal cord. Ann. N. Y. Acad. Sci. 1991, 627, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Osman, J.; Field-Fote, E.C. Cortical vs. afferent stimulation as an adjunct to functional task practice training: A randomized, comparative pilot study in people with cervical spinal cord injury. Clin. Rehabil. 2015, 29, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Field-Fote, E.C. Exciting recovery: Augmenting practice with stimulation to optimize outcomes after spinal cord injury. Prog. Brain Res. 2015, 218, 103–126. [Google Scholar] [CrossRef] [PubMed]
- Schildt, C.J.; Thomas, S.H.; Powell, E.S.; Sawaki, L.; Sunderam, S. Closed-loop afferent electrical stimulation for recovery of hand function in individuals with motor incomplete spinal injury: Early clinical results. In Proceedings of the 2016 38th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Orlando, FL, USA, 16–20 August 2016; pp. 1552–1555. [Google Scholar] [CrossRef] [PubMed]
- Fee, M.S. The role of efference copy in striatal learning. Curr. Opin. Neurobiol. 2014, 25, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Alstermark, B.; Ekerot, C.F. The lateral reticular nucleus; integration of descending and ascending systems regulating voluntary forelimb movements. Front. Comput. Neurosci. 2015, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Person, A.L. Corollary Discharge Signals in the Cerebellum. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2019, 4, 813–819. [Google Scholar] [CrossRef]
- Van Kemenade, B.M.; Arikan, B.E.; Podranski, K.; Steinstrater, O.; Kircher, T.; Straube, B. Distinct Roles for the Cerebellum, Angular Gyrus, and Middle Temporal Gyrus in Action-Feedback Monitoring. Cereb. Cortex 2019, 29, 1520–1531. [Google Scholar] [CrossRef]
- Kandel, E.; Koster, J.; Mack, S.; Siegelbaum, S. Principles of Neural Science, 5th ed.; McGraw-Hill: New York, NY, USA, 2013. [Google Scholar]
- Sweatt, J.D. Neural plasticity and behavior—Sixty years of conceptual advances. J. Neurochem. 2016, 139, 179–199. [Google Scholar] [CrossRef] [PubMed]
- Bliss, T.V.; Lomo, T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 1973, 232, 331–356. [Google Scholar] [CrossRef] [PubMed]
- Hebb, D.O. The Organization of Behavior: A Neuropsychological Theory; John Wiley & Sons: New York, NY, USA, 1949. [Google Scholar]
- Zenke, F.; Gerstner, W. Hebbian plasticity requires compensatory processes on multiple timescales. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diering, G.H.; Huganir, R.L. The AMPA Receptor Code of Synaptic Plasticity. Neuron 2018, 100, 314–329. [Google Scholar] [CrossRef] [Green Version]
- Dittman, J.S.; Ryan, T.A. The control of release probability at nerve terminals. Nat. Rev. Neurosci. 2019, 20, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.M. Structural LTP: From synaptogenesis to regulated synapse enlargement and clustering. Curr. Opin. Neurobiol. 2020, 63, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Zucker, R.S.; Regehr, W.G. Short-term synaptic plasticity. Annu. Rev. Physiol. 2002, 64, 355–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackman, S.L.; Regehr, W.G. The Mechanisms and Functions of Synaptic Facilitation. Neuron 2017, 94, 447–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luscher, C.; Malenka, R.C. NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb. Perspect. Biol. 2012, 4, a005710. [Google Scholar] [CrossRef] [Green Version]
- Koike-Tani, M.; Kanda, T.; Saitoh, N.; Yamashita, T.; Takahashi, T. Involvement of AMPA receptor desensitization in short-term synaptic depression at the calyx of Held in developing rats. J. Physiol. 2008, 586, 2263–2275. [Google Scholar] [CrossRef] [PubMed]
- Herring, B.E.; Nicoll, R.A. Long-Term Potentiation: From CaMKII to AMPA Receptor Trafficking. Annu. Rev. Physiol. 2016, 78, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, G.; Hansel, C. Cerebellar long-term potentiation: Cellular mechanisms and role in learning. Int. Rev. Neurobiol. 2014, 117, 39–51. [Google Scholar] [CrossRef]
- Wolpaw, J.R. Spinal cord plasticity in acquisition and maintenance of motor skills. Acta Physiol. 2007, 189, 155–169. [Google Scholar] [CrossRef]
- Roth, R.H.; Cudmore, R.H.; Tan, H.L.; Hong, I.; Zhang, Y.; Huganir, R.L. Cortical Synaptic AMPA Receptor Plasticity during Motor Learning. Neuron 2020, 105, 895–908.e5. [Google Scholar] [CrossRef]
- Adkins, D.L.; Boychuk, J.; Remple, M.S.; Kleim, J.A. Motor training induces experience-specific patterns of plasticity across motor cortex and spinal cord. J. Appl. Physiol. 2006, 101, 1776–1782. [Google Scholar] [CrossRef] [Green Version]
- Hoy, K.C.; Huie, J.R.; Grau, J.W. AMPA receptor mediated behavioral plasticity in the isolated rat spinal cord. Behav. Brain Res. 2013, 236, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Ganzer, P.D.; Beringer, C.R.; Shumsky, J.S.; Nwaobasi, C.; Moxon, K.A. Serotonin receptor and dendritic plasticity in the spinal cord mediated by chronic serotonergic pharmacotherapy combined with exercise following complete SCI in the adult rat. Exp. Neurol. 2018, 304, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J. Glutamatergic synapses are structurally and biochemically complex because of multiple plasticity processes: Long-term potentiation, long-term depression, short-term potentiation and scaling. Philos. Trans. R. Soc. Lond. B Biol. Sci 2017, 372, 20160260. [Google Scholar] [CrossRef]
- Purkey, A.M.; Dell’Acqua, M.L. Phosphorylation-Dependent Regulation of Ca(2+)-Permeable AMPA Receptors During Hippocampal Synaptic Plasticity. Front. Synaptic. Neurosci. 2020, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Kulik, Y.D.; Watson, D.J.; Cao, G.; Kuwajima, M.; Harris, K.M. Structural plasticity of dendritic secretory compartments during LTP-induced synaptogenesis. Elife 2019, 8, e46356. [Google Scholar] [CrossRef]
- Whishaw, I.Q.; Pellis, S.M. The structure of skilled forelimb reaching in the rat: A proximally driven movement with a single distal rotatory component. Behav. Brain Res. 1990, 41, 49–59. [Google Scholar] [CrossRef]
- Whishaw, I.Q.; Pellis, S.M.; Gorny, B.; Kolb, B.; Tetzlaff, W. Proximal and distal impairments in rat forelimb use in reaching follow unilateral pyramidal tract lesions. Behav. Brain Res. 1993, 56, 59–76. [Google Scholar] [CrossRef]
- Haines, D.E. Neuroanatomy: An Atlas of Structures, Sections, and Systems; Lippincott Williams and Wilkons: Philadelphia, PA, USA, 2000; Volume 5. [Google Scholar]
- Pizzimenti, M.A.; Darling, W.G.; Rotella, D.L.; McNeal, D.W.; Herrick, J.L.; Ge, J.; Stilwell-Morecraft, K.S.; Morecraft, R.J. Measurement of reaching kinematics and prehensile dexterity in nonhuman primates. J. Neurophysiol. 2007, 98, 1015–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, H.X.; Gharbawie, O.A.; Wynne, K.W.; Kaas, J.H. Impairment and recovery of hand use after unilateral section of the dorsal columns of the spinal cord in squirrel monkeys. Behav. Brain Res. 2013, 252, 363–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geed, S.; McCurdy, M.L.; van Kan, P.L. Neuronal Correlates of Functional Coupling between Reach- and Grasp-Related Components of Muscle Activity. Front. Neural. Circuits 2017, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, S.N. The primate reticulospinal tract, hand function and functional recovery. J. Physiol. 2011, 589, 5603–5612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathyamurthy, A.; Barik, A.; Dobrott, C.I.; Matson, K.J.E.; Stoica, S.; Pursley, R.; Chesler, A.T.; Levine, A.J. Cerebellospinal Neurons Regulate Motor Performance and Motor Learning. Cell Rep. 2020, 31, 107595. [Google Scholar] [CrossRef] [PubMed]
- McKenna, J.E.; Prusky, G.T.; Whishaw, I.Q. Cervical motoneuron topography reflects the proximodistal organization of muscles and movements of the rat forelimb: A retrograde carbocyanine dye analysis. J. Comp. Neurol. 2000, 419, 286–296. [Google Scholar] [CrossRef]
- Klein, A.; Sacrey, L.A.; Dunnett, S.B.; Whishaw, I.Q.; Nikkhah, G. Proximal movements compensate for distal forelimb movement impairments in a reach-to-eat task in Huntington’s disease: New insights into motor impairments in a real-world skill. Neurobiol. Dis. 2011, 41, 560–569. [Google Scholar] [CrossRef]
- Klein, A.; Sacrey, L.A.; Whishaw, I.Q.; Dunnett, S.B. The use of rodent skilled reaching as a translational model for investigating brain damage and disease. Neurosci. Biobehav. Rev. 2012, 36, 1030–1042. [Google Scholar] [CrossRef]
- Krisa, L.; Runyen, M.; Detloff, M.R. Translational Challenges of Rat Models of Upper Extremity Dysfunction After Spinal Cord Injury. Top. Spinal Cord Inj. Rehabil. 2018, 24, 195–205. [Google Scholar] [CrossRef]
- Gallegos, C.; Carey, M.; Zheng, Y.; He, X.; Cao, Q.L. Reaching and Grasping Training Improves Functional Recovery After Chronic Cervical Spinal Cord Injury. Front. Cell Neurosci. 2020, 14, 110. [Google Scholar] [CrossRef]
- Montoya, C.P.; Campbell-Hope, L.J.; Pemberton, K.D.; Dunnett, S.B. The “staircase test”: A measure of independent forelimb reaching and grasping abilities in rats. J. Neurosci. Methods 1991, 36, 219–228. [Google Scholar] [CrossRef]
- Sindhurakar, A.; Butensky, S.D.; Meyers, E.; Santos, J.; Bethea, T.; Khalili, A.; Sloan, A.P.; Rennaker, R.L., 3rd; Carmel, J.B. An Automated Test of Rat Forelimb Supination Quantifies Motor Function Loss and Recovery After Corticospinal Injury. Neurorehabil. Neural. Repair. 2017, 31, 122–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvine, K.A.; Ferguson, A.R.; Mitchell, K.D.; Beattie, S.B.; Beattie, M.S.; Bresnahan, J.C. A novel method for assessing proximal and distal forelimb function in the rat: The Irvine, Beatties and Bresnahan (IBB) forelimb scale. J. Vis. Exp. 2010, 46, e2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvine, K.A.; Ferguson, A.R.; Mitchell, K.D.; Beattie, S.B.; Lin, A.; Stuck, E.D.; Huie, J.R.; Nielson, J.L.; Talbott, J.F.; Inoue, T.; et al. The Irvine, Beatties, and Bresnahan (IBB) Forelimb Recovery Scale: An Assessment of Reliability and Validity. Front. Neurol. 2014, 5, 116. [Google Scholar] [CrossRef] [PubMed]
- Ballermann, M.; Metz, G.A.; McKenna, J.E.; Klassen, F.; Whishaw, I.Q. The pasta matrix reaching task: A simple test for measuring skilled reaching distance, direction, and dexterity in rats. J. Neurosci. Methods 2001, 106, 39–45. [Google Scholar] [CrossRef]
- Bertelli, J.A.; Mira, J.C. Behavioral evaluating methods in the objective clinical assessment of motor function after experimental brachial plexus reconstruction in the rat. J. Neurosci. Methods 1993, 46, 203–208. [Google Scholar] [CrossRef]
- Anderson, K.D.; Gunawan, A.; Steward, O. Quantitative assessment of forelimb motor function after cervical spinal cord injury in rats: Relationship to the corticospinal tract. Exp. Neurol. 2005, 194, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Khaing, Z.Z.; Geissler, S.A.; Jiang, S.; Milman, B.D.; Aguilar, S.V.; Schmidt, C.E.; Schallert, T. Assessing forelimb function after unilateral cervical spinal cord injury: Novel forelimb tasks predict lesion severity and recovery. J. Neurotraum. 2012, 29, 488–498. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.M.; Fisher, K.M.; Ahloy-Dallaire, J.; Darian-Smith, C. Somatosensory corticospinal tract axons sprout within the cervical cord following a dorsal root/dorsal column spinal injury in the rat. J. Comp. Neurol. 2020, 528, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Rosario, A.; Berrios-Torres, I.; Adeli, H.; Buford, J.A. Combined corticospinal and reticulospinal effects on upper limb muscles. Neurosci. Lett. 2014, 561, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.N.; Zaaimi, B.; Fisher, K.M.; Edgley, S.A.; Soteropoulos, D.S. Pathways mediating functional recovery. Prog. Brain Res. 2015, 218, 389–412. [Google Scholar] [CrossRef]
- Palmer, E.; Ashby, P. Corticospinal projections to upper limb motoneurones in humans. J. Physiol. 1992, 448, 397–412. [Google Scholar] [CrossRef] [Green Version]
- De Noordhout, A.M.; Rapisarda, G.; Bogacz, D.; Gerard, P.; De Pasqua, V.; Pennisi, G.; Delwaide, P.J. Corticomotoneuronal synaptic connections in normal man: An electrophysiological study. Brain 1999, 122, 1327–1340. [Google Scholar] [CrossRef] [Green Version]
- Tazoe, T.; Perez, M.A. Cortical and reticular contributions to human precision and power grip. J. Physiol. 2017, 595, 2715–2730. [Google Scholar] [CrossRef] [Green Version]
- Schrimsher, G.W.; Reier, P.J. Forelimb motor performance following dorsal column, dorsolateral funiculi, or ventrolateral funiculi lesions of the cervical spinal cord in the rat. Exp. Neurol. 1993, 120, 264–276. [Google Scholar] [CrossRef]
- Morecraft, R.J.; Ge, J.; Stilwell-Morecraft, K.S.; McNeal, D.W.; Pizzimenti, M.A.; Darling, W.G. Terminal distribution of the corticospinal projection from the hand/arm region of the primary motor cortex to the cervical enlargement in rhesus monkey. J. Comp. Neurol. 2013, 521, 4205–4235. [Google Scholar] [CrossRef] [Green Version]
- Oudega, M.; Perez, M.A. Corticospinal reorganization after spinal cord injury. J. Physiol. 2012, 590, 3647–3663. [Google Scholar] [CrossRef] [PubMed]
- Weidner, N.; Ner, A.; Salimi, N.; Tuszynski, M.H. Spontaneous corticospinal axonal plasticity and functional recovery after adult central nervous system injury. Proc. Natl. Acad. Sci. USA 2001, 98, 3513–3518. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Ninomiya, T.; Yamashita, T.; Takada, M. Reorganization of corticospinal tract fibers after spinal cord injury in adult macaques. Sci. Rep. 2015, 5, 11986. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, D.M. Supraspinal contributions to the initiation and control of locomotion in the cat. Prog. Neurobiol. 1986, 26, 273–361. [Google Scholar] [CrossRef]
- Shik, M.L.; Orlovsky, G.N. Neurophysiology of locomotor automatism. Physiol. Rev. 1976, 56, 465–501. [Google Scholar] [CrossRef]
- Dougherty, K.J.; Kiehn, O. Functional organization of V2a-related locomotor circuits in the rodent spinal cord. Ann. N. Y. Acad. Sci. 2010, 1198, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Alaynick, W.A.; Jessell, T.M.; Pfaff, S.L. SnapShot: Spinal cord development. Cell 2011, 146, 178–178.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.C.; Niu, T.; Alaynick, W.A. Molecular and cellular development of spinal cord locomotor circuitry. Front. Mol. Neurosci. 2015, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Rybak, I.A.; Dougherty, K.J.; Shevtsova, N.A. Organization of the Mammalian Locomotor CPG: Review of Computational Model and Circuit Architectures Based on Genetically Identified Spinal Interneurons(1,2,3). eNeuro 2015, 2, 1–20. [Google Scholar] [CrossRef]
- Flynn, J.R.; Conn, V.L.; Boyle, K.A.; Hughes, D.I.; Watanabe, M.; Velasquez, T.; Goulding, M.D.; Callister, R.J.; Graham, B.A. Anatomical and Molecular Properties of Long Descending Propriospinal Neurons in Mice. Front. Neuroanat. 2017, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zholudeva, L.V.; Qiang, L.; Marchenko, V.; Dougherty, K.J.; Sakiyama-Elbert, S.E.; Lane, M.A. The Neuroplastic and Therapeutic Potential of Spinal Interneurons in the Injured Spinal Cord. Trends Neurosci. 2018, 41, 625–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrott, C.I.; Sathyamurthy, A.; Levine, A.J. Decoding Cell Type Diversity Within the Spinal Cord. Curr. Opin. Physiol. 2019, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zholudeva, L.V.; Abraira, V.E.; Satkunendrarajah, K.; McDevitt, T.C.; Goulding, M.D.; Magnuson, D.S.K.; Lane, M.A. Spinal Interneurons as Gatekeepers to Neuroplasticity after Injury or Disease. J. Neurosci. 2021, 41, 845–854. [Google Scholar] [CrossRef]
- Abraira, V.E.; Kuehn, E.D.; Chirila, A.M.; Springel, M.W.; Toliver, A.A.; Zimmerman, A.L.; Orefice, L.L.; Boyle, K.A.; Bai, L.; Song, B.J.; et al. The Cellular and Synaptic Architecture of the Mechanosensory Dorsal Horn. Cell 2017, 168, 295–310.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatto, G.; Bourane, S.; Ren, X.; Di Costanzo, S.; Fenton, P.K.; Halder, P.; Seal, R.P.; Goulding, M.D. A Functional Topographic Map for Spinal Sensorimotor Reflexes. Neuron 2021, 109, 91–104.e5. [Google Scholar] [CrossRef]
- Pocratsky, A.M.; Shepard, C.T.; Morehouse, J.R.; Burke, D.A.; Riegler, A.S.; Hardin, J.T.; Beare, J.E.; Hainline, C.; States, G.J.; Brown, B.L.; et al. Long ascending propriospinal neurons provide flexible, context-specific control of interlimb coordination. Elife 2020, 9, e53565. [Google Scholar] [CrossRef] [PubMed]
- Bareyre, F.M.; Kerschensteiner, M.; Raineteau, O.; Mettenleiter, T.C.; Weinmann, O.; Schwab, M.E. The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat. Neurosci. 2004, 7, 269–277. [Google Scholar] [CrossRef]
- Raineteau, O.; Schwab, M.E. Plasticity of motor systems after incomplete spinal cord injury. Nat. Rev. Neurosci. 2001, 2, 263–273. [Google Scholar] [CrossRef]
- Courtine, G.; Song, B.; Roy, R.R.; Zhong, H.; Herrmann, J.E.; Ao, Y.; Qi, J.; Edgerton, V.R.; Sofroniew, M.V. Recovery of supraspinal control of stepping via indirect propriospinal relay connections after spinal cord injury. Nat. Med. 2008, 14, 69–74. [Google Scholar] [CrossRef]
- Benthall, K.N.; Hough, R.A.; McClellan, A.D. Descending propriospinal neurons mediate restoration of locomotor function following spinal cord injury. J. Neurophysiol. 2017, 117, 215–229. [Google Scholar] [CrossRef]
- Morris, R.; Tosolini, A.P.; Goldstein, J.D.; Whishaw, I.Q. Impaired arpeggio movement in skilled reaching by rubrospinal tract lesions in the rat: A behavioral/anatomical fractionation. J. Neurotraum. 2011, 28, 2439–2451. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Hinckley, C.A.; Driscoll, S.P.; Moore, N.J.; Levine, A.J.; Hilde, K.L.; Sharma, K.; Pfaff, S.L. Graded Arrays of Spinal and Supraspinal V2a Interneuron Subtypes Underlie Forelimb and Hindlimb Motor Control. Neuron 2018, 97, 869–884.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whishaw, I.Q.; Gorny, B. Arpeggio and fractionated digit movements used in prehension by rats. Behav. Brain Res. 1994, 60, 15–24. [Google Scholar] [CrossRef]
- Isa, T.; Kinoshita, M.; Nishimura, Y. Role of Direct vs. Indirect Pathways from the Motor Cortex to Spinal Motoneurons in the Control of Hand Dexterity. Front. Neurol. 2013, 4, 191. [Google Scholar] [CrossRef] [Green Version]
- Azim, E.; Jiang, J.; Alstermark, B.; Jessell, T.M. Skilled reaching relies on a V2a propriospinal internal copy circuit. Nature 2014, 508, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Brouwer, B.; Hopkins-Rosseel, D.H. Motor cortical mapping of proximal upper extremity muscles following spinal cord injury. Spinal Cord 1997, 35, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Calancie, B.; Alexeeva, N.; Broton, J.G.; Suys, S.; Hall, A.; Klose, K.J. Distribution and latency of muscle responses to transcranial magnetic stimulation of motor cortex after spinal cord injury in humans. J. Neurotraum. 1999, 16, 49–67. [Google Scholar] [CrossRef]
- McPherson, J.G.; Chen, A.; Ellis, M.D.; Yao, J.; Heckman, C.J.; Dewald, J.P.A. Progressive recruitment of contralesional cortico-reticulospinal pathways drives motor impairment post stroke. J. Physiol. 2018, 596, 1211–1225. [Google Scholar] [CrossRef]
- Sangari, S.; Perez, M.A. Distinct Corticospinal and Reticulospinal Contributions to Voluntary Control of Elbow Flexor and Extensor Muscles in Humans with Tetraplegia. J. Neurosci. 2020, 40, 8831–8841. [Google Scholar] [CrossRef]
- Ditunno, J.F., Jr.; Stover, S.L.; Freed, M.M.; Ahn, J.H. Motor recovery of the upper extremities in traumatic quadriplegia: A multicenter study. Arch. Phys. Med. Rehabil. 1992, 73, 431–436. [Google Scholar]
- Ditunno, J.F., Jr.; Cohen, M.E.; Hauck, W.W.; Jackson, A.B.; Sipski, M.L. Recovery of upper-extremity strength in complete and incomplete tetraplegia: A multicenter study. Arch. Phys. Med. Rehabil. 2000, 81, 389–393. [Google Scholar] [CrossRef]
- Mateo, S.; Di Marco, J.; Cucherat, M.; Gueyffier, F.; Rode, G. Inconclusive efficacy of intervention on upper-limb function after tetraplegia: A systematic review and meta-analysis. Ann. Phys. Rehabil. Med. 2020, 63, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Kasten, M.R.; Sunshine, M.D.; Secrist, E.S.; Horner, P.J.; Moritz, C.T. Therapeutic intraspinal microstimulation improves forelimb function after cervical contusion injury. J. Neural. Eng. 2013, 10, 044001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajiboye, A.B.; Willett, F.R.; Young, D.R.; Memberg, W.D.; Murphy, B.A.; Miller, J.P.; Walter, B.L.; Sweet, J.A.; Hoyen, H.A.; Keith, M.W.; et al. Restoration of reaching and grasping movements through brain-controlled muscle stimulation in a person with tetraplegia: A proof-of-concept demonstration. Lancet 2017, 389, 1821–1830. [Google Scholar] [CrossRef] [Green Version]
- Mateo, S.; Roby-Brami, A.; Reilly, K.T.; Rossetti, Y.; Collet, C.; Rode, G. Upper limb kinematics after cervical spinal cord injury: A review. J. Neuroeng. Rehabil. 2015, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.F.; Wang, R.H.; Zariffa, J. Generalizability of Hand-Object Interaction Detection in Egocentric Video across Populations with Hand Impairment. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2020, 2020, 3228–3231. [Google Scholar] [CrossRef]
- Asboth, L.; Friedli, L.; Beauparlant, J.; Martinez-Gonzalez, C.; Anil, S.; Rey, E.; Baud, L.; Pidpruzhnykova, G.; Anderson, M.A.; Shkorbatova, P.; et al. Cortico-reticulo-spinal circuit reorganization enables functional recovery after severe spinal cord contusion. Nat. Neurosci. 2018, 21, 576–588. [Google Scholar] [CrossRef]
- Davidson, A.G.; Buford, J.A. Bilateral actions of the reticulospinal tract on arm and shoulder muscles in the monkey: Stimulus triggered averaging. Exp. Brain Res. 2006, 173, 25–39. [Google Scholar] [CrossRef]
- Davidson, A.G.; Schieber, M.H.; Buford, J.A. Bilateral spike-triggered average effects in arm and shoulder muscles from the monkey pontomedullary reticular formation. J. Neurosci. 2007, 27, 8053–8058. [Google Scholar] [CrossRef]
- Herbert, W.J.; Davidson, A.G.; Buford, J.A. Measuring the motor output of the pontomedullary reticular formation in the monkey: Do stimulus-triggered averaging and stimulus trains produce comparable results in the upper limbs? Exp. Brain Res. 2010, 203, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Filli, L.; Engmann, A.K.; Zorner, B.; Weinmann, O.; Moraitis, T.; Gullo, M.; Kasper, H.; Schneider, R.; Schwab, M.E. Bridging the gap: A reticulo-propriospinal detour bypassing an incomplete spinal cord injury. J. Neurosci. 2014, 34, 13399–13410. [Google Scholar] [CrossRef] [Green Version]
- Zorner, B.; Bachmann, L.C.; Filli, L.; Kapitza, S.; Gullo, M.; Bolliger, M.; Starkey, M.L.; Rothlisberger, M.; Gonzenbach, R.R.; Schwab, M.E. Chasing central nervous system plasticity: The brainstem’s contribution to locomotor recovery in rats with spinal cord injury. Brain 2014, 137, 1716–1732. [Google Scholar] [CrossRef] [Green Version]
- May, Z.; Fenrich, K.K.; Dahlby, J.; Batty, N.J.; Torres-Espin, A.; Fouad, K. Following Spinal Cord Injury Transected Reticulospinal Tract Axons Develop New Collateral Inputs to Spinal Interneurons in Parallel with Locomotor Recovery. Neural. Plast 2017, 2017, 1932875. [Google Scholar] [CrossRef] [PubMed]
- Taub, E.; Wolf, S.L. Constraint Induced Movement Techniques To Facilitate Upper Extremity Use in Stroke Patients. Top. Stroke Rehabil. 1997, 3, 38–61. [Google Scholar] [CrossRef] [PubMed]
- Dos Anjos, S.; Morris, D.; Taub, E. Constraint-Induced Movement Therapy for Lower Extremity Function: Describing the LE-CIMT Protocol. Phys. Ther. 2020, 100, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Popp, W.L.; Brogioli, M.; Albisser, U.; Ortmann, S.; Velstra, I.M.; Demko, L.; Gassert, R.; Curt, A. Predicting upper limb compensation during prehension tasks in tetraplegic spinal cord injured patients using a single wearable sensor. IEEE Int. Conf. Rehabil. Robot. 2019, 2019, 1000–1006. [Google Scholar] [CrossRef]
- Bockbrader, M.A.; Francisco, G.; Lee, R.; Olson, J.; Solinsky, R.; Boninger, M.L. Brain Computer Interfaces in Rehabilitation Medicine. PM R 2018, 10, S233–S243. [Google Scholar] [CrossRef] [Green Version]
- Jorge, A.; Royston, D.A.; Tyler-Kabara, E.C.; Boninger, M.L.; Collinger, J.L. Classification of Individual Finger Movements Using Intracortical Recordings in Human Motor Cortex. Neurosurgery 2020, 87, 630–638. [Google Scholar] [CrossRef]
- Zimmermann, J.B.; Jackson, A. Closed-loop control of spinal cord stimulation to restore hand function after paralysis. Front. Neurosci. 2014, 8, 87. [Google Scholar] [CrossRef]
- Benavides, F.D.; Jo, H.J.; Lundell, H.; Edgerton, V.R.; Gerasimenko, Y.; Perez, M.A. Cortical and Subcortical Effects of Transcutaneous Spinal Cord Stimulation in Humans with Tetraplegia. J. Neurosci. 2020, 40, 2633–2643. [Google Scholar] [CrossRef]
- Anderson, K.D.; Bryden, A.M.; Moynahan, M. Risk-benefit value of upper extremity function by an implanted electrical stimulation device targeting chronic cervical spinal cord injury. Spinal. Cord. Ser. Cases 2019, 5, 68. [Google Scholar] [CrossRef]
- Ganzer, P.D.; Colachis, S.C.t.; Schwemmer, M.A.; Friedenberg, D.A.; Dunlap, C.F.; Swiftney, C.E.; Jacobowitz, A.F.; Weber, D.J.; Bockbrader, M.A.; Sharma, G. Restoring the Sense of Touch Using a Sensorimotor Demultiplexing Neural Interface. Cell 2020, 181, 763–773.e712. [Google Scholar] [CrossRef] [PubMed]
- Geed, S.; van Kan, P.L.E. Grasp-Based Functional Coupling Between Reach- and Grasp-Related Components of Forelimb Muscle Activity. J. Mot. Behav. 2017, 49, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Gatto, G.; Smith, K.M.; Ross, S.E.; Goulding, M. Neuronal diversity in the somatosensory system: Bridging the gap between cell type and function. Curr. Opin. Neurobiol. 2019, 56, 167–174. [Google Scholar] [CrossRef]
- Abraira, V.E.; Ginty, D.D. The sensory neurons of touch. Neuron 2013, 79, 618–639. [Google Scholar] [CrossRef] [Green Version]
- Fink, A.J.; Croce, K.R.; Huang, Z.J.; Abbott, L.F.; Jessell, T.M.; Azim, E. Presynaptic inhibition of spinal sensory feedback ensures smooth movement. Nature 2014, 509, 43–48. [Google Scholar] [CrossRef]
- Koch, S.C.; Del Barrio, M.G.; Dalet, A.; Gatto, G.; Gunther, T.; Zhang, J.; Seidler, B.; Saur, D.; Schule, R.; Goulding, M. RORbeta Spinal Interneurons Gate Sensory Transmission during Locomotion to Secure a Fluid Walking Gait. Neuron 2017, 96, 1419–1431.e5. [Google Scholar] [CrossRef] [Green Version]
- Knikou, M.; Chaudhuri, D.; Kay, E.; Schmit, B.D. Pre- and post-alpha motoneuronal control of the soleus H-reflex during sinusoidal hip movements in human spinal cord injury. Brain Res. 2006, 1103, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.N.; Chambers, W.W. Intraspinal sprouting of dorsal root axons; development of new collaterals and preterminals following partial denervation of the spinal cord in the cat. AMA Arch. Neurol. Psychiatry 1958, 79, 46–61. [Google Scholar] [CrossRef]
- Goldberger, M.E.; Murray, M. Restitution of function and collateral sprouting in the cat spinal cord: The deafferented animal. J. Comp. Neurol. 1974, 158, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Goldberger, M.E. Restitution of function and collateral sprouting in the cat spinal cord: The partially hemisected animal. J. Comp. Neurol. 1974, 158, 19–36. [Google Scholar] [CrossRef]
- Polistina, D.C.; Murray, M.; Goldberger, M.E. Plasticity of dorsal root and descending serotoninergic projections after partial deafferentation of the adult rat spinal cord. J. Comp. Neurol. 1990, 299, 349–363. [Google Scholar] [CrossRef]
- Rank, M.M.; Galea, M.P.; Callister, R.; Callister, R.J. Is more always better? How different ’doses’ of exercise after incomplete spinal cord injury affects the membrane properties of deep dorsal horn interneurons. Exp. Neurol. 2018, 300, 201–211. [Google Scholar] [CrossRef]
- Peirs, C.; Dallel, R.; Todd, A.J. Recent advances in our understanding of the organization of dorsal horn neuron populations and their contribution to cutaneous mechanical allodynia. J. Neural. Transm. 2020, 127, 505–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurin, J.; Pertici, V.; Dousset, E.; Marqueste, T.; Decherchi, P. Group III and IV muscle afferents: Role on central motor drive and clinical implications. Neuroscience 2015, 290, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Honda, M.; Tanabe, M.; Ono, H. Noxious stimuli evoke a biphasic flexor reflex composed of A delta-fiber-mediated short-latency and C-fiber-mediated long-latency withdrawal movements in mice. J. Pharmacol. Sci. 2004, 95, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajac, A.; Chalimoniuk, M.; Maszczyk, A.; Golas, A.; Lngfort, J. Central and Peripheral Fatigue During Resistance Exercise—A Critical Review. J. Hum. Kinet 2015, 49, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Ondarza, A.B.; Ye, Z.; Hulsebosch, C.E. Direct evidence of primary afferent sprouting in distant segments following spinal cord injury in the rat: Colocalization of GAP-43 and CGRP. Exp. Neurol. 2003, 184, 373–380. [Google Scholar] [CrossRef]
- Detloff, M.R.; Quiros-Molina, D.; Javia, A.S.; Daggubati, L.; Nehlsen, A.D.; Naqvi, A.; Ninan, V.; Vannix, K.N.; McMullen, M.K.; Amin, S.; et al. Delayed Exercise Is Ineffective at Reversing Aberrant Nociceptive Afferent Plasticity or Neuropathic Pain After Spinal Cord Injury in Rats. Neurorehabil. Neural. Repair. 2016, 30, 685–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedi, S.S.; Yang, Q.; Crook, R.J.; Du, J.; Wu, Z.; Fishman, H.M.; Grill, R.J.; Carlton, S.M.; Walters, E.T. Chronic spontaneous activity generated in the somata of primary nociceptors is associated with pain-related behavior after spinal cord injury. J. Neurosci. 2010, 30, 14870–14882. [Google Scholar] [CrossRef]
- Kramer, J.K.; Taylor, P.; Steeves, J.D.; Curt, A. Dermatomal somatosensory evoked potentials and electrical perception thresholds during recovery from cervical spinal cord injury. Neurorehabil. Neural. Repair 2010, 24, 309–317. [Google Scholar] [CrossRef]
- Inanici, F.; Samejima, S.; Gad, P.; Edgerton, V.R.; Hofstetter, C.P.; Moritz, C.T. Transcutaneous Electrical Spinal Stimulation Promotes Long-Term Recovery of Upper Extremity Function in Chronic Tetraplegia. IEEE Trans. Neural. Syst. Rehabil. Eng. 2018, 26, 1272–1278. [Google Scholar] [CrossRef]
- Osuagwu, B.A.C.; Timms, S.; Peachment, R.; Dowie, S.; Thrussell, H.; Cross, S.; Shirley, R.; Segura-Fragoso, A.; Taylor, J. Home-based rehabilitation using a soft robotic hand glove device leads to improvement in hand function in people with chronic spinal cord injury:a pilot study. J. Neuroeng. Rehabil. 2020, 17, 40. [Google Scholar] [CrossRef]
- McDonald, C.G.; Sullivan, J.L.; Dennis, T.A.; O’Malley, M.K. A Myoelectric Control Interface for Upper-Limb Robotic Rehabilitation Following Spinal Cord Injury. IEEE Trans. Neural. Syst. Rehabil. Eng. 2020, 28, 978–987. [Google Scholar] [CrossRef]
- Foldes, S.T.; Boninger, M.L.; Weber, D.J.; Collinger, J.L. Effects of MEG-based neurofeedback for hand rehabilitation after tetraplegia: Preliminary findings in cortical modulations and grip strength. J. Neural. Eng. 2020, 17, 026019. [Google Scholar] [CrossRef] [PubMed]
- Roy, F.D.; Yang, J.F.; Gorassini, M.A. Afferent regulation of leg motor cortex excitability after incomplete spinal cord injury. J. Neurophysiol. 2010, 103, 2222–2233. [Google Scholar] [CrossRef] [PubMed]
- Rodionov, A.; Savolainen, S.; Kirveskari, E.; Makela, J.P.; Shulga, A. Restoration of hand function with long-term paired associative stimulation after chronic incomplete tetraplegia: A case study. Spinal. Cord Ser. Cases 2019, 5, 81. [Google Scholar] [CrossRef]
- Dunkelberger, N.; Schearer, E.M.; O’Malley, M.K. A review of methods for achieving upper limb movement following spinal cord injury through hybrid muscle stimulation and robotic assistance. Exp. Neurol. 2020, 328, 113274. [Google Scholar] [CrossRef] [PubMed]
- Corbett, E.A.; Sachs, N.A.; Kording, K.P.; Perreault, E.J. Multimodal decoding and congruent sensory information enhance reaching performance in subjects with cervical spinal cord injury. Front. Neurosci. 2014, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Battistuzzo, C.R.; Zoghi, M.; Galea, M.P. Effects of training on upper limb function after cervical spinal cord injury: A systematic review. Clin. Rehabil. 2015, 29, 3–13. [Google Scholar] [CrossRef]
- Hurd, C.; Weishaupt, N.; Fouad, K. Anatomical correlates of recovery in single pellet reaching in spinal cord injured rats. Exp. Neurol. 2013, 247, 605–614. [Google Scholar] [CrossRef]
- Garcia-Alias, G.; Truong, K.; Shah, P.K.; Roy, R.R.; Edgerton, V.R. Plasticity of subcortical pathways promote recovery of skilled hand function in rats after corticospinal and rubrospinal tract injuries. Exp. Neurol. 2015, 266, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Fenrich, K.K.; May, Z.; Torres-Espin, A.; Forero, J.; Bennett, D.J.; Fouad, K. Single pellet grasping following cervical spinal cord injury in adult rat using an automated full-time training robot. Behav. Brain Res. 2016, 299, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Fenrich, K.K.; Hallworth, B.W.; Vavrek, R.; Raposo, P.J.F.; Misiaszek, J.E.; Bennett, D.J.; Fouad, K.; Torres-Espin, A. Self-directed rehabilitation training intensity thresholds for efficient recovery of skilled forelimb function in rats with cervical spinal cord injury. Exp. Neurol. 2021, 339, 113543. [Google Scholar] [CrossRef]
- Garcia-Ramirez, D.L.; Ha, N.T.B.; Bibu, S.; Stachowski, N.J.; Dougherty, K.J. Spinal cord injury alters spinal Shox2 interneurons by enhancing excitatory synaptic input and serotonergic modulation while maintaining intrinsic properties in mouse. J. Neurosci. 2021, 41, 5833–5848. [Google Scholar] [CrossRef]
- Detloff, M.R.; Smith, E.J.; Quiros Molina, D.; Ganzer, P.D.; Houle, J.D. Acute exercise prevents the development of neuropathic pain and the sprouting of non-peptidergic (GDNF- and artemin-responsive) c-fibers after spinal cord injury. Exp. Neurol. 2014, 255, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Nees, T.A.; Tappe-Theodor, A.; Sliwinski, C.; Motsch, M.; Rupp, R.; Kuner, R.; Weidner, N.; Blesch, A. Early-onset treadmill training reduces mechanical allodynia and modulates calcitonin gene-related peptide fiber density in lamina III/IV in a mouse model of spinal cord contusion injury. Pain 2016, 157, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Sliwinski, C.; Nees, T.A.; Puttagunta, R.; Weidner, N.; Blesch, A. Sensorimotor Activity Partially Ameliorates Pain and Reduces Nociceptive Fiber Density in the Chronically Injured Spinal Cord. J. Neurotraum. 2018, 35, 2222–2238. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.V.; Rees, K.M.; Seibt, E.J.; Wood, B.D.; Wade, A.D.; Morehouse, J.; Shum-Siu, A.; Magnuson, D.S.K. Electromyographic patterns of the rat hindlimb in response to muscle stretch after spinal cord injury. Spinal. Cord 2018, 56, 560–568. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.V.; Hainline, C.; Rees, K.; Krupp, S.; Prince, D.; Wood, B.D.; Shum-Siu, A.; Burke, D.A.; Petruska, J.C.; Magnuson, D.S.K. Nociceptor-dependent locomotor dysfunction after clinically-modeled hindlimb muscle stretching in adult rats with spinal cord injury. Exp. Neurol. 2019, 318, 267–276. [Google Scholar] [CrossRef]
- De Los Reyes-Guzman, A.; Lozano-Berrio, V.; Alvarez-Rodriguez, M.; Lopez-Dolado, E.; Ceruelo-Abajo, S.; Talavera-Diaz, F.; Gil-Agudo, A. RehabHand: Oriented-tasks serious games for upper limb rehabilitation by using Leap Motion Controller and target population in spinal cord injury. NeuroRehabilitation 2021, 48, 365–373. [Google Scholar] [CrossRef] [PubMed]
- De Miguel-Rubio, A.; Rubio, M.D.; Alba-Rueda, A.; Salazar, A.; Moral-Munoz, J.A.; Lucena-Anton, D. Virtual Reality Systems for Upper Limb Motor Function Recovery in Patients With Spinal Cord Injury: Systematic Review and Meta-Analysis. JMIR Mhealth Uhealth 2020, 8, e22537. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, J.R.; Detloff, M.R. Plasticity in Cervical Motor Circuits following Spinal Cord Injury and Rehabilitation. Biology 2021, 10, 976. https://doi.org/10.3390/biology10100976
Walker JR, Detloff MR. Plasticity in Cervical Motor Circuits following Spinal Cord Injury and Rehabilitation. Biology. 2021; 10(10):976. https://doi.org/10.3390/biology10100976
Chicago/Turabian StyleWalker, John R., and Megan Ryan Detloff. 2021. "Plasticity in Cervical Motor Circuits following Spinal Cord Injury and Rehabilitation" Biology 10, no. 10: 976. https://doi.org/10.3390/biology10100976