
Effect of Essential Oil from Lippia origanoides on the Transcriptional Expression of Genes Related to Quorum Sensing, Biofilm Formation, and Virulence of Escherichia coli and Staphylococcus aureus

 , and
, and
Abstract
:1. Introduction
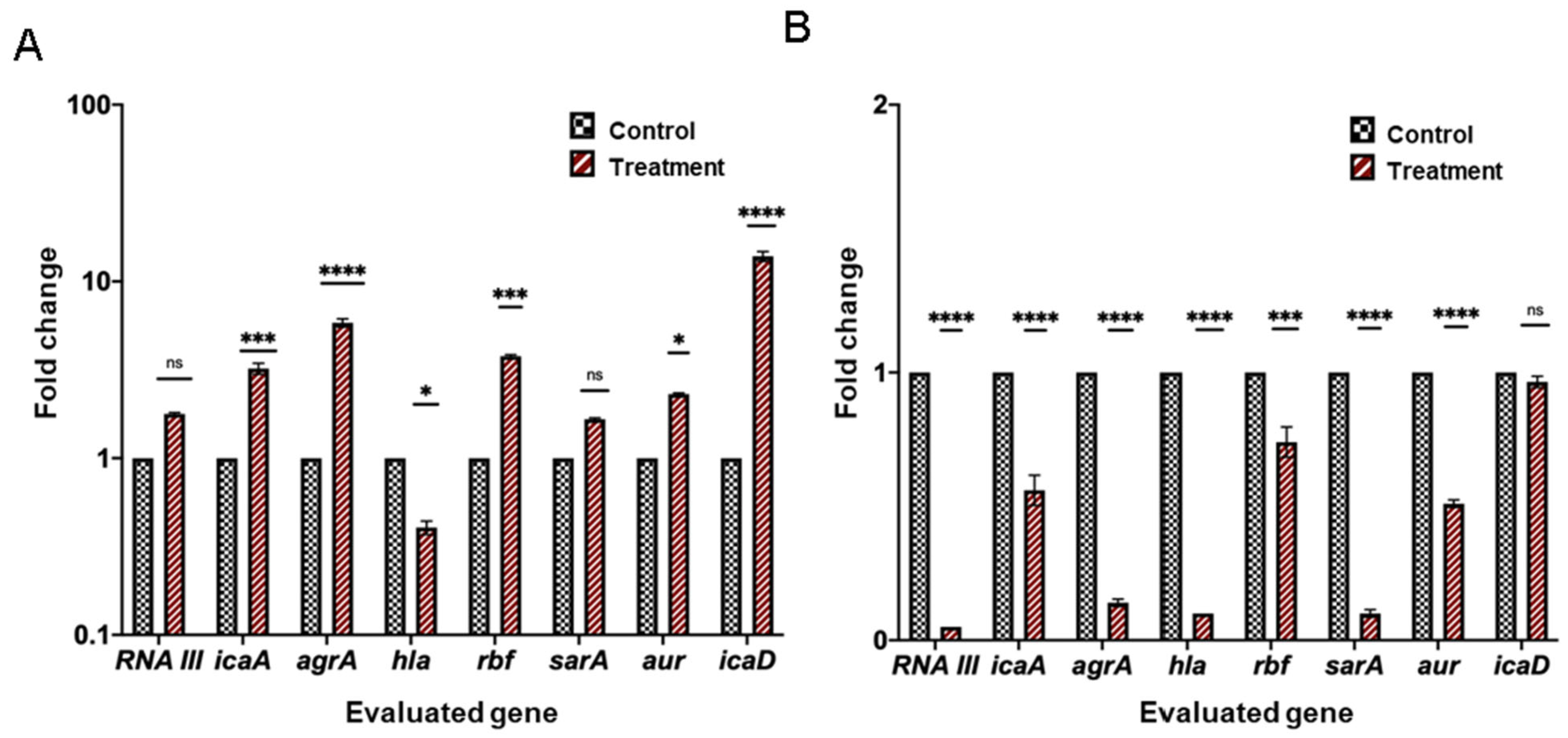
2. Results
2.1. Chemical Composition of the L. origanoides EO
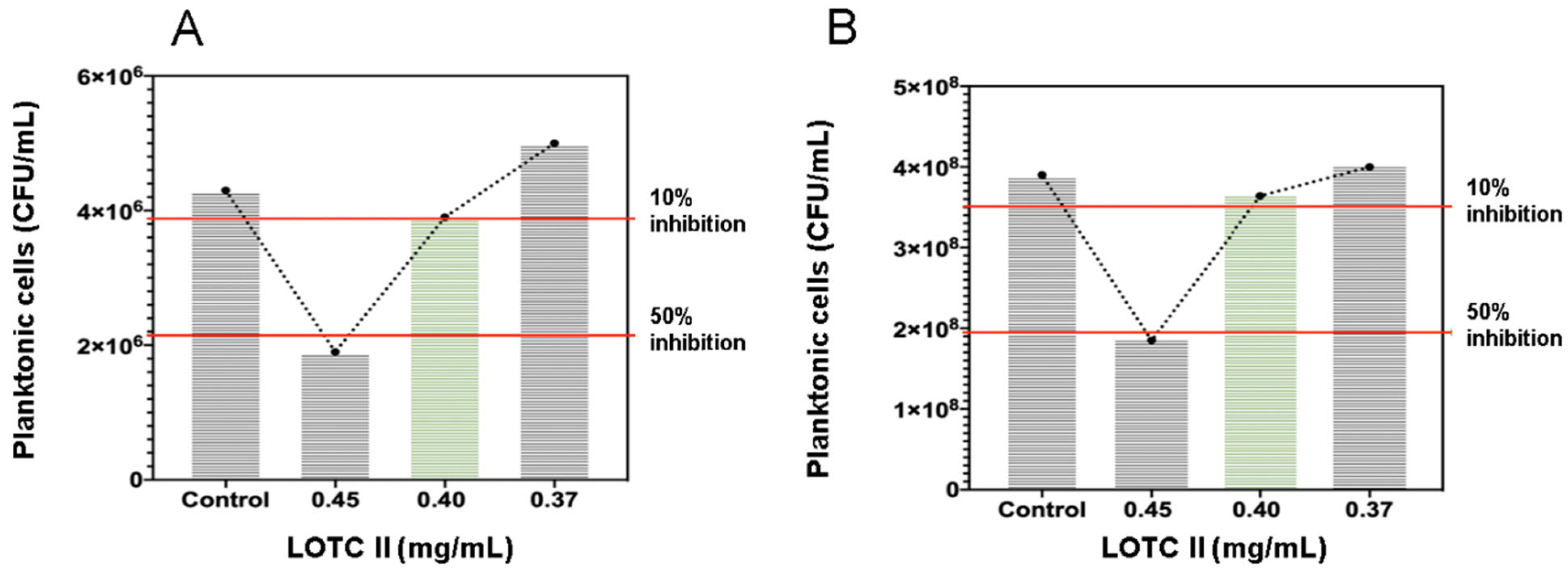
2.2. Antimicrobial Activity of the LOTC II EO on Biofilm Formation of E. coli and S. aureus
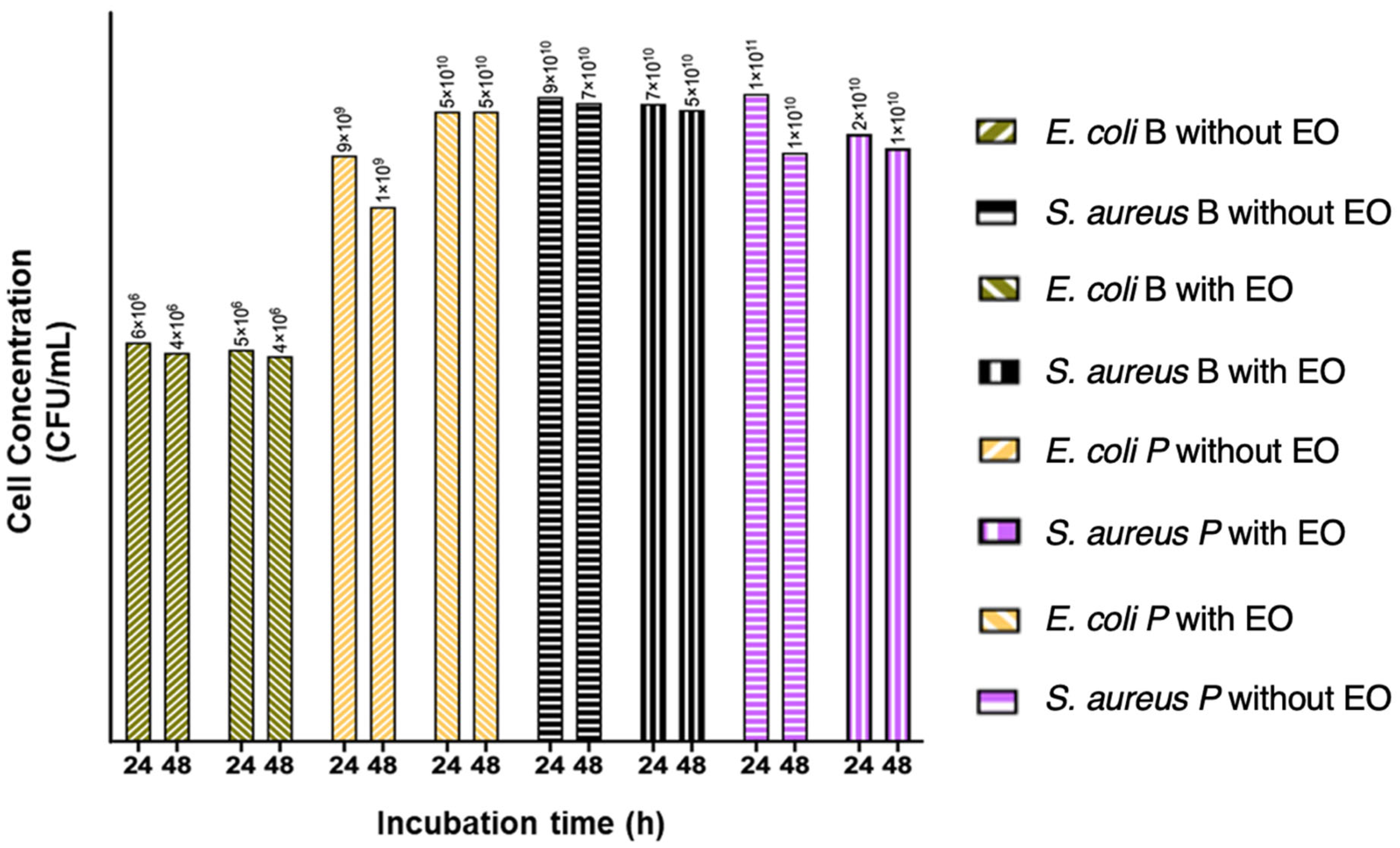
2.3. Obtaining Cell Biomass from Treated and Untreated Bacterial Biofilms with LOTC II EO in Bioreactors of 50 mL
2.4. Total RNA Extraction and cDNA Synthesis
2.5. EO Effect on Swimming Motility of E. coli ATCC 25922
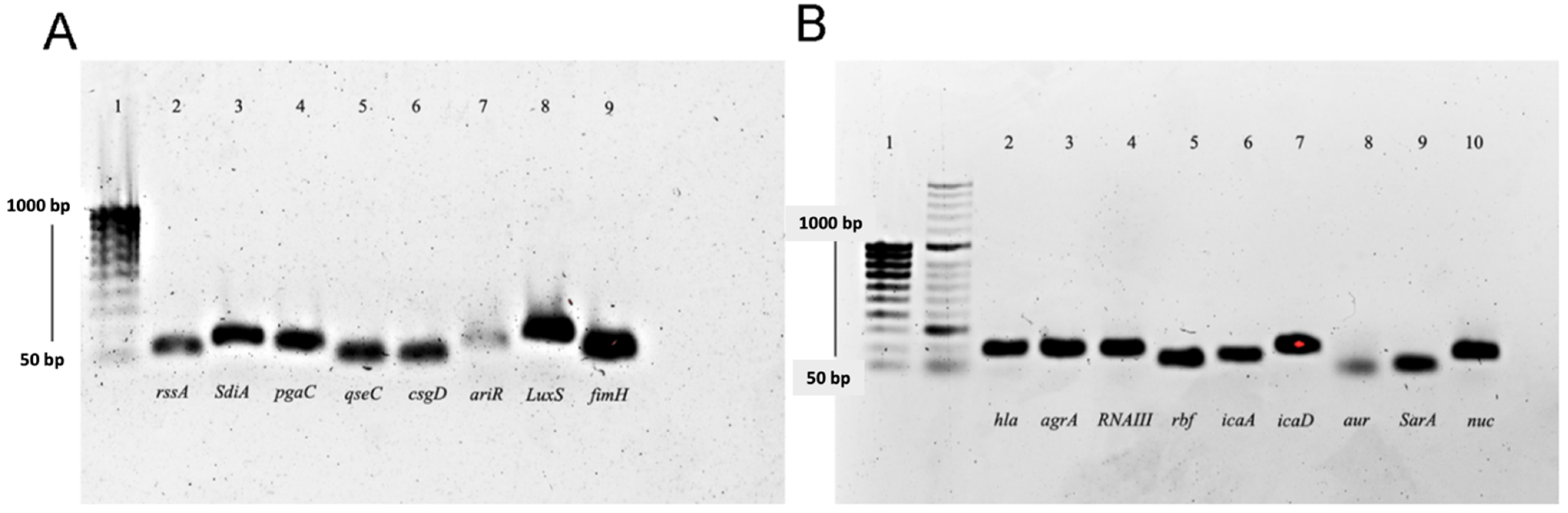
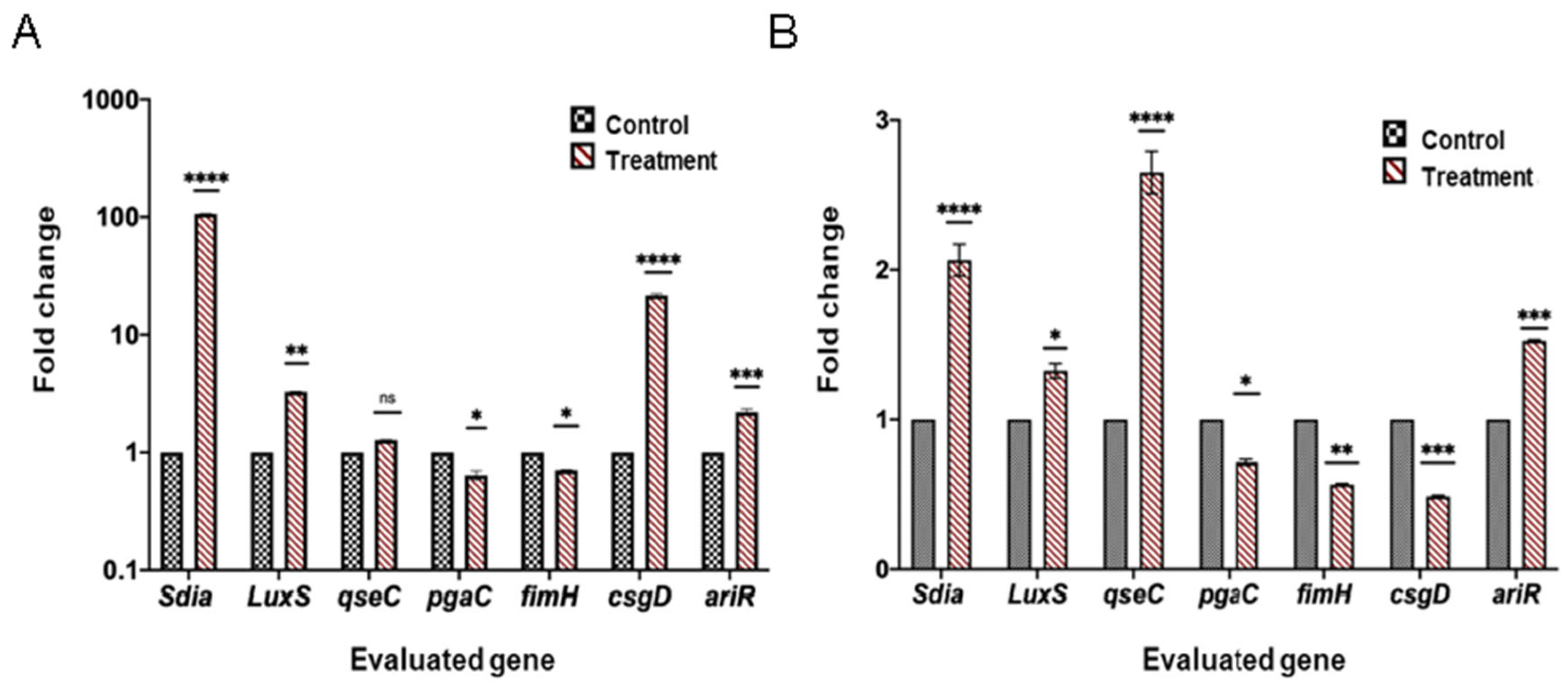
2.6. Differential Expression Analysis of Genes Related to Quorum Sensing, Biofilm Formation, and Virulence
3. Discussion
4. Materials and Methods
4.1. Materials
Bacterial Strains and Plant Material
4.2. Essential Oil Distillation and Analysis
4.3. Determination of the Minimum Inhibitory Concentration of the LOTC II EO on E. coli and S. aureus Biofilm Formation
4.4. Obtaining Biomass from Biofilm Treated and Untreated with the LOTC II EO II Bioreactors
4.5. Extraction of Total RNA and Synthesis of cDNA
4.6. Primer Design
4.7. Analysis of Differential Gene Expression
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Gales, A.C.; Castanheira, M.; Jones, R.N.; Sader, H. Antimicrobial resistance among Gram-negative bacilli isolated from Latin America: Results from SENTRY Antimicrobial Surveillance Program (Latin America, 2008–2010). Diagn. Microbiol. Infect. Dis. 2012, 73, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.; Reynolds, N.D.; Simons, M.P. Resistencia emergente a los antibióticos: Una amenaza global y un problema crítico en el cuidado de la salud. Rev. Peru. Med. Exp. Salud Pública 2015, 32, 139–145. [Google Scholar] [CrossRef]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the mechanism of bacterial biofilms resistance to antimicrobial agents. Open Microbiol. J. 2017, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Focus: Infectious diseases: Vancomycin resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269. [Google Scholar]
- Guo, Y.; Song, G.; Sun, M.; Wang, J.; Wang, Y. Prevalence and Therapies of Antibiotic-Resistance in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2020, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- David, M.Z.; Daum, R.S. Treatment of Staphylococcus aureus Infections. In Staphylococcus aureus: Microbiology, Pathology, Immunology, Therapy and Prophylaxis; Springer: Cham, Switzerland, 2017; pp. 325–383. [Google Scholar]
- Duan, J.; Li, M.; Hao, Z.; Shen, X.; Liu, L.; Jin, Y.; Wang, S.; Guo, Y.; Yang, L.; Wang, L. Subinhibitory concentrations of resveratrol reduce alpha-hemolysin production in Staphylococcus aureus isolates by downregulating saeRS. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mello, P.L.; Riboli, D.F.M.; Martins, L.D.A.; Brito, M.A.V.P.; Victória, C.; Romero, L.C.; de Souza da Cunha, M.D.L.R. Staphylococcus spp. isolated from bovine subclinical mastitis in different regions of Brazil: Molecular typing and biofilm gene expression analysis by RT-qPCR. Antibiotics 2020, 9, 888. [Google Scholar] [CrossRef]
- Kitichalermkiat, A.; Katsuki, M.; Sato, J.; Sonoda, T.; Masuda, Y.; Honjoh, K.-I.; Miyamoto, T. Effect of epigallocatechin gallate on gene expression of Staphylococcus aureus. J. Glob. Antimicrob. Resist. 2020, 22, 854–859. [Google Scholar] [CrossRef]
- Makvana, S.; Krilov, L.R. Escherichia coli infections. Pediatr. Rev. 2015, 36, 167–170. [Google Scholar] [CrossRef]
- Crim, S.M.; Griffin, P.M.; Tauxe, R.; Marder, E.P.; Gilliss, D.; Cronquist, A.B.; Cartter, M.; Tobin-D’Angelo, M.; Blythe, D.; Smith, K.; et al. Preliminary incidence and trends of infection with pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 US sites, 2006–2014. Morb. Mortal. Wkly. Rep. 2015, 64, 495. [Google Scholar]
- Gómez-Sequeda, N.; Ruiz, J.; Ortiz, C.; Urquiza, M.; Torres, R. Potent and Specific Antibacterial Activity against Escherichia coli O157:H7 and Methicillin Resistant Staphylococcus aureus (MRSA) of G17 and G19 Peptides Encapsulated into Poly-Lactic-Co-Glycolic Acid (PLGA) Nanoparticles. Antibiotics 2020, 9, 384. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, N.A.; Múnera, M.I.; López, J.A.; Sierra, P.; Robledo, C.; Robledo, J. Tendencias de la resistencia a antibióticos en Medellín y en los municipios del área metropolitana entre 2007 y 2012: Resultados de seis años de vigilancia. Biomédica 2014, 34, 433–446. [Google Scholar] [CrossRef]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) Infections: Virulence Factors, Bladder Responses, Antibiotic, and Non-antibiotic Antimicrobial Strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef] [PubMed]
- Zagaglia, C.; Ammendolia, M.G.; Maurizi, L.; Nicoletti, M.; Longhi, C. Urinary Tract Infections Caused by Uropathogenic Escherichia coli Strains—New Strategies for an Old Pathogen. Microorganisms 2022, 10, 1425. [Google Scholar] [CrossRef]
- Dinh, C.V.; Prather, K.L.J. Development of an autonomous and bifunctional quorum-sensing circuit for metabolic flux control in engineered Escherichia coli. Proc. Natl. Acad. Sci. USA 2019, 116, 25562–25568. [Google Scholar] [CrossRef]
- Shrestha, R.; Khanal, S.; Poudel, P.; Khadayat, K.; Ghaju, S.; Bhandari, A.; Lekhak, S.; Pant, N.D.; Sharma, M.; Marasini, B.P. Extended spectrum β-lactamase producing uropathogenic Escherichia coli and the correlation of biofilm with antibiotics resistance in Nepal. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 1–6. [Google Scholar] [CrossRef]
- Stanton, M.M.; Park, B.W.; Vilela, D.; Bente, K.; Faivre, D.; Sitti, M.; Sánchez, S. Magnetotactic bacteria powered biohybrids target E. coli biofilms. ACS Nano 2017, 11, 9968–9978. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential Oils and Antifungal Activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Barkauskaite, S.; Jaiswal, A.K.; Jaiswal, S. Essential oils as additives in active food packaging. Food Chem. 2021, 343, 128403. [Google Scholar] [CrossRef] [PubMed]
- Noorbakhsh, F.; Rahmati, P. Effects of Thymus vulgaris and Cinnamomum verum Essential Oils on bap and ica Gene Expression in Staphylococcus aureus. Arch. Clin. Infect. Dis. 2022, 17, e122410. [Google Scholar] [CrossRef]
- Cáceres, M.; Hidalgo, W.; Stashenko, E.; Torres, R.; Ortiz, C. Essential Oils of Aromatic Plants with Antibacterial, Anti-Biofilm and Anti-Quorum Sensing Activities against Pathogenic Bacteria. Antibiotics 2020, 9, 147. [Google Scholar] [CrossRef]
- Martínez, A.; Manrique-Moreno, M.; Klaiss-Luna, M.C.; Stashenko, E.; Zafra, G.; Ortiz, C. Effect of essential oils on growth inhibition, biofilm formation and membrane integrity of Escherichia coli and Staphylococcus aureus. Antibiotics 2021, 10, 1474. [Google Scholar] [CrossRef]
- E Stashenko, E.; E Jaramillo, B.; Martínez, J.R. Comparison of different extraction methods for the analysis of volatile secondary metabolites of Lippia alba (Mill.) N.E. Brown, grown in Colombia, and evaluation of its in vitro antioxidant activity. J. Chromatogr. A 2004, 1025, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Memar, M.Y.; Raei, P.; Alizadeh, N.; Aghdam, M.A.; Kafil, H.S. Carvacrol and thymol: Strong antimicrobial agents against resistant isolates. Rev. Med. Microbiol. 2017, 28, 63–68. [Google Scholar] [CrossRef]
- Chen, Q.; Xie, S.; Lou, X.; Cheng, S.; Liu, X.; Zheng, W.; Zheng, Z.; Wang, H. Biofilm formation and prevalence of adhesion genes among Staphylococcus aureus isolates from different food sources. MicrobiologyOpen 2020, 9, e00946. [Google Scholar] [CrossRef]
- Avila-Novoa, M.-G.; Iñíguez-Moreno, M.; Solís-Velázquez, O.-A.; González-Gómez, J.-P.; Guerrero-Medina, P.-J.; Gutiérrez-Lomelí, M. Biofilm Formation by Staphylococcus aureus Isolated from Food Contact Surfaces in the Dairy Industry of Jalisco, Mexico. J. Food Qual. 2018, 2018, 1746139. [Google Scholar] [CrossRef]
- Fitzpatrick, F.; Humphreys, H.; O’Gara, J.P. Evidence for icaADBC-Independent Biofilm Development Mechanism in Methicillin-Resistant Staphylococcus aureus Clinical Isolates. J. Clin. Microbiol. 2005, 43, 1973–1976. [Google Scholar] [CrossRef]
- Hoang, T.-M.; Zhou, C.; Lindgren, J.K.; Galac, M.R.; Corey, B.; Endres, J.E.; Olson, M.E.; Fey, P.D. Transcriptional Regulation of icaADBC by both IcaR and TcaR in Staphylococcus epidermidis. J. Bacteriol. 2019, 201, e00524-18. [Google Scholar] [CrossRef] [PubMed]
- Jeng, W.-Y.; Ko, T.-P.; Liu, C.-I.; Guo, R.-T.; Liu, C.-L.; Shr, H.-L.; Wang, A.H.-J. Crystal structure of IcaR, a repressor of the TetR family implicated in biofilm formation in Staphylococcus epidermidis. Nucleic Acids Res. 2008, 36, 1567–1577. [Google Scholar] [CrossRef] [PubMed]
- Beenken, K.E.; Blevins, J.S.; Smeltzer, M.S. Mutation of sarA in Staphylococcus aureus Limits Biofilm Formation. Infect. Immun. 2003, 71, 4206–4211. [Google Scholar] [CrossRef]
- Valle, J.; Toledo-Arana, A.; Berasain, C.; Ghigo, J.M.; Amorena, B.; Penadés, J.R.; Lasa, I. SarA and not σB is essential for biofilm development by Staphylococcus aureus. Mol. Microbiol. 2003, 48, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Valle, J.; Echeverz, M.; Lasa, I. σB inhibits poly-N-acetylglucosamine exopolysaccharide synthesis and biofilm formation in Staphylococcus aureus. J. Bacteriol. 2019, 201, e00098-19. [Google Scholar] [CrossRef]
- Blevins, J.S.; Beenken, K.E.; Elasri, M.O.; Hurlburt, B.K.; Smeltzer, M.S. Strain-dependent differences in the regulatory roles of sarA and agr in Staphylococcus aureus. Infect. Immun. 2002, 70, 470–480. [Google Scholar] [CrossRef]
- Foster, T.J. The MSCRAMM Family of Cell-Wall-Anchored Surface Proteins of Gram-Positive Cocci. Trends Microbiol. 2019, 27, 927–941. [Google Scholar] [CrossRef]
- Beenken, K.E.; Mrak, L.N.; Griffin, L.M.; Zielinska, A.K.; Shaw, L.; Rice, K.C.; Horswill, A.R.; Bayles, K.W.; Smeltzer, M.S. Epistatic Relationships between sarA and agr in Staphylococcus aureus Biofilm Formation. PLoS ONE 2010, 5, e10790. [Google Scholar] [CrossRef]
- Arora, S.; Li, X.; Hillhouse, A.; Konganti, K.; Little, S.V.; Lawhon, S.D.; Threadgill, D.; Shelburne, S.; Hook, M. Staphylococcus epidermidis MSCRAMM SesJ Is Encoded in Composite Islands. mBio 2020, 11, e02911-19. [Google Scholar] [CrossRef]
- Caballero, C.J.; Menendez-Gil, P.; Catalan-Moreno, A.; Vergara-Irigaray, M.; Garcia, B.; Segura, V.; Irurzun, N.; Villanueva, M.; Mozos, I.R.D.L.; Solano, C.; et al. The regulon of the RNA chaperone CspA and its auto-regulation in Staphylococcus aureus. Nucleic Acids Res. 2018, 46, 1345–1361. [Google Scholar] [CrossRef] [PubMed]
- Caiazza, N.C.; O’Toole, G.A. Alpha-Toxin Is Required for Biofilm Formation by Staphylococcus aureus. J. Bacteriol. 2003, 185, 3214–3217. [Google Scholar] [CrossRef]
- Anderson, M.J.; Schaaf, E.; Breshears, L.M.; Wallis, H.W.; Johnson, J.R.; Tkaczyk, C.; Sellman, B.R.; Sun, J.; Peterson, M.L. Alpha-Toxin Contributes to Biofilm Formation among Staphylococcus aureus Wound Isolates. Toxins 2018, 10, 157. [Google Scholar] [CrossRef]
- Vuong, C.; Saenz, H.L.; Götz, F.; Otto, M. Impact of the agr quorum-sensing system on adherence to polystyrene in Staphylococcus aureus. J. Infect. Dis. 2000, 182, 1688–1693. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Li, S.R.; Jiang, B.; Hu, X.M.; Li, S. Therapeutic Targeting of the Staphylococcus aureus Accessory Gene Regulator (agr) System. Front. Microbiol. 2018, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Derakhshan, S.; Navidinia, M.; Haghi, F. Antibiotic susceptibility of human-associated Staphylococcus aureus and its relation to agr typing, virulence genes, and biofilm formation. BMC Infect. Dis. 2021, 21, 1–10. [Google Scholar] [CrossRef]
- Morales, L.; Echeverz, M.; Trobos, M.; Solano, C.; Lasa, I. Diversity in regulatory regions of icaADBCR and fnbAB genes among Staphylococcus aureus strains isolated from periprosthetic joint infections (No. biofilms9-71). In Proceedings of the Copernicus Meetings, 2020. Biofilms 9 Online Conference, Karlsruhe Institute of Technology (KIT), Karlsruhe, Germany, 29 September–1 October 2020. [Google Scholar]
- Fang, B.; Liu, B.; Sun, B. Transcriptional regulation of virulence factors Hla and phenol-soluble modulins α by AraC-type regulator Rbf in Staphylococcus aureus. Int. J. Med. Microbiol. 2020, 310, 151436. [Google Scholar] [CrossRef]
- Lim, Y.; Jana, M.; Luong, T.T.; Lee, C.Y. Control of glucose-and NaCl-induced biofilm formation by rbf in Staphylococcus aureus. J. Bacteriol. 2004, 186, 722–729. [Google Scholar] [CrossRef]
- Cue, D.; Lei, M.G.; Luong, T.T.; Kuechenmeister, L.; Dunman, P.M.; O’Donnell, S.; Rowe, S.; O’Gara, J.P.; Lee, C.Y. Rbf promotes biofilm formation by Staphylococcus aureus via repression of icaR, a negative regulator of icaADBC. J. Bacteriol. 2009, 191, 6363–6373. [Google Scholar] [CrossRef]
- Prüß, B.M.; Besemann, C.; Denton, A.; Wolfe, A.J. A Complex Transcription Network Controls the Early Stages of Biofilm Development by Escherichia coli. J. Bacteriol. 2006, 188, 3731–3739. [Google Scholar] [CrossRef]
- Wood, T.K.; Barrios, A.F.G.; Herzberg, M.; Lee, J. Motility influences biofilm architecture in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 72, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Brombacher, E.; Baratto, A.; Dorel, C.; Landini, P. Gene Expression Regulation by the Curli Activator CsgD Protein: Modulation of Cellulose Biosynthesis and Control of Negative Determinants for Microbial Adhesion. J. Bacteriol. 2006, 188, 2027–2037. [Google Scholar] [CrossRef]
- Ogasawara, H.; Ishizuka, T.; Hotta, S.; Aoki, M.; Shimada, T.; Ishihama, A. Novel regulators of the csgD gene encoding the master regulator of biofilm formation in Escherichia coli K-12. Microbiology 2020, 166, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Thiriet-Rupert, S.; Mayer, L.; Beloin, C.; Ghigo, J.-M. Selection for nonspecific adhesion is a driver of FimH evolution increasing Escherichia coli biofilm capacity. Microlife 2022, 3, 1–14. [Google Scholar] [CrossRef]
- Zuberi, A.; Ahmad, N.; Khan, A.U. CRISPRi Induced Suppression of Fimbriae Gene (fimH) of a Uropathogenic Escherichia coli: An Approach to Inhibit Microbial Biofilms. Front. Immunol. 2017, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Scotti, R.; Stringaro, A.; Nicolini, L.; Zanellato, M.; Boccia, P.; Maggi, F.; Gabbianelli, R. Effects of Essential Oils from Cymbopogon spp. and Cinnamomum verum on Biofilm and Virulence Properties of Escherichia coli O157: H7. Antibiotics 2022, 10, 113. [Google Scholar] [CrossRef]
- Itoh, Y.; Rice, J.D.; Goller, C.; Pannuri, A.; Taylor, J.; Meisner, J.; Beveridge, T.J.; Preston, J.F., III; Romeo, T. Roles of pgaABCD genes in synthesis, modification, and export of the Escherichia coli biofilm adhesin poly-β-1, 6-N-acetyl-d-glucosamine. J. Bacteriol. 2008, 190, 3670–3680. [Google Scholar] [CrossRef]
- Attila, C.; Ueda, A.; Wood, T.K. 5-Fluorouracil reduces biofilm formation in Escherichia coli K-12 through global regulator AriR as an antivirulence compound. Appl. Microbiol. Biotechnol. 2009, 82, 525–533. [Google Scholar] [CrossRef]
- Wood, T.K. Insights on Escherichia coli biofilm formation and inhibition from whole-transcriptome profiling. Environ. Microbiol. 2009, 11, 1–15. [Google Scholar] [CrossRef]
- Yao, Y.; Martinez-Yamout, M.A.; Dickerson, T.J.; Brogan, A.P.; Wright, P.E.; Dyson, H.J. Structure of the Escherichia coli Quorum Sensing Protein SdiA: Activation of the Folding Switch by Acyl Homoserine Lactones. J. Mol. Biol. 2006, 355, 262–273. [Google Scholar] [CrossRef]
- Jani, S.; Seely, A.L.; Peabody V, G.L.; Jayaraman, A.; Manson, M.D. Chemotaxis to self-generated AI-2 promotes biofilm formation in Escherichia coli. Microbiology 2017, 163, 1778–1790. [Google Scholar] [CrossRef]
- Sperandio, V.; Torres, A.G.; Kaper, J.B. Quorum sensing Escherichia coli regulators B and C (QseBC): A novel two-component regulatory system involved in the regulation of flagella and motility by quorum sensing in E. coli. Mol. Microbiol. 2002, 43, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Meng, J.; Huang, Y.-C.; Ye, L.-H.; Li, G.-J.; Huang, J.; Chen, H.-M. The Role of the QseC Quorum-Sensing Sensor Kinase in Epinephrine-Enhanced Motility and Biofilm Formation by Escherichia coli. Cell Biochem. Biophys. 2014, 70, 391–398. [Google Scholar] [CrossRef]
- Lee, J.; Maeda, T.; Hong, S.H.; Wood, T.K. Reconfiguring the Quorum-Sensing Regulator SdiA of Escherichia coli to Control Biofilm Formation via Indole and N -Acylhomoserine Lactones. Appl. Environ. Microbiol. 2009, 75, 1703–1716. [Google Scholar] [CrossRef]
- Culler, H.F.; Couto, S.C.F.; Higa, J.S.; Ruiz, R.M.; Yang, M.J.; Bueris, V.; Franzolin, M.R.; Sircili, M.P. Role of SdiA on Biofilm Formation by Atypical Enteropathogenic Escherichia coli. Genes 2018, 9, 253. [Google Scholar] [CrossRef]
- Kim, T.; Duong, T.; Wu, C.A.; Choi, J.; Lan, N.; Kang, S.W.; Lokanath, N.K.; Shin, D.; Hwang, H.Y.; Kim, K.K. Structural insights into the molecular mechanism of Escherichia coli SdiA, a quorum-sensing receptor. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 694–707. [Google Scholar] [CrossRef] [PubMed]
- Stashenko, E.E.; Martínez, J.R.; Ruíz, C.A.; Arias, G.; Durán, C.; Salgar, W.; Cala, M. Lippia origanoides chemotype differentiation based on essential oil GC-MS and principal component analysis. J. Sep. Sci. 2010, 33, 93–103. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35 (Suppl. 2), W43–W46. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, R.; LaBauve, A.E.; Akoolo, L.; Patel, S.; Alqarzaee, A.A.; Lung, T.W.F.; Poorey, K.; Stinear, T.P.; Thomas, V.C.; Meagher, R.J.; et al. Dual Gene Expression Analysis Identifies Factors Associated with Staphylococcus aureus Virulence in Diabetic Mice. Infect. Immun. 2019, 87, e00163-19. [Google Scholar] [CrossRef]
- Tuttobene, M.R.; Pérez, J.F.; Pavesi, E.S.; Mora, B.P.; Biancotti, D.; Cribb, P.; Altilio, M.; Müller, G.L.; Gramajo, H.; Tamagno, G.; et al. Light Modulates Important Pathogenic Determinants and Virulence in ESKAPE Pathogens Acinetobacter baumannii, Pseudomonas aeruginosa, and Staphylococcus aureus. J. Bacteriol. 2021, 203, e00566-20. [Google Scholar] [CrossRef] [PubMed]
- Yeo, W.-S.; Anokwute, C.; Marcadis, P.; Levitan, M.; Ahmed, M.; Bae, Y.; Kim, K.; Kostrominova, T.; Liu, Q.; Bae, T. A Membrane-Bound Transcription Factor is Proteolytically Regulated by the AAA+ Protease FtsH in Staphylococcus aureus. J. Bacteriol. 2020, 202, e00019-20. [Google Scholar] [CrossRef]
- Wang, B.; Duan, J.; Jin, Y.; Zhan, Q.; Xu, Y.; Zhao, H.; Wang, X.; Rao, L.; Guo, Y.; Yu, F. Functional Insights of MraZ on the Pathogenicity of Staphylococcus aureus. Infect. Drug Resist. 2021, 14, 4539–4551. [Google Scholar] [CrossRef] [PubMed]
- Divyakolu, S.; Chikkala, R.; Kamaraju, S.; Sritharan, V. Quorum quenching as a strategy for treating Methicillin Resistant S. aureus (MRSA)—Effect of ε-Polylysine, ethanolic extracts of guava leaves and mango seed kernel. Indian J. Biochem. Biophys. 2021, 58, 171–177. [Google Scholar] [CrossRef]
- Atshan, S.S.; Shamsudin, M.N.; Karunanidhi, A.; van Belkum, A.; Lung, L.T.T.; Sekawi, Z.; Nathan, J.J.; Ling, K.H.; Seng, J.S.C.; Ali, A.M.; et al. Quantitative PCR analysis of genes expressed during biofilm development of methicillin resistant Staphylococcus aureus (MRSA). Infect. Genet. Evol. 2013, 18, 106–112. [Google Scholar] [CrossRef]
- Mahmoudi, H.; Pourhajibagher, M.; Alikhani, M.Y.; Bahador, A. The effect of antimicrobial photodynamic therapy on the expression of biofilm associated genes in Staphylococcus aureus strains isolated from wound infections in burn patients. Photodiagnosis Photodyn. Ther. 2019, 25, 406–413. [Google Scholar] [CrossRef]
- Kalinka, J.; Hachmeister, M.; Geraci, J.; Sordelli, D.; Hansen, U.; Niemann, S.; Oetermann, S.; Peters, G.; Löffler, B.; Tuchscherr, L. Staphylococcus aureus isolates from chronic osteomyelitis are characterized by high host cell invasion and intracellular adaptation, but still induce inflammation. Int. J. Med. Microbiol. 2014, 304, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Demirci, M.; Yiğin, A.; Demir, C. Efficacy of antimicrobial peptide LL-37 against biofilm forming Staphylococcus aureus strains obtained from chronic wound infections. Microb. Pathog. 2022, 162, 105368. [Google Scholar] [CrossRef]
- Ma, R.; Qiu, S.; Jiang, Q.; Sun, H.; Xue, T.; Cai, G.; Sun, B. AI-2 quorum sensing negatively regulates rbf expression and biofilm formation in Staphylococcus aureus. Int. J. Med. Microbiol. 2017, 307, 257–267. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Plant Species | Chemotype | Major Components |
|---|---|---|---|
| LOTC II | Lippia origanoides (Verbenaceae) | Thymol-carvacrol II | γ-Terpinene (5.2%), p-cymene (1.1%), thymol (32.7%), carvacrol (18.8%), and trans-β-caryophyllene (6.4%) |
| LOTC II (mg/mL) | E. coli ATCC 25922 | S. aureus ATCC 29213 | ||||
|---|---|---|---|---|---|---|
| Absorbance (OD 595 nm) | Biofilm Formation Inhibition (%) | Planktonic Cell Concentration (CFU/mL) | Absorbance (OD 595 nm) | Biofilm Formation Inhibition (%) | Planktonic Cell Concentration (CFU/mL) | |
| 0.37 | 0.410 | 24 | 5.0 × 106 | 0.730 | 19 | 4.10 × 108 |
| 0.40 | 0.134 | 76 | 3.9 × 106 | 0.253 | 72 | 3.60 × 108 |
| 0.45 | 0.034 | 94 | 1.9 × 106 | 0.045 | 95 | 1.85 × 108 |
| Control | 0.540 | - | 4.3 × 106 | 0.896 | - | 3.90 × 108 |
| Sample Condition | Concentration (ng/μL) | Absorbance Ratio (260/280) |
|---|---|---|
| Planktonic E. coli with no-treatment | 63 | 1.99 |
| Planktonic E. coli with treatment | 58 | 2.02 |
| Planktonic E. coli with no-treatment | 49 | 2.10 |
| E. coli biofilm with treatment | 43 | 1.98 |
| Planktonic E. coli with no-treatment | 120 | 2.00 |
| Planktonic S. aureus with treatment | 100 | 1.98 |
| Planktonic E. coli with no-treatment | 97 | 2.01 |
| S. aureus biofilm with treatment | 80 | 2.00 |
| Gene | Primer | Sequence 5′ to 3′ | Product Size (pb) | Tm (°C) | % GC | References |
|---|---|---|---|---|---|---|
| SdiA | SdiA 1 SdiA 2 | CGGTGCTGAACCCTGAA CGCTGCAACGGGAAAA | 177 | 59.3 60.5 | 58.8 56.2 | (This work) |
| LuxS | LuxS 1 LuxS 2 | TGTTGCTGATGCCTGGAA CTTTCGGCAGTGCCAGTT | 194 | 59.9 60.0 | 50.0 55.6 | (This work) |
| FimH | FimH 1 FimH 2 | GGCTGCGATGTTTCTGCT CCCCAGGTTTTGGCTTTT | 105 | 60.1 59.9 | 55.6 50 | (This work) |
| csgD | csgD 1 csgD 2 | CCGTACCGCGACATTGA CGCCTTGCAACCCATT | 91 | 60.2 59.1 | 58.8 56.2 | (This work) |
| ariR | ariR 1 ariR 2 | TGTTAGGGCAGGCTGTCA TCGCAACACGATTTCCAG | 149 | 58.9 59.3 | 55.6 50.0 | (This work) |
| pgaC | pgaC 1 pgaC 2 | TTGATGGCGATGCGTTATTA GGAATACTCGCCAACCTGAA | 153 | 60.1 60.1 | 40 50 | (This work) |
| qseC | qseC 1 qseC 2 | ACCCACGACGGCAGAAT GCCCGTCAGCAAAACCT | 88 | 60.1 59.8 | 58.8 58.8 | (This work) |
| RNAr | rssA 1 rssA 2 | AGGTGATCCGCCCGATA CGGCAAAAGTTCGTCCA | 130 | 60.0 59.3 | 58.8 52.9 | (This work) |
| Gene | Primer | Sequence 5′ to 3′ | Product Size (pb) | Tm (°C) | % GC | References |
|---|---|---|---|---|---|---|
| hla | Hla 1 Hla 2 | GGCCTTATTGGTGCAAATGT CCATATACCGGGTTCCAAGA | 176 | 59.8 59.6 | 45 50 | [72,73,74] |
| agrA | agrA 1 agrA 2 | CAACCACAAGTTGTTAAAGCAG TCGTTGTTTGCTTCAGTGATTC | 173 | 57.6 60.3 | 40.9 40.9 | (This work) |
| RNAIII | RNAIII 1 RNAIII 2 | CATGGTTATTAAGTTGGGATGGC GAAGGAGTGATTTCAATGGCACA | 188 | 58.31 60.02 | 43.48 43.48 | [75,76] |
| icaA | icaA 1 icaA 2 | GAGGTAAAGCCAACGCACTC CCTGTAACCGCACCAAGTTT | 151 | 59.70 59.18 | 55 50 | [77,78] |
| icaD | icaD 1 icaD 2 | ACCCAACGCTAAAATCATCG GCGAAAATGCCCATAGTTTC | 211 | 56.99 56.16 | 45 45 | [77,78] |
| aur | Aur 1 Aur 2 | ACCGTGTGTTAATTCGTGTGCTA ATGGTCGCACATTCACAAGTTT | 65 | 61.33 59.90 | 43.49 40.91 | [79] |
| SarA | SarA 1 SarA 2 | GTAATGAGCATGATGAAAGAACTGT CGTTGTTTGCTTCAGTGATTCG | 111 | 58.44 59.53 | 36 45.45 | [80] |
| rbf | Rbf 1 Rbf 2 | AACCACCTAACTGATGTTATAC GACAACTTGACTGTTCTTATTC | 156 | 53.77 53.59 | 36.36 36.36 | [81] |
| RNAr | Nuc 1 Nuc 2 | AATATGGACGTGGCTTAGCGT TTGACCTGAATCAGCGTTGTCTT | 197 | 60.38 61.28 | 47.62 43.48 | (This work) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, A.; Stashenko, E.E.; Sáez, R.T.; Zafra, G.; Ortiz, C. Effect of Essential Oil from Lippia origanoides on the Transcriptional Expression of Genes Related to Quorum Sensing, Biofilm Formation, and Virulence of Escherichia coli and Staphylococcus aureus. Antibiotics 2023, 12, 845. https://doi.org/10.3390/antibiotics12050845
Martínez A, Stashenko EE, Sáez RT, Zafra G, Ortiz C. Effect of Essential Oil from Lippia origanoides on the Transcriptional Expression of Genes Related to Quorum Sensing, Biofilm Formation, and Virulence of Escherichia coli and Staphylococcus aureus. Antibiotics. 2023; 12(5):845. https://doi.org/10.3390/antibiotics12050845
Chicago/Turabian StyleMartínez, Andrés, Elena E. Stashenko, Rodrigo Torres Sáez, German Zafra, and Claudia Ortiz. 2023. "Effect of Essential Oil from Lippia origanoides on the Transcriptional Expression of Genes Related to Quorum Sensing, Biofilm Formation, and Virulence of Escherichia coli and Staphylococcus aureus" Antibiotics 12, no. 5: 845. https://doi.org/10.3390/antibiotics12050845