Multimodal Role of Amino Acids in Microbial Control and Drug Development


Abstract
:1. Introduction
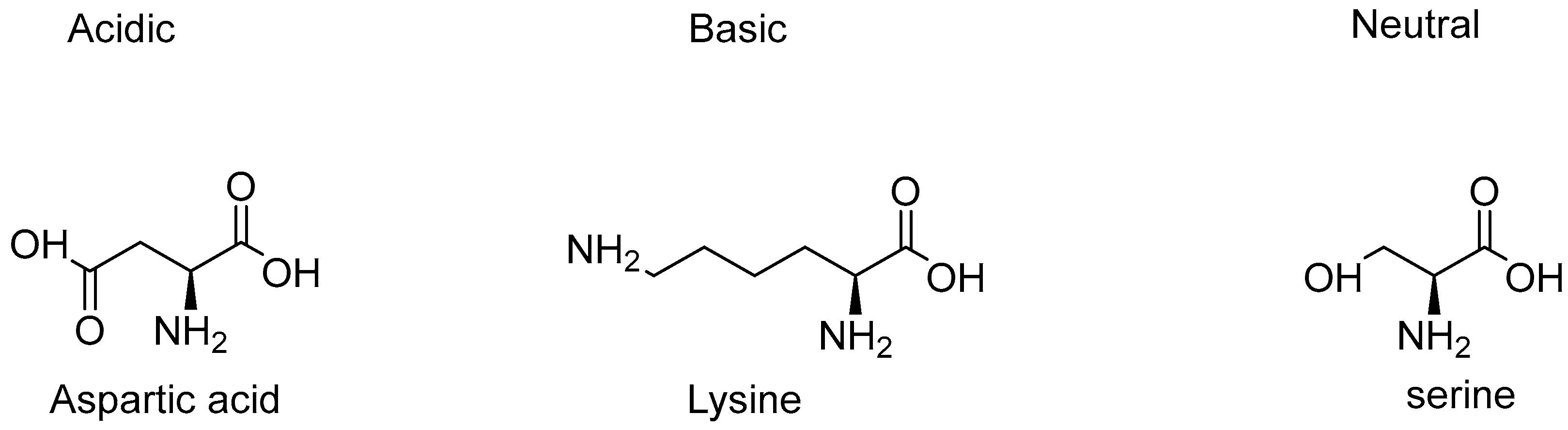
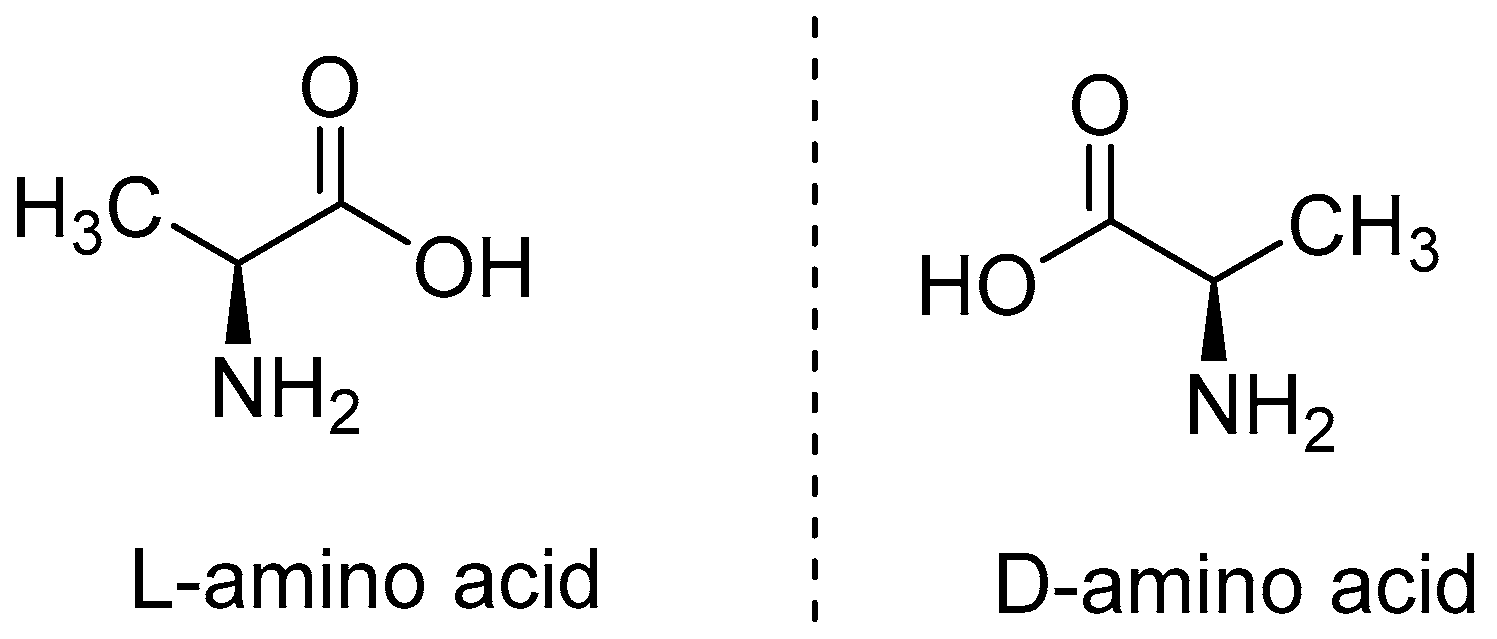
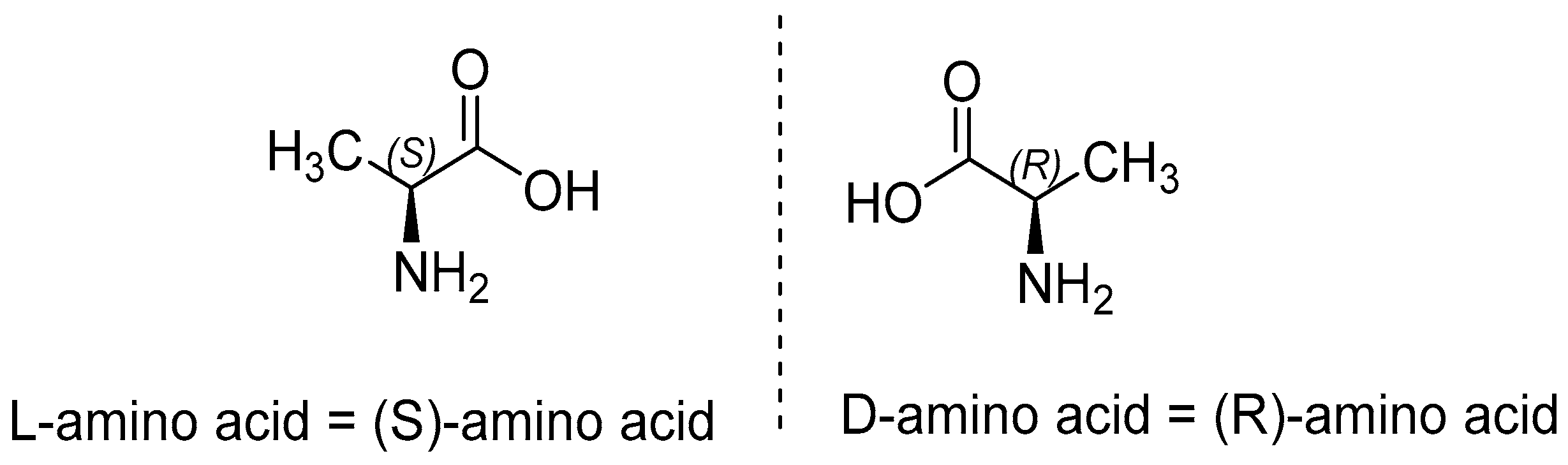
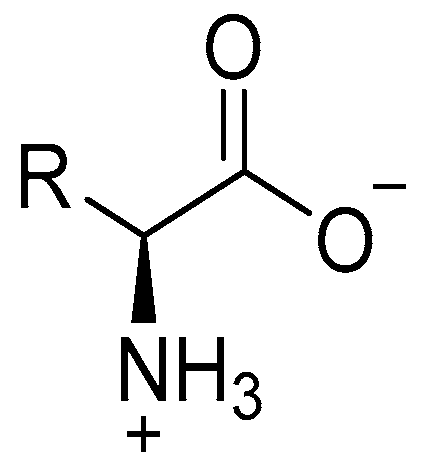
Types of Amino Acids and Chirality
2. Microbial Biosynthesis of Amino Acids
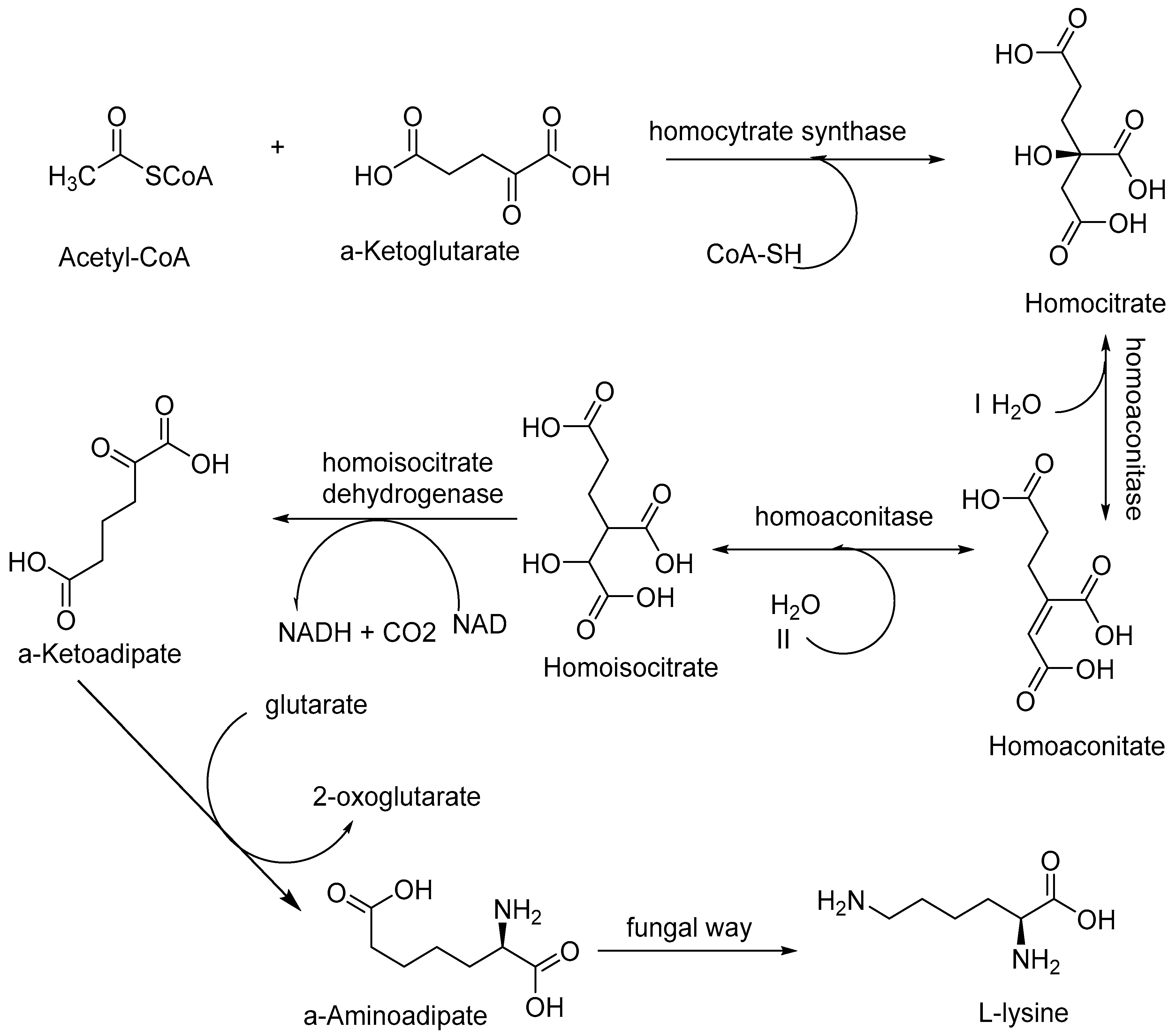
2.1. Fungal Biosynthesis of Amino Acids
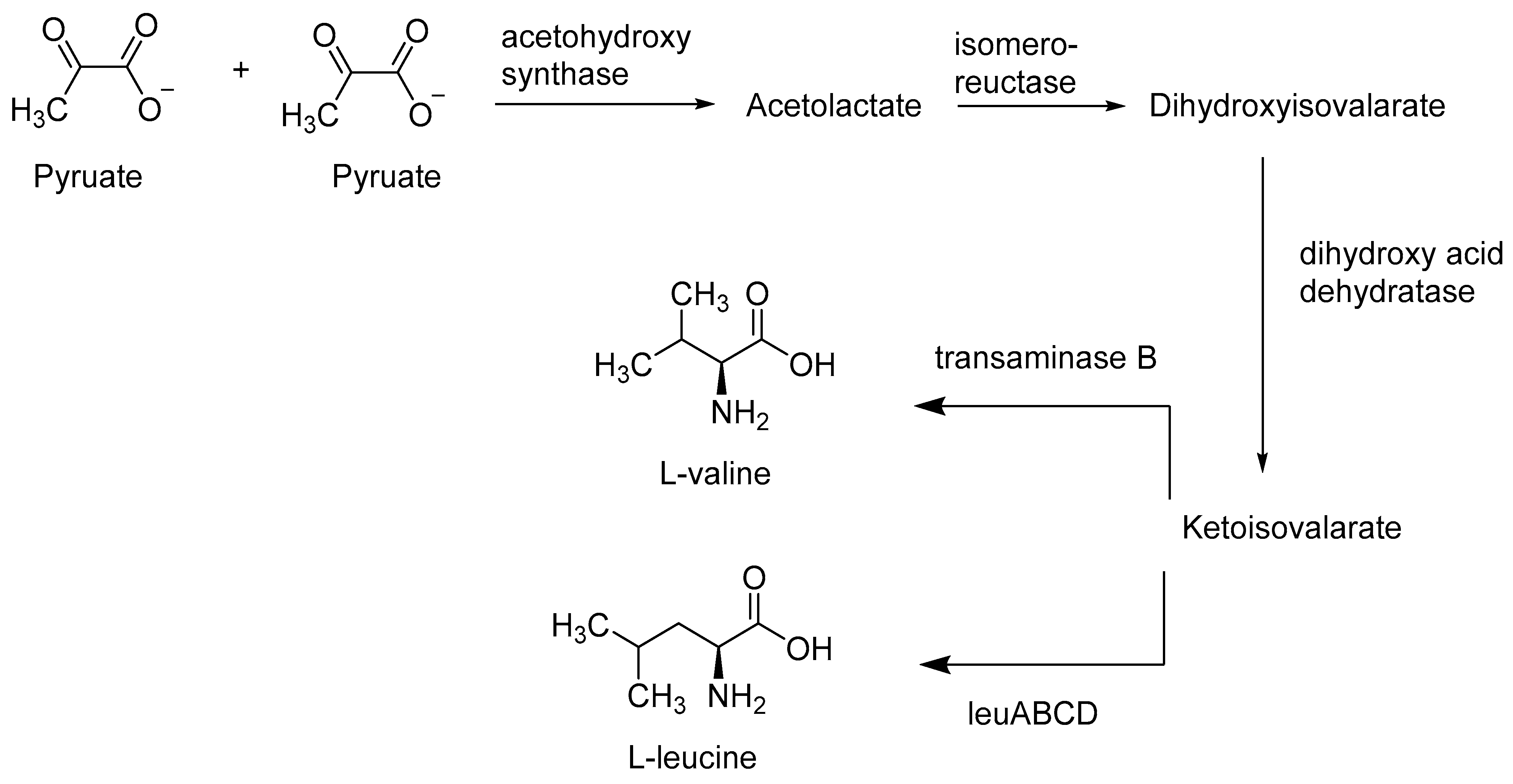
2.2. Bacterial Biosynthesis of Amino Acids
3. Antimicrobial Resistance (AMR) and Role of Amino Acids
4. Role of Amino Acids in Microbial Infections and Quorum Sensing
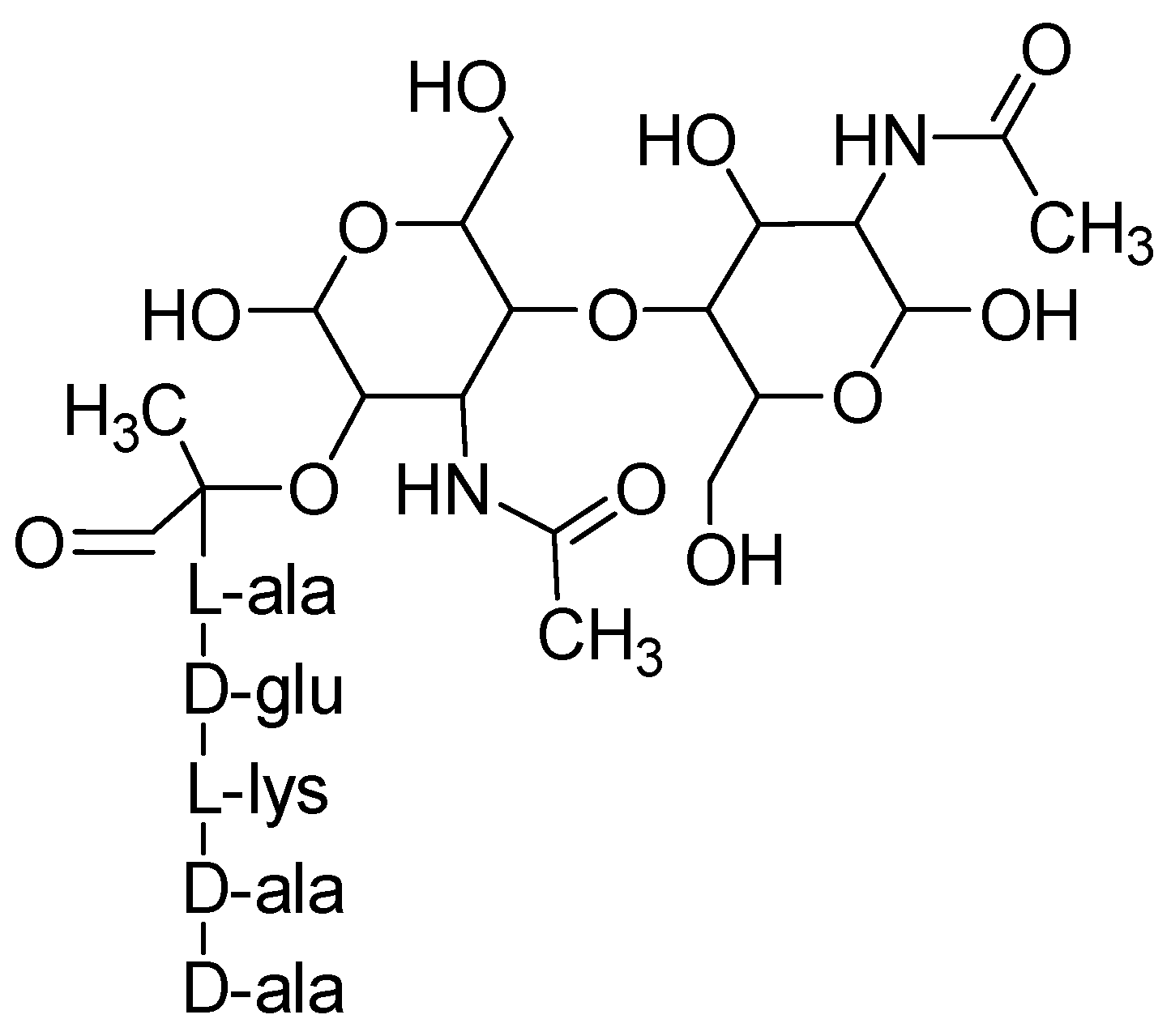
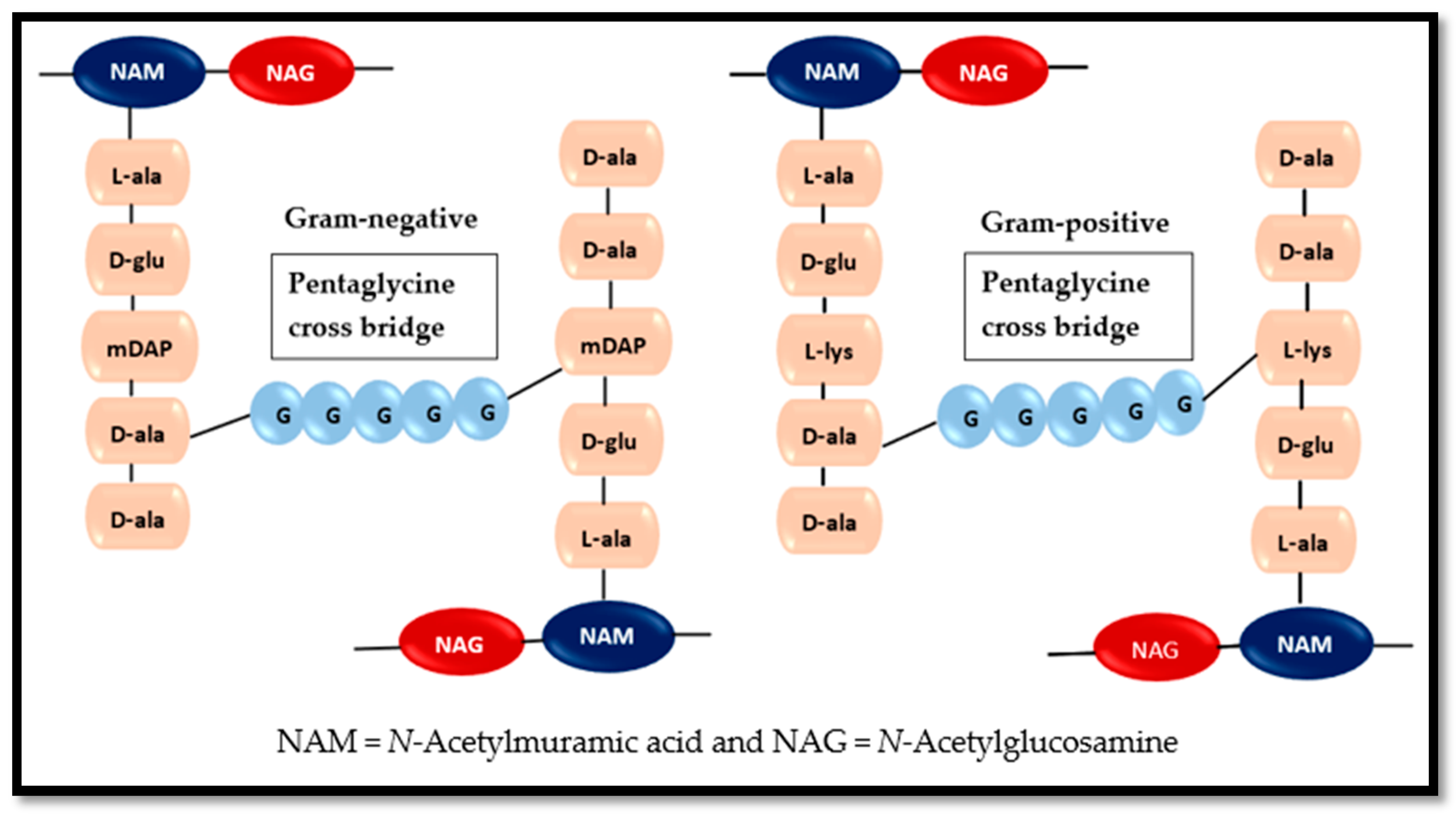
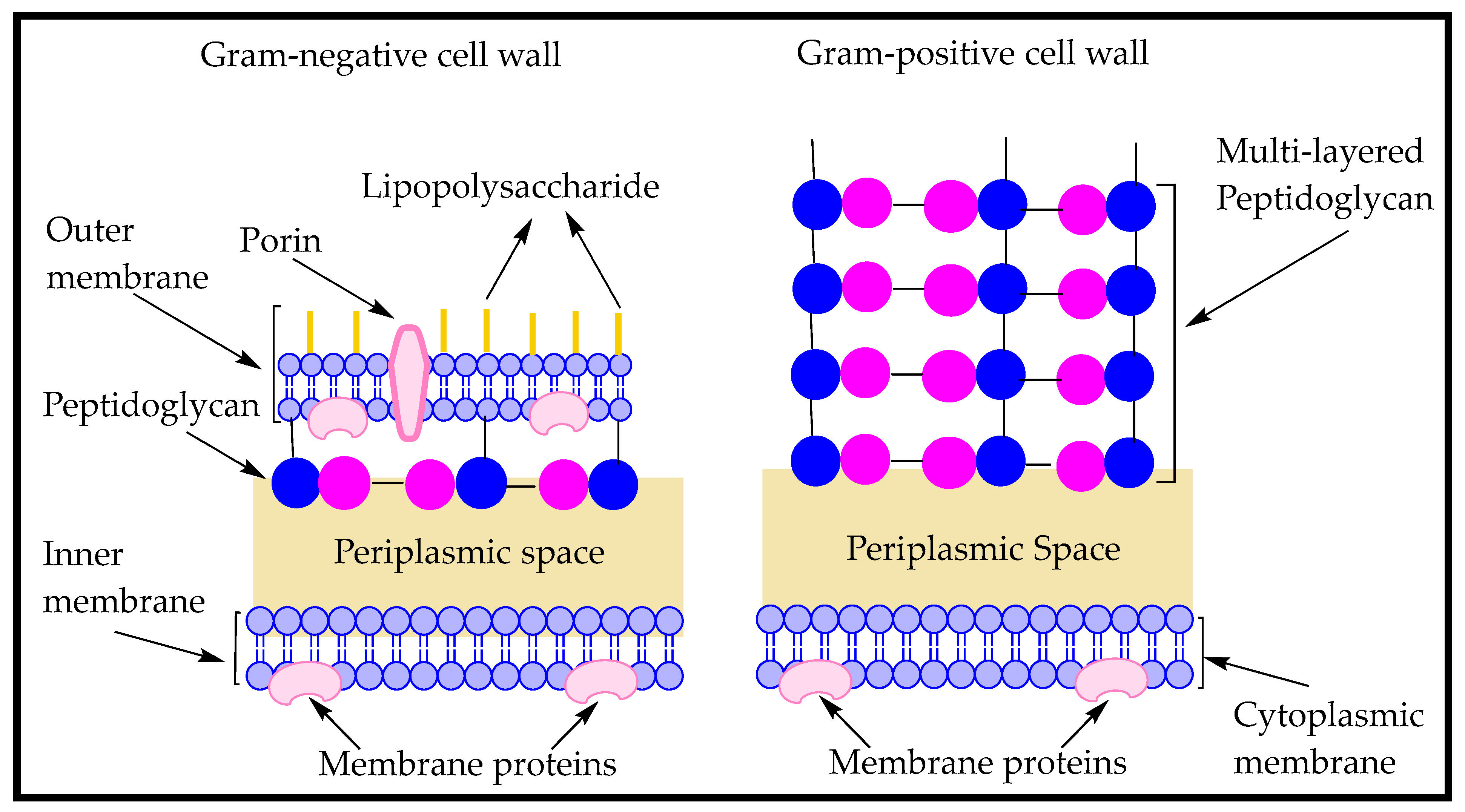
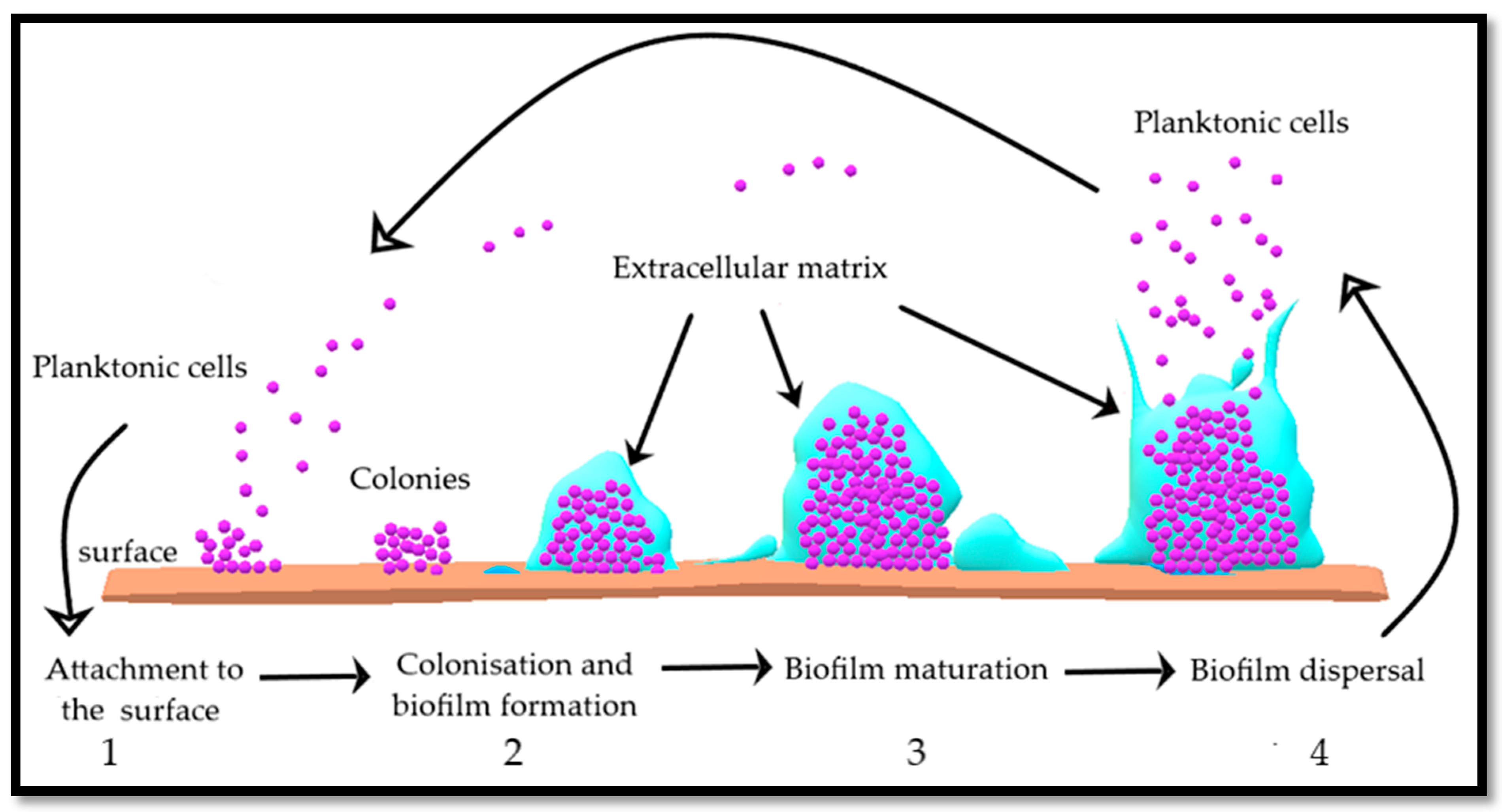
5. Role of Amino Acids in Bacterial Cell Wall Formation and Controlling Planktonic Bacteria
6. d-Amino Acids as Antibiofilm Agents, Adjuvants, and Potential Drugs Excipients
7. Amino Acids as Solubility Enhancing Agents
8. Expert Opinion
Author Contributions
Funding
Conflicts of Interest
References
- Moini, J. Epidemiology of Diet and Diabetes Mellitus. In Epidemiology of Diabetes, 1st ed.; Moini, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 57–73. [Google Scholar]
- Bischoff, R.; Schlüter, H. Amino acids: Chemistry, functionality and selected non-enzymatic post-translational modifications. J. Proteom. 2012, 75, 2275–2296. [Google Scholar] [CrossRef] [Green Version]
- Sparkman, O.D.; Penton, Z.E.; Kitson, F.G. Amino Acids. In Gas Chromatography and Mass Spectrometry; Sparkman, O.D., Penton, Z.E., Kitson, F.G., Eds.; Academic Press: Amsterdam, The Netherlands, 2011; pp. 265–271. [Google Scholar]
- Ouellette, R.J.; Rawn, J.D. Amino acids, peptides, and proteins. In Organic Chemistry Study Guide; Ouellette, R.J., Rawn, J.D., Eds.; Elsevier: Boston, MA, USA, 2015; pp. 569–586. [Google Scholar]
- Makishima, A. Fundamental knowledges and techniques in biochemistry. In Biochemistry for Materials; Makishima, A., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2019; pp. 35–51. [Google Scholar]
- Parker, J. Peptide Bond. In Encyclopedia of Genetics, 1st ed.; Brenner, S., Miller, J.H., Eds.; Academic Press: New York, NY, USA, 2001; pp. 1429–1430. [Google Scholar]
- Singh, P.; Manda, S.L.K.; Samanta, K.; Panda, G. α-Amino acids with electrically charged and polar uncharged side chains as chiral synthon: Application to the synthesis of bioactive alkaloids (1996–Dec 2013). Tetrahedron 2017, 73, 1911–2008. [Google Scholar] [CrossRef]
- Pizzarello, S. Non-Protein amino acids. In Encyclopedia of Astrobiology, 1st ed.; Gargaud, M., Amils, R., Quintanilla, J.C., Cleaves, H.J., Irvine, W.M., Pinti, D.L., Viso, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1128–1132. [Google Scholar]
- Yamane, H.; Konno, K.; Sabelis, M.; Takabayashi, J.; Sassa, T.; Oikawa, H. Chemical defence and toxins of plants. In Comprehensive Natural Products II, 1st ed.; Liu, H., Mander, L., Eds.; Elsevier: Oxford, UK, 2010; pp. 339–385. [Google Scholar]
- Ribas de Pouplana, L. Not an inside job: Non-coded amino acids compromise the genetic code. EMBO J. 2014, 33, 1619–1620. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Jander, G.; de Vos, M. Non-protein amino acids in plant defense against insect herbivores: Representative cases and opportunities for further functional analysis. Phytochemistry 2011, 72, 1531–1537. [Google Scholar] [CrossRef]
- Rodgers, K.J.; Samardzic, K.; Main, B.J. Toxic nonprotein amino acids. In Plant Toxins; Carlini, C.R., Ligabue-Braun, R., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 263–285. [Google Scholar]
- Song, Y.; Zhou, H.; Vo, M.N.; Shi, Y.; Nawaz, M.H.; Vargas-Rodriguez, O.; Diedrich, J.K.; Yates, J.R.; Kishi, S.; Musier-Forsyth, K.; et al. Double mimicry evades tRNA synthetase editing by toxic vegetable-sourced non-proteinogenic amino acid. Nat. Commun. 2017, 8, 2281. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, E.; Zhou, H.; Krasinska, K.M.; Chien, A.; Becker, C.H. Azetidine-2-carboxylic acid in garden beets (Beta vulgaris). Phytochemistry 2006, 67, 898–903. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Kevin, A.; Indira, R.; Taralyn, T. Structure & Function—Amino Acids—Biology LibreTexts. Available online: https://bio.libretexts.org/Bookshelves/Biochemistry/Book:_Biochemistry_Free_For_All_(Ahern,_Rajagopal,_and_Tan)/2:_Structure_and_Function/2.2:_Structure_and_Function_-_Amino_Acids (accessed on 12 November 2019).
- Litwack, G. Proteins. In Human Biochemistry; Academic Press: Boston, MA, USA, 2018; pp. 63–94. [Google Scholar]
- Engelking, L.R. Amino acid modifications. In Textbook of Veterinary Physiological Chemistry, 3rd ed.; Engelking, L.R., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 12–17. [Google Scholar]
- Bo-Hyun, C.; Coloff, J.L. The diverse functions of non-essential amino acids in cancer. Cancers 2019, 11, 675. [Google Scholar]
- Naef, R. A generally applicable computer algorithm based on the group additivity method for the calculation of seven molecular descriptors: Heat of combustion, LogP O/W, LogS, refractivity, polarizability, toxicity and LogBB of organic compounds; scope and limits of applicability. Molecules 2015, 20, 18279–18351. [Google Scholar]
- National Center for Biotechnology Information. PubChem Database Glutamic Acid | C5H9NO4—PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Glutamic-acid (accessed on 27 January 2020).
- Burton, A.S.; Berger, E.L. Berger Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6027462/ (accessed on 12 November 2019).
- Fujii, N.; Kaji, Y.; Fujii, N. d-Amino acids in aged proteins: Analysis and biological relevance. J. Chromatogr. B 2011, 879, 3141–3147. [Google Scholar] [CrossRef]
- Zhang, G.; Sun, H.J. Racemization in reverse: Evidence that D-Amino acid toxicity on earth is controlled by bacteria with racemases. PLoS ONE 2014, 9, e92101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.A.; He, H.; Pham-Huy, C. Chiral drugs: An overview. Int. J. Biomed. Sci. 2006, 2, 85–100. [Google Scholar] [PubMed]
- Jakubowski, H. Amino Acid Stereochemistry—Biology LibreTexts. Available online: https://bio.libretexts.org/Bookshelves/Biochemistry/Book:_Biochemistry_Online_(Jakubowski)/02:_PROTEIN_STRUCTURE/2A:_Amino_Acids/A02._Amino_Acid_Stereochemistry (accessed on 23 April 2020).
- Patzold, R.; Bruckner, H. Chiral separation of amino acids by gas chromatography. In Journal of Chromatography Library; MolnÃir-Perl, I., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 70, pp. 98–118. [Google Scholar]
- Cava, F.; Lam, H.; de Pedro, M.A.; Waldor, M.K. Emerging knowledge of regulatory roles of D-Amino acids in Bacteria. Cell. Mol. Life Sci. 2010, 68, 817–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiriyama, Y.; Nochi, H. D-Amino acids in the nervous and endocrine systems. Scientifica 2016, 216, 6494621. [Google Scholar] [CrossRef] [PubMed]
- Pawelek, P.D.; Cheah, J.; Coulombe, R.; Macheroux, P.; Ghisla, S.; Vrielink, A. The structure of L-amino acid oxidase reveals the substrate trajectory into an enantiomerically conserved active site. EMBO J. 2000, 19, 4204–4215. [Google Scholar] [CrossRef] [Green Version]
- Marcone, G.L.; Binda, E.; Rosini, E.; Abbondi, M.; Pollegioni, L. Antibacterial Properties of D-Amino acid oxidase: Impact on the food industry. Front. Microbiol. 2019, 10, 2786. [Google Scholar] [CrossRef]
- Kitani, Y.; Kikuchi, N.; Zhang, G.; Ishizaki, S.; Shimakura, K.; Shiomi, K.; Nagashima, Y. Antibacterial action of L-amino acid oxidase from the skin mucus of rockfish Sebastes schlegelii. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 149, 394–400. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Q.; Sun, M.; Teng, M.; Niu, L. Hydrogen peroxide produced by two amino acid oxidases mediates antibacterial actions. J. Microbiol. 2004, 42, 336–339. [Google Scholar]
- Berg, J.M.; Tymoczko, J.; Stryer, L. (Eds.) The Biosynthesis of Amino Acids; W. H. Freeman: New York, NY, USA, 2002; Volume 2019. [Google Scholar]
- Nakayama, K.; Araki, K.; Kase, H. Microbial production of essential amino acid with Corynebacterium glutamicum mutants. Adv. Exp. Med. Biol. 1978, 105, 649–661. [Google Scholar]
- Hernández-Montes, G.; Díaz-Mejía, J.J.; Pérez-Rueda, E.; Segovia, L. The hidden universal distribution of amino acid biosynthetic networks: A genomic perspective on their origins and evolution. Genome Biol. 2008, 9, R95. [Google Scholar] [CrossRef] [Green Version]
- Akashi, H.; Gojobori, T. Metabolic efficiency and amino acid composition in the proteomes of Escherichia coli and Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2002, 99, 3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim Franco, T.M.; Blanchard, J.S. Bacterial branched-chain amino acid biosynthesis: Structures, mechanisms, and drugability. Biochemistry 2017, 56, 5849–5865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Yang, M.; Gaur, U.; Xu, H.; Yao, Y.; Li, D. Alpha-Ketoglutarate: Physiological functions and applications. Biomol. Ther. 2016, 24, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelley, J.W. Citric acid cycle, electron transport chain, and oxidative phosphorylation. In Elsevier’s Integrated Biochemistry; Pelley, J.W., Ed.; Mosby: Philadelphia, PA, USA, 2007; pp. 55–63. [Google Scholar]
- Jastrzebowska, K.; Gabriel, I. Inhibitors of amino acids biosynthesis as antifungal agents. Amino Acids 2015, 47, 227–249. [Google Scholar] [CrossRef] [Green Version]
- Fazius, F.; Shelest, E.; Gebhardt, P.; Brock, M. The fungal α-aminoadipate pathway for lysine biosynthesis requires two enzymes of the aconitase family for the isomerization of homocitrate to homoisocitrate. Mol. Microbiol. 2012, 86, 1508–1530. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, J.; Kobashi, N.; Nishiyama, M.; Yamane, H. Functional and evolutionary relationship between arginine biosynthesis and prokaryotic lysine biosynthesis through. α-Aminoadipate. J. Bacteriol. 2001, 183, 5067. [Google Scholar] [CrossRef] [Green Version]
- Garbe, E.V. Role of amino acid metabolism in the virulence of human pathogenic fungi. Curr. Clin. Microbiol. Rep. 2019, 6, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Du, G.; Zhou, J.; Chen, J. Regulation of Amino Acid, Nucleotide, and Phosphate Metabolism in Saccharomyces Cerevisiae. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3296254/?report=reader (accessed on 25 December 2019).
- Price, M.N.; Zane, G.M.; Kuehl, J.V.; Melnyk, R.A.; Wall, J.D.; Deutschbauer, A.M.; Arkin, A.P. Filling gaps in bacterial amino acid biosynthesis pathways with high-throughput genetics. PLoS Genet. 2018, 14, 1007147. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Tsuchisaka, A.; Yukawa, H. Branched-Chain amino acids. Adv. Biochem. Eng. Biotechnol. 2017, 159, 103–128. [Google Scholar]
- Bringel, F.; Hubert, J. Extent of genetic lesions of the arginine and pyrimidine biosynthetic pathways in Lactobacillus plantarum, L. paraplantarum, L. pentosus, and L. casei: Prevalence of CO(2)-dependent auxotrophs and characterization of deficient arg genes in L. plantarum. Appl. Environ. Microbiol. 2003, 69, 2674–2683. [Google Scholar] [CrossRef] [Green Version]
- Radmacher, E.; Vaitsikova, A.; Burger, U.; Krumbach, K.; Sahm, H.; Eggeling, L. Linking Central metabolism with increased pathway flux: L-Valine accumulation by Corynebacterium glutamicum. Appl. Environ. Microbiol. 2002, 68, 2246. [Google Scholar] [CrossRef] [Green Version]
- D’Este, M.; Alvarado-Morales, M.; Angelidaki, I. Amino acids production focusing on fermentation technologies—A review. Biotechnol. Adv. 2018, 36, 14–25. [Google Scholar] [CrossRef]
- Sanchez, S.; Rodriguez-Sanoja, R.; Ramos, A.; Demain, A.L. Our microbes not only produce antibiotics, they also overproduce amino acids. J. Antibiot. 2017, 71, 26–36. [Google Scholar] [CrossRef]
- Ikeda, M. Amino acid production processes. Adv. Biochem. Eng. Biotechnol. 2003, 79, 1–35. [Google Scholar]
- Hermann, T. Industrial production of amino acids by coryneform bacteria. J. Biotechnol. 2003, 104, 155–172. [Google Scholar] [CrossRef]
- Sugden, R.; Kelly, R.; Davies, S. Combatting antimicrobial resistance globally. Nat. Microbiol. 2016, 1, 1–2. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis. Available online: https://www.who.int/news-room/detail/29–04–2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis (accessed on 20 May 2020).
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Kostyanev, T.; Can, F. The global crisis of antimicrobial resistance. In Antimicrobial Stewardship; Pulcini, C., Ergönül, Ö., Can, F., Beović, B., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 3–12. [Google Scholar]
- Hasdemir, U. The role of cell wall organization and active efflux pump systems in multidrug resistance of bacteria. Mikrobiyol. Bul. 2007, 41, 309–327. [Google Scholar]
- Nguyen Van, J.C.; Gutmann, L. Resistance to antibiotics caused by decrease of the permeability in gram-negative bacteria. Presse Med. 1994, 23, 522, 527–531. [Google Scholar]
- Egorov, A.M.; Ulyashova, M.M.; Rubtsova, M.Y. Bacterial enzymes and antibiotic resistance. Acta Nat. 2018, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.J., Jr.; Akers, K.S.; Romano, D.R.; Woodbury, R.L.; Hardy, S.K.; Murray, C.K.; Wenke, J.C. D-amino acids enhance the activity of antimicrobials against biofilms of clinical wound isolates of Staphylococcus aureus and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 4353–4361. [Google Scholar] [CrossRef] [Green Version]
- Kolodkin-Gal, I.; Romero, D.; Cao, S.; Clardy, J.; Kolter, R.; Losick, R. D-amino acids trigger biofilm disassembly. Science 2010, 328, 627–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Elshaer, A.; Hanson, P.; Worthington, T.; Lambert, P.; Mohammed, A.R. Preparation and characterization of amino acids-based trimethoprim salts. Pharmaceutics 2012, 4, 179–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ampornaramveth, R.S.; Akeatichod, N.; Lertnukkhid, J.; Songsang, N. Application of D-Amino acids as biofilm dispersing agent in dental unit waterlines. Int. J. Dent. 2018, 2018, 9413925. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.W.; Na, W.; Kim, H.O.; Yeom, M.; Park, G.; Kang, A.; Chun, H.; Park, C.; Oh, S.; Le, V.P.; et al. Cationic Poly(Amino Acid) vaccine adjuvant for promoting both cell-mediated and humoral immunity against influenza virus. Adv. Healthc. Mater. 2019, 8, 1800953. [Google Scholar] [CrossRef]
- ElShaer, A.; Ouyang, D.; Hanson, P.; Mohammed, A.R. Preparation and evaluation of amino acid based salt forms of model zwitterionic drug ciprofloxacin. J. Pharm. Drug. Deliv. Res. 2013, 2, 1–10. [Google Scholar]
- Kaiko, G.E.; Stappenbeck, T.S. Host-microbe interactions shaping the gastrointestinal environment. Trends Immunol. 2014, 35, 538–548. [Google Scholar] [CrossRef] [Green Version]
- Bahia, D.; Satoskar, A.R.; Dussurget, O. Editorial: Cell signaling in host-pathogen interactions: The host point of view. Front. Immunol. 2018, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.; Rajendran, R.; Zhao, Y.; Tan, B.; Wu, G.; Bazer, F.W.; Zhu, G.; Peng, Y.; Huang, X.; Deng, J.; et al. Amino acids as mediators of metabolic cross talk between host and pathogen. Front. Immunol. 2018, 9, 319. [Google Scholar] [CrossRef] [PubMed]
- Aliashkevich, A.; Alvarez, L.; Cava, F. New insights into the mechanisms and biological roles of D-Amino acids in complex Eco-Systems. Front. Microbiol. 2018, 9, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Anuwongcharoen, N.; Malik, A.A.; Prachayasittikul, V.; Wikberg, J.E.; Nantasenamat, C. Roles of d-Amino acids on the bioactivity of host defense peptides. Int. J. Mol. Sci. 2016, 17, 1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The human microbiota in health and disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the human microbiome. Nutr. Rev. 2012, 70, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Neis, E.P.; Dejong, C.H.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [Green Version]
- Baothman, O.A.; Zamzami, M.A.; Taher, I.; Abubaker, J.; Abu-Farha, M. The role of Gut Microbiota in the development of obesity and Diabetes. Lipids Health Dis. 2016, 15, 108. [Google Scholar] [CrossRef] [Green Version]
- Bauer, M.; Knebel, J.; Lechner, M.; Pickl, P.; Frey, E. Ecological feedback in quorum-sensing microbial populations can induce heterogeneous production of autoinducers. eLife 2017, 6, 25773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatimah, N.; Pande, G.S.J.; Natrah, F.M.I.; Meritha, W.W.; Widanarni, W.; Sucipto, A.; Ekasari, J. The role of microbial quorum sensing on the characteristics and functionality of bioflocs in aquaculture systems. Aquaculture 2019, 504, 420–426. [Google Scholar] [CrossRef]
- Sturme, M.H.; Kleerebezem, M.; Nakayama, J.; Akkermans, A.D.; Vaugha, E.E.; de Vos, W.M. Cell to cell communication by autoinducing peptides in gram-positive bacteria. Antonie Van Leeuwenhoek 2002, 81, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Reading, N.C.; Sperandio, V. Quorum sensing: The many languages of bacteria. FEMS Microbiol. Lett. 2006, 254, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.; Gupta, A.K.; Kumar, M. Prediction and analysis of quorum sensing peptides based on sequence features. PLoS ONE 2015, 10, e0120066. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Chen, J.; Yang, C.; Yin, Y.; Yao, K. Quorum sensing: A prospective therapeutic target for bacterial diseases. BioMed Res. Int. 2019, 2019, 2015978. [Google Scholar] [CrossRef] [Green Version]
- Schaible, U.E.; Kaufmann, S.H.E. Malnutrition and infection: Complex mechanisms and global impacts (research in translation). PLoS Med. 2007, 4, 115. [Google Scholar] [CrossRef] [Green Version]
- Kurpad, A.V. The requirements of protein & amino acid during acute & chronic infections. Indian J. Med. Res. 2006, 124, 129–148. [Google Scholar]
- Shrestha, R.; Sorg, J.A. Hierarchical recognition of amino acid co-germinants during Clostridioides difficile spore germination. Anaerobe 2018, 49, 41–47. [Google Scholar] [CrossRef]
- Pollegioni, L.; Piubelli, L.; Sacchi, S.; Pilone, M.S.; Molla, G. Physiological functions of D-amino acid oxidases: From yeast to humans. Cell. Mol. Life Sci. 2007, 64, 1373–1394. [Google Scholar] [CrossRef]
- Eissa, M.E.; Abd El Naby, M.; Beshir, M.M. Bacterial vs. fungal spore resistance to peroxygen biocide on inanimate surfaces. Bull. Fac. Pharm. Cairo Univ. 2014, 52, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Swick, M.C.; Koehler, T.M.; Driks, A. Surviving between hosts: Sporulation and transmission. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Sorg, J.A.; Sun, X. Biology: Sporulation, germination, and corresponding therapies for infection. Front. Cell. Infect. Microbiol. 2018, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vylkova, S.; Carman, A.J.; Danhof, H.A.; Collette, J.R.; Zhou, H.; Lorenz, M.C. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. mBio 2011, 2, e00055-11. [Google Scholar] [CrossRef] [Green Version]
- Soriani, M.; Telford, J.L. Relevance of pili in pathogenic streptococci pathogenesis and vaccine development. Future Microbiol. 2010, 5, 735–747. [Google Scholar] [CrossRef]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-That, H. Pili in Gram-positive bacteria: Assembly, involvement in colonization and biofilm development. Trends Microbiol. 2008, 16, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Padhi, A.; Sengupta, M.; Sengupta, S.; Roehm, K.H.; Sonawane, A. Antimicrobial peptides and proteins in mycobacterial therapy: Current status and future prospects. Tuberculosis 2014, 94, 363–373. [Google Scholar] [CrossRef]
- Carvajal-Rondanelli, P.; Arostica, M.; Alvarez, C.A.; Ojeda, C.; Albericio, F.; Aguilar, L.F.; Marshall, S.H.; Guzman, F. Understanding the antimicrobial properties/activity of an 11-residue Lys homopeptide by alanine and proline scan. Amino Acids 2018, 50, 557–568. [Google Scholar] [CrossRef]
- She, P.; Chen, L.; Liu, H.; Zou, Y.; Luo, Z.; Koronfel, A.; Wu, Y. The effects of D-Tyrosine combined with amikacin on the biofilms of Pseudomonas aeruginosa. Microb. Pathog. 2015, 86, 38–44. [Google Scholar] [CrossRef]
- Yehuda, A.; Slamti, L.; Malach, E.; Lereclus, D.; Hayouka, Z. Elucidating the hot spot residues of quorum sensing peptidic autoinducer PapR by multiple amino acid replacements. Front. Microbiol. 2019, 10, 1246. [Google Scholar] [CrossRef]
- Santos, J.C.P.; Sousa, R.C.S.; Otoni, C.G.; Moraes, A.R.F.; Souza, V.G.L.; Medeiros, E.A.A.; Espitia, P.J.P.; Pires, A.C.S.; Coimbra, J.S.R.; Soares, N.F.F. Nisin and other antimicrobial peptides: Production, mechanisms of action, and application in active food packaging. Innov. Food Sci. Emerg. Technol. 2018, 48, 179–194. [Google Scholar] [CrossRef]
- Foster, S.J.; Popham, D.L. Structure and synthesis of cell wall, spore cortex, teichoic acids, S-Layers, and Capsules. In Bacillus Subtilis and Its Closest Relatives; American Society of Microbiology: Washington, DC, USA, 2002. [Google Scholar]
- Lam, H.; Oh, D.; Cava, F.; Takacs, C.N.; Clardy, J.; de Pedro, M.A.; Waldor, M.K. D-amino acids govern stationary phase cell wall remodeling in bacteria. Science 2009, 325, 1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Typas, A.; Banzhaf, M.; Gross, C.A.; Vollmer, W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nat. Rev. Microbiol. 2011, 10, 123–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, C.; Khan, I.A. UDP-GlcNAc pathway: Potential target for inhibitor discovery against M. tuberculosis. Eur. J. Pharm. Sci. 2016, 83, 62–70. [Google Scholar] [CrossRef]
- Lovering, A.L.; Safadi, S.S.; Strynadka, N.C. Structural perspective of peptidoglycan biosynthesis and assembly. Annu. Rev. Biochem. 2012, 81, 451–478. [Google Scholar] [CrossRef]
- Kaiser, G. The Peptidoglycan Cell Wall—Biology LibreTexts. Available online: https://bio.libretexts.org/Bookshelves/Microbiology/Book:_Microbiology_(Kaiser)/Unit_1:_Introduction_to_Microbiology_and_Prokaryotic_Cell_Anatomy/2:_The_Prokaryotic_Cell_-_Bacteria/2.3:_The_Peptidoglycan_Cell_Wall (accessed on 19 April 2020).
- Liu, Y.; Breukink, E. The membrane steps of bacterial cell wall synthesis as antibiotic targets. Antibiotics 2016, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Rohs, P.D.A.; Buss, J.; Sim, S.I.; Squyres, G.R.; Srisuknimit, V.; Smith, M.; Cho, H.; Sjodt, M.; Kruse, A.C.; Garner, E.C.; et al. A central role for PBP2 in the activation of peptidoglycan polymerization by the bacterial cell elongation machinery. PLoS Genet. 2018, 14, 1007726. [Google Scholar] [CrossRef] [PubMed]
- Popham, D.L. Visualizing the production and arrangement of peptidoglycan in Gram-positive cells. Mol. Microbiol. 2013, 88, 645–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irazoki, O.; Hernandez, S.B.; Cava, F. Peptidoglycan muropeptides: Release, perception, and functions as signaling molecules. Front. Microbiol. 2019, 10, 500. [Google Scholar] [CrossRef]
- Que, Y.; Moreillon, P. Staphylococcus aureus (Including Staphylococcal Toxic Shock Syndrome). In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; Content Repository Only: Philadelphia, PA, USA, 2015; pp. 2237–2271. [Google Scholar]
- Allison, D.G.; Lambert, P.A. Modes of action of antibacterial agents. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: Boston, MA, USA, 2015; p. 583. [Google Scholar]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harbor Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef]
- MartÃnez-Carmona, M.; Gun’ko, Y.K.; Vallet-RegÃ, M. Mesoporous silica materials as drug delivery: “The nightmare” of bacterial infection. Pharmaceutics 2018, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Caparros, M.; Pisabarro, A.G.; de Pedro, M.A. Effect of D-amino acids on structure and synthesis of peptidoglycan in Escherichia coli. J. Bacteriol. 1992, 174, 5549–5559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammes, W.; Schleifer, K.H.; Kandler, O. Mode of action of glycine on the biosynthesis of peptidoglycan. J. Bacteriol. 1973, 116, 1029–1053. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Montalban-Lopez, M.; Kuipers, O.P. Increasing the antimicrobial activity of nisin-based lantibiotics against gram-negative pathogens. Appl. Environ. Microbiol. 2018, 84, e00052-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.C.; Delves-Broughton, J. NISIN. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 4128–4135. [Google Scholar]
- Esteban, J.; García-Coca, M. Biofilms. Front. Microbiol. 2017, 8, 2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Aparna, M.S.; Yadav, S. Biofilms: Microbes and disease. Braz. J. Infect. Dis. 2008, 12, 526–530. [Google Scholar] [CrossRef] [Green Version]
- Crouzet, M.; Le Senechal, C.; BrÃzel, V.S.; Costaglioli, P.; Barthe, C.; Bonneu, M.; Garbay, B.; Vilain, S. Exploring early steps in biofilm formation: Set-up of an experimental system for molecular studies. BMC Microbiol. 2014, 14, 253. [Google Scholar] [CrossRef] [Green Version]
- Di Luca, M.; Maccari, G.; Nifosì, R. Treatment of microbial biofilms in the post-antibiotic era: Prophylactic and therapeutic use of antimicrobial peptides and their design by bioinformatics tools. Pathog. Dis. 2014, 70, 257–270. [Google Scholar] [CrossRef] [Green Version]
- Tanner, M.E. Understanding nature’s strategies for enzyme-catalyzed racemization and epimerization. Acc. Chem. Res. 2002, 35, 237–246. [Google Scholar] [CrossRef]
- Chen, D.; Cao, Y.; Yu, L.; Tao, Y.; Zhou, Y.; Zhi, Q.; Lin, H. Characteristics and influencing factors of amyloid fibers in S. mutans biofilm. AMB Express 2019, 9, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochbaum, A.I.; Kolodkin-Gal, I.; Foulston, L.; Kolter, R.; Aizenberg, J.; Losick, R. Inhibitory effects of d-Amino acids on staphylococcus aureus biofilm development. J. Bacteriol. 2011, 193, 5616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Z.; Zhang, L.; Ling, J.; Jian, Y.; Huang, L.; Deng, D.; Tandheelkunde, P. An in vitro study on the effect of free amino acids alone or in combination with nisin on biofilms as well as on planktonic bacteria of Streptococcus mutans. PLoS ONE 2014, 9, e99513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiers, A.J.; Bohannon, J.; Gehrig, S.M.; Rainey, P.B. Biofilm formation at the air-liquid interface by the Pseudomonas fluorescens SBW25 wrinkly spreader requires an acetylated form of cellulose. Mol. Microbiol. 2003, 50, 15–27. [Google Scholar] [CrossRef]
- Zhao, L.; Jiang, J.; Zhu, Z.; Liao, Z.; Yao, X.; Yang, Y.; Cao, Y.; Jiang, Y. Lysine enhances the effect of amphotericin B against Candida albicans in vitro. Acta Biochim. Biophys. Sin. 2016, 48, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Baldo, V.; Bertoncello, C.; Cocchio, S.; Fonzo, M.; Pillon, P.; Buja, A.; Baldovin, T. The new pandemic influenza A/(H1N1)pdm09 virus: Is it really “new”? J. Prev. Med. Hyg. 2016, 57, 19–22. [Google Scholar]
- Li, Y.; Jia, R.; Al-Mahamedh, H.; Xu, D.; Gu, T. Enhanced biocide mitigation of field biofilm consortia by a mixture of D-Amino Acids. Front. Microbiol. 2016, 7, 896. [Google Scholar] [CrossRef]
- Xu, D.; Li, Y.; Gu, T. D-Methionine as a biofilm dispersal signaling molecule enhanced tetrakis hydroxymethyl phosphonium sulfate mitigation of Desulfovibrio vulgaris biofilm and biocorrosion pitting. Mater. Corros. 2014, 65, 837–845. [Google Scholar] [CrossRef]
- De Jonge, B.L.M.; Gage, D.; Xu, N. The carboxyl terminus of peptidoglycan stem peptides is a determinant for methicillin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 3151–3155. [Google Scholar] [CrossRef] [Green Version]
- Österberg, T.; Wadsten, T. Physical state of l-histidine after freeze-drying and long-term storage. Eur. J. Pharm. Sci. 1999, 8, 301–308. [Google Scholar] [CrossRef]
- Hansen, L.J.J.; Daoussi, R.; Vervaet, C.; Remon, J.; De Beer, T.R.M. Freeze-drying of live virus vaccines: A review. Vaccine 2015, 33, 5507–5519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, A.R.; Coombes, A.G.A.; Perrie, Y. Amino acids as cryoprotectants for liposomal delivery systems. Eur. J. Pharm. Sci. 2007, 30, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Van Winden, E.C.A. Freeze-Drying of liposomes: Theory and practice. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2003; Volume 367, pp. 99–110. [Google Scholar]
- Khan, I.; Elhissi, A.; Shah, M.; Alhnan, M.A.; Ahmed, W. Liposome-based carrier systems and devices used for pulmonary drug delivery. In Biomaterials and Medical Tribology; Davim, J.P., Ed.; Woodhead Publishing: Sawston, UK, 2013; pp. 395–443. [Google Scholar]
- Munro, H.M. Nutritional consequences of excess amino acid intake. Adv. Exp. Med. Biol. 1978, 105, 119–129. [Google Scholar]
- Mittal, B. Pharmacokinetics and preformulation. In How to Develop Robust Solid Oral Dosage Forms from Conception to Post-Approval; Mittal, B., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 17–37. [Google Scholar]
- Savjani, K.T.; Gajjar, A.K.; Savjani, J.K. Drug solubility: Importance and enhancement techniques. ISRN Pharm. 2012, 2012, 195727. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Grant, D.J.W.; Brittain, H.G. Principles of solubility. In Solvent Systems and Their Selection in Pharmaceutics and Biopharmaceutics; Augustijns, P., Brewster, M.E., Eds.; Springer: New York, NY, USA, 2007; pp. 1–27. [Google Scholar]
- Kalepu, S.; Nekkanti, V. Insoluble drug delivery strategies: Review of recent advances and business prospects. Acta Pharm. Sin. B 2015, 5, 442–453. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharm. 2019, 570, 118642. [Google Scholar] [CrossRef]
- Serajuddin, A.T. Salt formation to improve drug solubility. Adv. Drug Deliv. Rev. 2007, 59, 603–616. [Google Scholar] [CrossRef]
- Lucas, S. The Pharmacology of Indomethacin. Headache 2016, 56, 436–446. [Google Scholar] [CrossRef]
- ElShaer, A.; Khan, S.; Perumal, D.; Hanson, P.; Mohammed, A.R. Use of amino acids as counterions improves the solubility of the BCS II model drug, indomethacin. Curr. Drug Deliv. 2011, 8, 363–372. [Google Scholar] [CrossRef]
- Iyire, A.; Alaayedi, M.; Mohammed, A.R. Pre-formulation and systematic evaluation of amino acid assisted permeability of insulin across in vitro buccal cell layers. Sci. Rep. 2016, 6, 32498. [Google Scholar] [CrossRef]
- Kelwade, J.; Parekh, H.; Dukle, V.; Sethi, B.K. How many oral antidiabetic drugs before insulin? Indian J. Endocrinol. Metab. 2017, 21, 249–250. [Google Scholar] [CrossRef] [PubMed]
- Reijenga, J.; van Hoof, A.; van Loon, A.; Teunissen, B. Development of methods for the determination of pKa values. Anal. Chem. Insights 2013, 8, 53–71. [Google Scholar]
- Manallack, D.T. The pK(a) distribution of drugs: Application to drug discovery. Perspect. Med. Chem. 2007, 1, 25–38. [Google Scholar]
- Ouellette, R.J.; Rawn, J.D. Amino acids, peptides, and proteins. In Organic Chemistry; Academic Press: Boston, MA, USA, 2018; pp. 929–971. [Google Scholar]
- Bhattachar, S.N.; Deschenes, L.A.; Wesley, J.A. Solubility: It’s not just for physical chemists. Drug Discov. Today 2006, 11, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Kallay, N.; Kovačević, D.; Žalac, S. Thermodynamics of the solid/liquid interface - its application to adsorption and colloid stability. Interface Sci. Technol. 2006, 11, 133–170. [Google Scholar]
- Barret, R. Importance and Evaluation of the pKa. In Therapeutical Chemistry; Barret, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 21–51. [Google Scholar]
- Hadjesfandiari, N.; Parambath, A. Stealth coatings for nanoparticles: Polyethylene glycol alternatives. In Engineering of Biomaterials for Drug Delivery Systems; Parambath, A., Ed.; Woodhead Publishing: Cambridge, UK, 2018; pp. 345–361. [Google Scholar]
- Moldoveanu, S.C.; David, V. Properties of analytes and matrices determining HPLC selection. In Selection of the HPLC Method in Chemical Analysis; Moldoveanu, S.C., David, V., Eds.; Elsevier: Boston, MA, USA, 2017; pp. 189–230. [Google Scholar]
- Haynes, W.M.; Lide, D.R.; Bruno, T.J. Properties of amino acids. In CRC Handbook of Chemistry and Physics, 97th ed.; Haynes, W.M., Lide, D.R., Bruno, T.J., Eds.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Kundukad, B.; Udayakumar, G.; Grela, E.; Kaur, D.; Rice, S.A.; Kjelleberg, S.; Doyle, P.S. Weak acids as an alternative anti-microbial therapy. Biofilm 2020, 2, 100019. [Google Scholar] [CrossRef]
- Doran, P.M. Unit operations. In Bioprocess Engineering Principles, 2nd ed.; Academic Press: London, UK, 2013; pp. 445–595. [Google Scholar]
- Aftabuddin, M.; Kundu, S. Hydrophobic, hydrophilic, and charged amino acid networks within protein. Biophys. J. 2007, 93, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Lodish, H.; Berk, A.; Zipursky, S.L.; Matsudaira, P.; Baltimore, D.; Darnell, J. Hierarchical Structure of Proteins—Molecular Cell Biology—NCBI Bookshelf. Available online: https://www.ncbi.nlm.nih.gov/books/NBK21581/ (accessed on 17 November 2019).
- Manallack, D.T.; Prankerd, R.J.; Yuriev, E.; Oprea, T.I.; Chalmers, D.K. The significance of acid/base properties in drug discovery. Chem. Soc. Rev. 2013, 42, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Nadendla, K.; Friedman, S.H. Light control of protein solubility through isoelectric point modulation. J. Am. Chem. Soc. 2017, 139, 17861–17869. [Google Scholar] [CrossRef]
- Fischel-Ghodsian, F.; Brown, L.; Mathiowitz, E.; Brandenburg, D.; Langer, R. Enzymatically controlled drug delivery. Proc. Natl. Acad. Sci. USA 1988, 85, 2403–2406. [Google Scholar] [CrossRef] [Green Version]
- Acharya, P.C.; Marwein, S.; Mishra, B.; Ghosh, R.; Vora, A.; Tekade, R.K. Role of salt selection in drug discovery and development. In Dosage Form Design Considerations; Tekade, R.K., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 435–472. [Google Scholar]
- Neau, S.H.; Loka, N.C. Pharmaceutical Salts. In Water-Insoluble Drug Formulation; Liu, R., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 451–469. [Google Scholar]
- Gupta, D.; Bhatia, D.; Dave, V.; Sutariya, V.; Varghese Gupta, S. Salts of therapeutic agents: Chemical, physicochemical, and biological considerations. Molecules 2018, 23, 1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, G. Chemistry and Biochemistry of the Amino Acids; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]












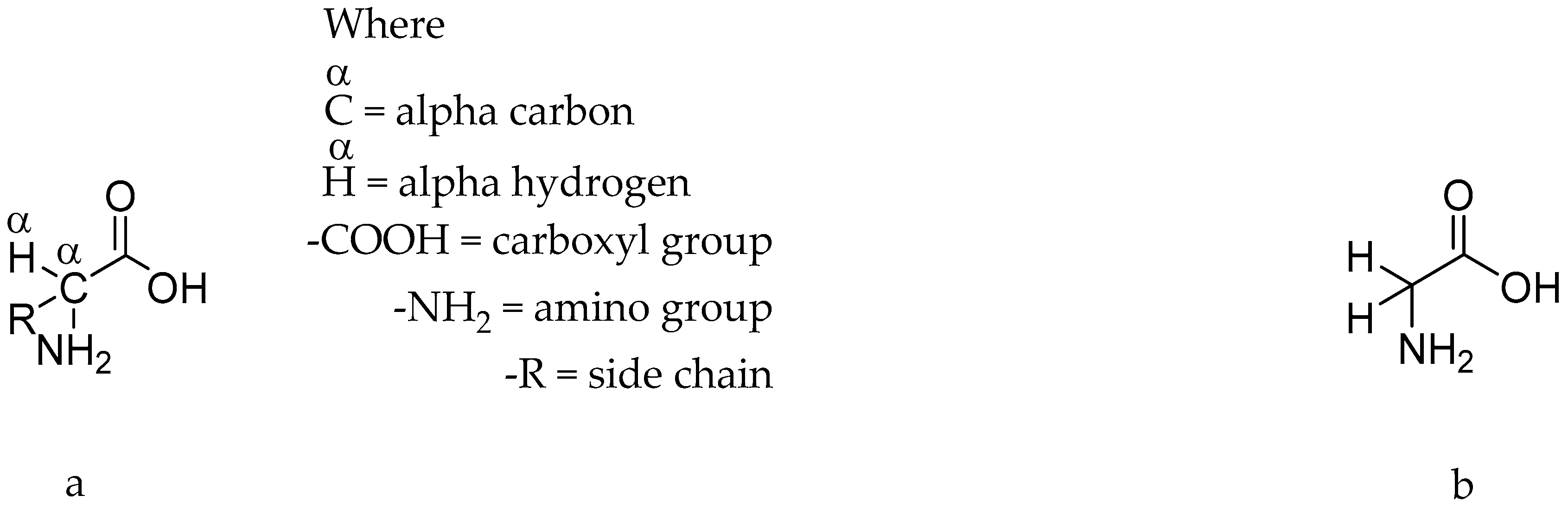{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Precursors | Intermediates | Amino Acids |
|---|---|---|
| α-ketoglutarate | Glutamate | Glutamine |
| Proline | ||
| Arginine | ||
| 3-phosphoglycerate | Serine | Glycine |
| Cysteine | ||
| Oxaloacetate | Aspartate | Asparagine |
| Methionine | ||
| Isoleucine | ||
| Lysine | ||
| Phosphoenolpyruvate + Erythrose 4-phosphate | ---------------------- | Tyrosine |
| Phenylalanine | ||
| Tryptophan | ||
| Pyruvate | ----------------------- | Alanine |
| Valine | ||
| Leucine | ||
| Ribose 5-phosphate | ------------------------ | Histidine |
| Name | Acidic | Basic | Neutral | pKa1 (α-Carboxyl Group) | PKa2 (α-Amino Group) | pKa3 (Side Chain) | pI (Isoelectronic Point) |
|---|---|---|---|---|---|---|---|
| Alanine | √ | 2.34 | 9.69 | 6.00 | |||
| Arginine | √ | 2.17 | 9.04 | 12.48 | 10.76 | ||
| Asparagine | √ | 2.02 | 8.80 | 5.41 | |||
| Aspartic acid | √ | 1.88 | 9.60 | 3.65 | 2.77 | ||
| Cysteine | √ | 1.96 | 10.28 | 8.18 | 5.07 | ||
| Glutamic acid | √ | 2.19 | 9.67 | 4.25 | 3.22 | ||
| Glutamine | √ | 2.17 | 9.13 | 5.65 | |||
| Glycine | √ | 2.34 | 9.60 | 5.97 | |||
| Histidine | √ | 1.82 | 9.17 | 6.00 | 7.95 | ||
| Isoleucine | √ | 2.36 | 9.60 | 6.02 | |||
| Leucine | √ | 2.36 | 9.60 | 5.98 | |||
| Lysine | √ | 2.18 | 8.95 | 10.53 | 9.74 | ||
| Methionine | √ | 2.28 | 9.21 | 5.74 | |||
| Phenylalanine | √ | 1.83 | 9.13 | 5.48 | |||
| Proline | √ | 1.99 | 10.60 | 6.30 | |||
| Serine | √ | 2.21 | 9.15 | 5.68 | |||
| Threonine | √ | 2.09 | 9.10 | 5.60 | |||
| Tryptophan | √ | 2.83 | 9.39 | 5.89 | |||
| Tyrosine | √ | 2.20 | 9.11 | 10.07 | 5.66 | ||
| Valine | √ | 2.32 | 9.62 | 5.96 |
| Hydrophilic Amino Acids | Charged (+/−/Uncharged) | logP | Hydrophobic Amino Acids | Charged (+/−/Uncharged) | logP |
|---|---|---|---|---|---|
| Asparagine | Uncharged | −4.3 | Arginine | + | −4.2 |
| Aspartic acid | − | −3.89 | Cysteine | Uncharged | −2.49 |
| Glutamine | Uncharged | −3.64 | Isoleucine | Uncharged | −1.7 |
| Glutamic acid | − | −3.69 | Glycine | Uncharged | −3.4 |
| Histidine | + | −3.32 | Phenylalanine | Uncharged | −1.38 |
| Lysine | + | −3.05 | Serine | Uncharged | −3.07 |
| Threonine | Uncharged | −3.5 | Leucine | Uncharged | −1.6 |
| Tyrosine | Uncharged | −2.4 | Valine | Uncharged | −2.26 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Idrees, M.; Mohammad, A.R.; Karodia, N.; Rahman, A. Multimodal Role of Amino Acids in Microbial Control and Drug Development. Antibiotics 2020, 9, 330. https://doi.org/10.3390/antibiotics9060330
Idrees M, Mohammad AR, Karodia N, Rahman A. Multimodal Role of Amino Acids in Microbial Control and Drug Development. Antibiotics. 2020; 9(6):330. https://doi.org/10.3390/antibiotics9060330
Chicago/Turabian StyleIdrees, Muhammad, Afzal R. Mohammad, Nazira Karodia, and Ayesha Rahman. 2020. "Multimodal Role of Amino Acids in Microbial Control and Drug Development" Antibiotics 9, no. 6: 330. https://doi.org/10.3390/antibiotics9060330