

Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru

1
Health Science Research Laboratory, Universidad Señor de Sipán, Chiclayo 14001, Peru
2
Faculty of Health Sciences, Universidad Señor de Sipán, Chiclayo 14001, Peru
*
Author to whom correspondence should be addressed.
Antibiotics 2023, 12(4), 779; https://doi.org/10.3390/antibiotics12040779
Submission received: 8 February 2023
/
Revised: 10 April 2023
/
Accepted: 16 April 2023
/
Published: 19 April 2023
(This article belongs to the Special Issue Antimicrobial and Anti-infective Activity of Natural Products)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Due to the emergence of microorganisms resistant to antibiotics and the failure of antibiotic therapies, there is an urgent need to search for new therapeutic options, as well as new molecules with antimicrobial potential. The objective of the present study was to evaluate the in vitro antibacterial activity of Apis mellifera venom collected in the beekeeping areas of the city of Lambayeque in northern Peru against Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus. Bee venom extraction was performed by electrical impulses and separated using the Amicon ultra centrifugal filter. Subsequently, the fractions were quantified by spectrometric 280 nm and evaluated under denaturant conditions in SDS-PAGE. The fractions were pitted against Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 29213, and Pseudomonas aeruginosa ATCC 27853. A purified fraction (PF) of the venom of A. mellifera and three low molecular weight bands of 7 KDa, 6 KDa, and 5 KDa were identified that showed activity against E. coli with a MIC of 6.88 µg/mL, while for P. aeruginosa and S. aureus, it did not present a MIC. No hemolytic activity at a concentration lower than 15.6 µg/mL and no antioxidant activity. The venom of A. mellifera contains a potential presence of peptides and a predilection of antibacterial activity against E. coli.
1. Introduction
World Health Organization (WHO) published the first global surveillance report on antibiotic resistance, showing that five out of six regions had more than 50% resistance to third-generation cephalosporins and fluoroquinolones in Escherichia coli and methicillin resistance in Staphylococcus aureus in hospital settings [1].
S. aureus is a human commensal that can cause systemic infections in the host; this requires evading the immune response and the ability to proliferate in different niches in the host; currently, the infection by staphylococci in the face of immune mediators and the disease is not well known [2]. However, the main agent of bacteremia and infective endocarditis (IE), as well as osteoarticular, skin, and soft tissue infections, pleuropulmonary infections [3], and even the appearance of methicillin-resistant S. aureus (MRSA), which is a therapeutic problem in patients [4]. Staphylococcal infection has also been reported from hosts or carriers of asymptomatic nasopharyngeal bacteria, even with certain risk factors such as passive smoking and a large family [5]. The results of certain studies have determined that S. aureus has generated resistance against ampicillin, penicillin, rifampicin, clindamycin, oxacillin, and erythromycin [6]. Variable susceptibility to levofloxacin, ciprofloxacin, gentamicin, tetracycline, and sulfamethoxazole-trimethoprim has also been shown [7], and patterns have also been shown regarding the mecA, rpoB, blaZ, ermB, tetM, and nuc genes [6,8]. This resistance has been acquired through different mechanisms, the most frequent being reduced membrane permeability, excessive production of β-lactamase, and acquiring resistance genes or gene mutations [9].
Likewise, Pseudomonas aeruginosa can cause nosocomial outbreaks related to its resistance and virulence properties [10], being a producer of β-lactamases and multiresistant to a wide range of antimicrobials such as penicillin, cephalosporin, cephamycin, and carbapenem [11]. In addition, 35 resistomes (antimicrobial resistance genes) have been identified that confer resistance to 18 different antibiotics (including four classes of beta-lactams) and 214 virulence factor genes [12], in addition to the susceptibility of P. aeruginosa to carbapenems, piperacillin-tazobactam, and amikacin has undergone alterations before and during COVID-19 [13]. There are phenotypic studies that P. aeruginosa, as producers of extended-spectrum beta-lactamase (ESBL) and metallo-beta-lactamases (MBL), also present genes associated with biofilm formation and virulence, such as toxA and lasB [14]. Even using polymerase chain reaction (PCR) techniques and molecular markers such as Random Amplified Polymorphic DNA (RAPD), they have identified strains resistant to imipenem, Ticarcillin + Clavulanate, Piperacillin, and Ticarcillin + Clavulanate, these strains being isolated from swimming pools [15].
E. coli is responsible for a large number of virulent variants associated with human diseases, such as urinary tract infection (UTI) with a resistance rate of >55% to first to fourth-generation cephalosporins [16], neonatal and traveler’s diarrhea [17], and multiresistant isolates (MDRs) the most prevalent genes being CTX-M-1, followed by NDM-1 for Betalactamases and the genes ermB and aac(6′)-Ib for resistance to macrolides and aminoglycosides [18]. E. coli is frequently discharged into the environment through feces, including wastewater, and is considered an indicator of fecal contamination. Many strains can carry resistance genes [19]. According to isolates, they have usually been reported to be sensitive to netilmicin, gentamicin, chloramphenicol, pipemidic acid, nalidixic acid, ciprofloxacin, amoxicillin/clavulanic acid, and nitrofurantoin, as well as increased susceptibility to cefotaxime, ceftriaxone, and aztreonam [20]. Uncomplicated UTI isolates have been found to have a higher susceptibility than complicated UTI isolates to amoxicillin, amoxicillin/clavulanic acid, and ciprofloxacin [21].
Antimicrobial resistance has been reported in gram-negative and gram-positive bacteria, with reports of up to 96.2% for Pseudomonas spp. and 66.7% for E. coli [22], with isolates reported in heart disease intensive care units [23] and in bloodstream infections [24] antimicrobial-resistant bacteria are also considered the most frequent uropathogens [25]. There are reports of MRSA in up to 50% of patients [26], with resistance profiles for cefoxitin, chloramphenicol, clindamycin, and gentamicin [27].
The failure of antibiotics, including the latest generation, to counteract superbugs highlights the need to search for new molecules with antimicrobial potential to control the global problem of antimicrobial resistance. One group of these new molecules are peptides that are antimicrobial (AMP) and are promising molecules for combating antimicrobial resistance (AMR) [28]. AMP has been found the most in the venoms of different organisms, such as scorpions [29,30,31], snakes [32], spiders [33], and bees [34], among other venoms.
The composition of bee venom is very variable, having Peptides: Melitin (the main component of the venom), Apamin, Mast Cell-Degranulating Peptide (MCD), Tertiapin, Secapin, and Its Isoforms, Adolapin, Procamine, and Minimine; Polypeptides: Api m 6, Cardiopep, Icarapin, and Major Royal Jelly Proteins; Enzymes: Phospholipase A2 (PLA2), Hyaluronidase, Acid Phosphatase, and Dipeptidylpeptidase IV; Serine Proteases [35].
Bees are insects found on all continents, many of these species have yet to be described and are an exciting source for the study and search for new molecules with antimicrobial properties. There are experimental and clinical reports on Apis mellifera venom and its anti-inflammatory, antimicrobial, and anticancer effects; the components present in the venom, such as proteins, vary from a summer season compared to a winter season [36,37,38], in addition, have shown different therapeutic properties against oxidative stress induced by beta-amyloid [39,40,41]. For Parkinson’s disease, the neuroprotective potential of bee venom against oxidative stress induced by rotenone (pesticide) has been demonstrated in a mouse model, including preventing the decrease in dopamine and also restoring locomotor activity in mice [42,43]. For Lyme disease, the melittin present in the venom showed in vitro antibacterial effects against the causative agent Borrelia burgdorferi [44] and even had significant antibacterial effects against E. coli, S. aureus, and Salmonella typhyimurium [45]. Melittin also exhibited antibacterial activity against MRSA strains [46], with antimicrobial potential against agents that cause dental caries, with antifungal capacity including suppression of biofilm formation [47,48]. Its significant antiviral potential has also been demonstrated in in vitro and in vivo assays on different enveloped (Influenza A) and non-enveloped (enterovirus-71) viruses [49]. In addition, phospholipase A2 (PLA2) can also block the replication of the virus, being shown to be responsible for the inhibition of HIV replication [50]. The present study aimed to evaluate the in vitro antibacterial activity of Apis mellifera venom collected in the beekeeping areas of the city of Lambayeque in northern Peru against Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus.
2. Results
As seen in Figure 1, 15% SDS-PAGE-Tricine of the purified fraction (PF) of crude venom from A. mellifera yielded 3 low molecular weight bands, i.e., 7 kDa, 6 kDa, and 5 kDa.
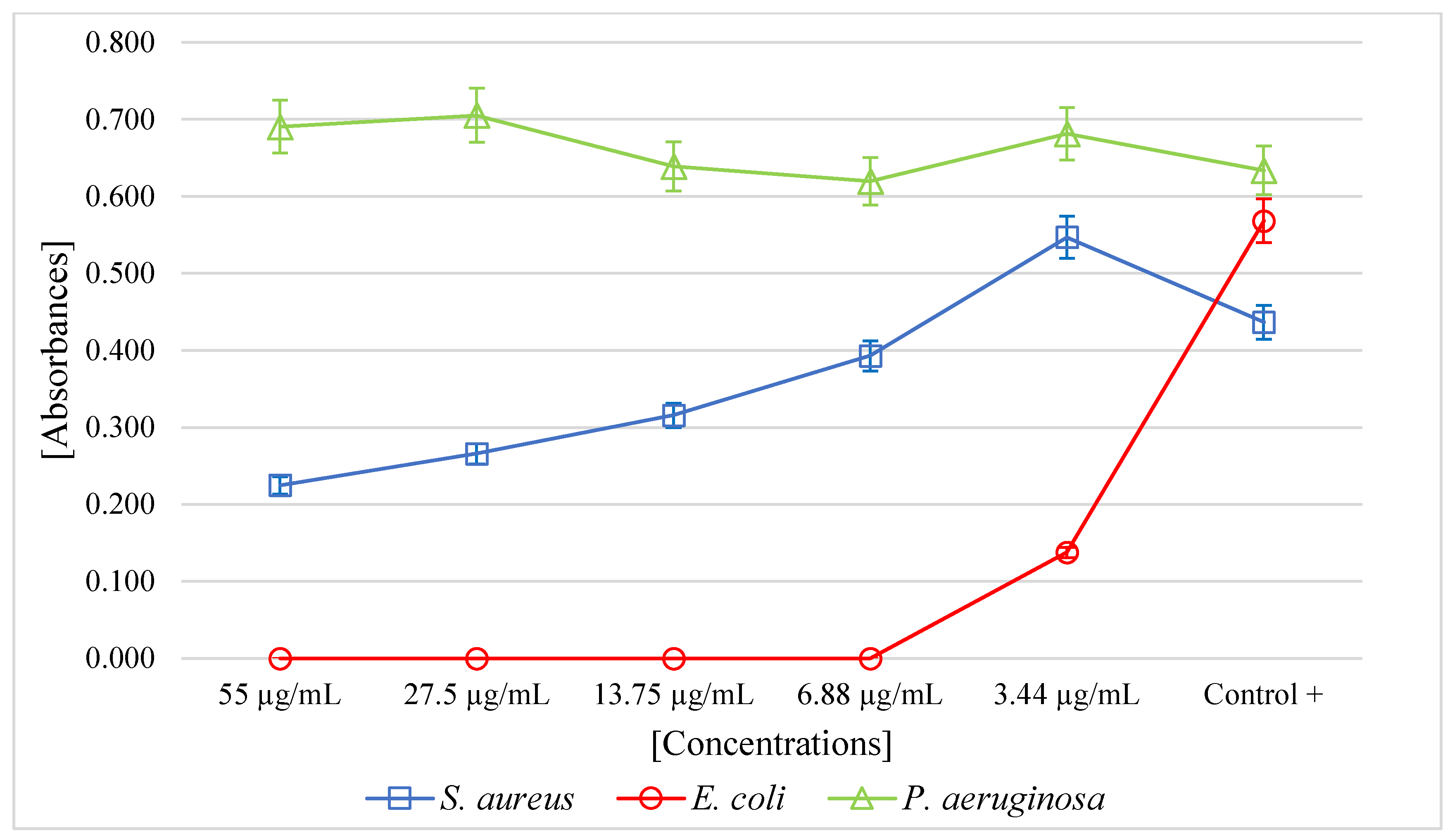
The PF of A. mellifera venom had a minimum inhibitory concentration (MIC) of 6.88 µg/mL (p < 0.05) for E. coli. For the S. aureus strain, at higher concentrations, the venom exhibited antibacterial activity. For P. aeruginosa, no antibacterial activity was observed (Figure 2).
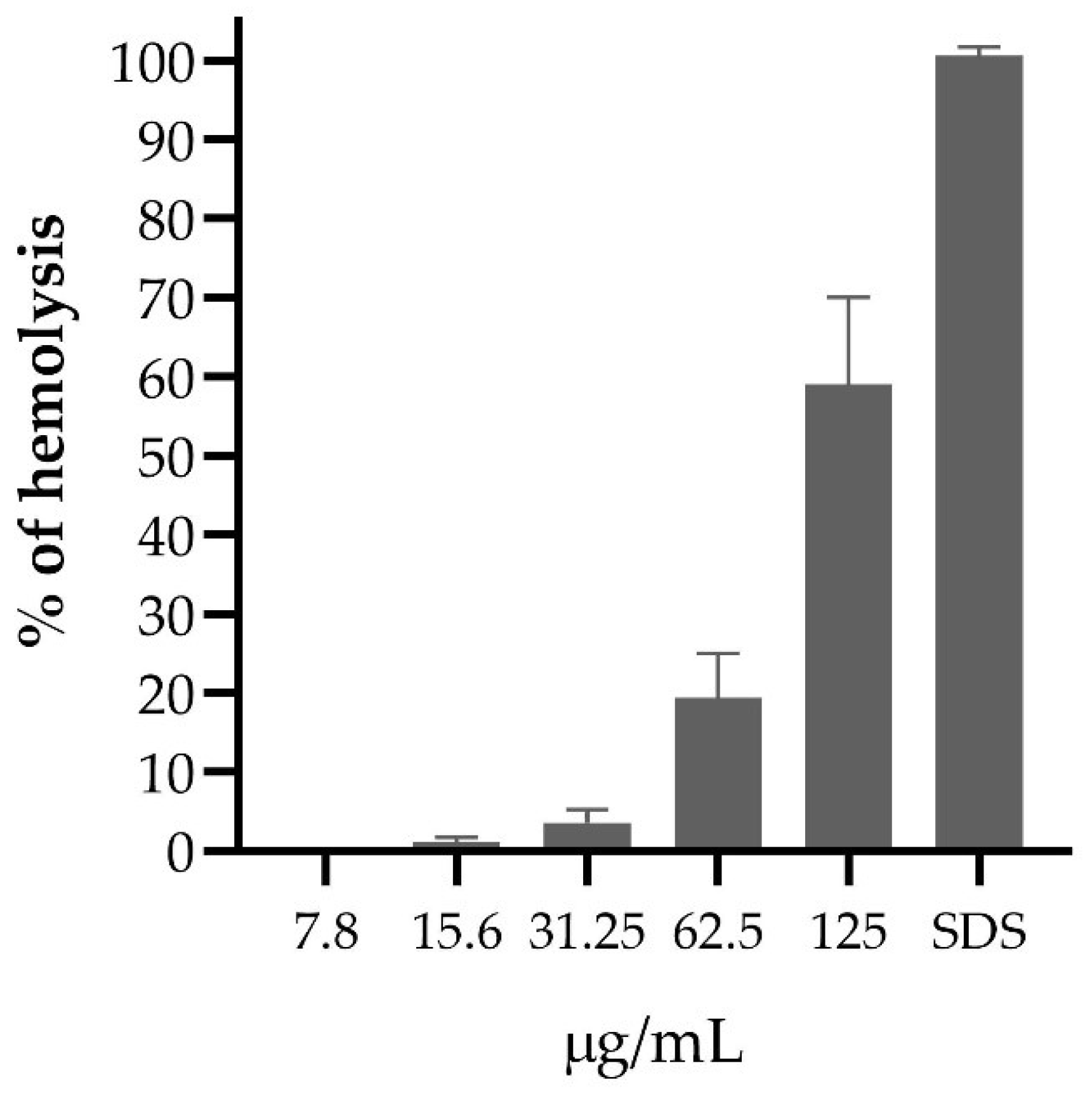
Concentrations of the PF of A. mellifera venom above 125 µg/mL in erythrocyte suspension produced more than 50% hemolysis. At a concentration of 31.25 µg /mL, less than 10% hemolysis occurred, but at 7.8 µg /mL PF, no erythrocyte lysis was evidenced (Figure 3).
The PF of the A. mellifera venom showed no antioxidant activity was observed at the concentrations evaluated.
3. Discussion
The purified fraction (PF) of the A. mellifera venom revealed the presence of peptides using the SDS-PAGE-Tricine technique and found three peptides of 7 kDa, 6 kDa, and 5 kDa. The chemical composition of the venom of A. mellifera is highly variable; such as melittin (3 kDa), apamin (2 kDa), and cecropin (4 kDa), enzymes, such as phospholipase A2 (19 kDa) and hyaluronidase (38 kDa), biologically active amines, such as histamine and epinephrine, as well as peptides not reported [39,51,52] and this suggests that many of these components may contribute their anti-inflammatory, antifungal, antiviral, healing and analgesic properties [36,53,54,55].
Our results were coherent with other studies; the PF of A. mellifera venom collected from Íllimo showed antibacterial activity against E. coli, but the same was not observed for P. aeruginosa despite being a gram-negative bacterium. For S. aureus, as the concentration of the venom fraction increased, bacterial growth was affected. Interestingly, the results of other studies of the antibacterial activity of the crude venom showed highly variable MICs against E. coli and for S. aureus [56], including also demonstrating the inhibitory effect through a viability assay at a temperature of 25 °C against E. coli and P. putida, causing membrane permeability and loss of ATP, showing no effect against P. fluorescens [57], as well as the crude bee venom extract in a region of Egypt significantly inhibited the growth of E. coli ATCC8739 and S. aureus ATCC 6538P [58], in addition, the action of crude bee venom from Iran demonstrated inhibition through the Kirby–Bauer method against E. coli and S. aureus, but not against P. aeruginosa [59].
Other studies have shown the existence of antimicrobial peptides in the venom; it follows that the interaction against the cell envelope of the bacteria is due to the attraction between the positively charged venom peptides and the phospholipids, causing a rupture or instability of the venom membrane, in addition to forming pores; however, this mechanism requires a certain concentration threshold [60]. Direct insertion of melittin leads to pore formation, whereas the parallel conformation is inactive and prevents other melittin molecules from being inserted, thus, preventing pore formation [61]. However, melittin has a molecular weight of 3 KDa; in our study, we found three peptides in the range of 5 KDa to 7 KDa; this finding demonstrates that melittin is not the only peptide present in bee venom with antibacterial activity.
The PF of A. mellifera venom at a concentration lower than 15.6 µg/mL demonstrated low hemolytic activity. Few studies have revealed the hemolytic activity of bee venom in Peru; on the contrary, in other latitudes, they have revealed that melittin has not presented significant hemolytic activity below a concentration of 0.25 µg/mL [62], the hemolytic action was also demonstrated against erythrocytes of different species, with variable sensitivity to bee venom pools, with sheep erythrocytes being the most resistant to hemolytic action compared to equine erythrocytes, including humans erythrocytes showed good resistance to hemolytic action, it follows that hemolysis can be increased by the action of phospholipase 2 (PLA2) after the action of melittin [63].
The PF of the venom of A. mellifera has not shown antioxidant capacity, but in other studies, they demonstrated antioxidant capacity. Curiously, they worked with the total venom or apitoxin, having the capacity to inhibit the free radical DPPH (2,2-diphenyl-1-picrylhydrazine) between 60% and 75% of antioxidant activity [64]. In the same sense, demonstrated with the venom of A. mellifera syriaca eliminating DPPH radicals between 50 to 65% [65]. When analyzing the venom of four bee species, A. dorsata, A. mellifera, A. florea, and A. cerena, they showed that A. dorsata contained the highest amount of melittin; they also revealed that the extract of A. dorsata had the highest antioxidant activity from the DPPH and ABTS (3-ethylbenzothiazoline-6-sulfonic acid) assays, including melittin alone, revealed very poor antioxidant activity among all bee venom extracts [66], this suggests that in our study, of the peptides present in the venom PF, melittin was not present.
The bioactive components present in the venom have generated much interest in medicine through the different species of the Apis genus, and their application in in vitro antimicrobial activity [67], their cytotoxic action against cancer cells [68], even the synergistic effect of the venom with some antibiotics such as Cephotax, Cefepime, and Tavanic has been revealed [69]. Through Transmission Electron Microscopy, the deformation of the cell wall was appreciated, resulting in the destruction of the cell wall, changes in the permeability of the membrane, leakage of cell content, inactivation of metabolic activity, and finally, cell death [57], as the inhibitory effect on F1-F0—ATPase has also been demonstrated [70].
4. Materials and Methods
4.1. Bee Venom Samples
The venom was obtained from Africanized bees A. mellifera (Linnaeus, 1758) in hives from the Cruz Verde town center of the Ilimo District located at latitude 6°28′26″ and longitude 79°50′34″ (Lambayeque); an electrical impulse of 3 volts with an electrical intensity of 0.004 A was passed through a collecting box (beeWhisper 6.0; Model 2020), without harming the specimens (Figure 4a,b) [71].
The venom was collected on a glass plate and allowed to dry; it was then transferred into a 50-mL Falcon tube (Figure 4c,d). Then, the bee venom was resuspended in sterile deionized water and centrifuged at 10,000× g at 4 °C for 10 min to remove insoluble materials. The supernatant was collected and stored in 2 mL microtubes at −20 °C.
4.2. Fraction Concentration and Electrophoresis
The fractions were collected and quantified via absorbance at 280 nm (Navi UV/vis Nano spectrophotometer, Seongnam-si, Republic of Korea) using the formula [mg/mL] = (1.56 × Abs 280 nm) − (0.76 × Abs 260 nm) [72].
Crude fractions of bee venom were collected and concentrated using the Amicon ultra centrifugal filter (Merck Millipore, Cork, Ireland) with Cutoff from 3 kDa to 100 kDa [73], quantified via absorbance, and 50 µg protein from the FP were evaluated on a gel Tricine-SDS-PAGE (15%) under denaturing conditions [74] with a voltage of 100 volts and stained with Coomassie blue [75].
4.3. Antimicrobial Activity Test
The MIC values of the fraction were determined using the broth microdilution method in 96-well plates [76] against the strains of E. coli ATCC 25922, S. aureus ATCC 29213, and P. aeruginosa ATCC 27853; 50 µL of bacterial solution containing 5 × 104 CFU/mL was placed in each well, then 50 µL of different concentrations of the fraction (55 µg/mL to 3.44 µg/mL) were added and incubated at 37 °C for 24 h. The positive control was broth plus inoculum, and the negative control was only broth. Growth of the positive control was determined by a growth button of ≥2 mm or defined turbidity. Finally, the plates were read by absorbance at 630 nm (SmartReader 96—Accuris) to determine the minimum inhibitory concentration (MIC). All assays were performed in triplicate.
4.4. Evaluation of Hemolytic Activity
The hemolytic test was evaluated following the protocol described by Oddo [77], red blood cells washed with PBS (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4) and resuspended at a concentration of 0.5% and incubated for 1 h with different concentrations of the fractions and then centrifuged at 10,000× g for 10 min, 60 µL of supernatant was transferred to a 96-well polypropylene plate, and the absorbance was read at 405 nm. The results were normalized with the positive controls of hemolysis (0.25% SDS) and negative controls (PBS). Assays were performed in triplicate.
4.5. Evaluation of Antioxidant Activity
20 μL of different concentrations of the fraction (55 µg/mL to 3.44 µg/mL) were added with 380 μL of ABTS radical in ethanol, incubated at room temperature protected from light for 30 min, then the absorbance of the mixture was measured at 734 nm [66]. To calculate the % decoloration, the following equation was used: % decoloration = [(C − S)/C] × 100, where C is the absorbance of the control, and S is the absorbance of the problem sample. Trolox was used as a positive control. The experiments were done in triplicate.
4.6. Statistic Analysis
The MegaStat add-in for Excel was used to determine the antibacterial activity of the purified fraction of Apis mellifera venom. Analysis of variance (ANOVA) was performed at a significance level of 5%.
5. Conclusions
In summary, A. mellifera bee venom contained peptides with weights of 7 kDa, 6 kDa, and 5 kDa and exhibited antibacterial activity against E. coli ATCC 25922 at a concentration of 6.88 µg/mL.
Author Contributions
Writing—preparation of the original draft, methodology, O.P.-D.; formal analysis, A.O.E.-C.; data curation, E.L.-L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Universidad Señor de Sipán with RD No. 074-2020/PD-USS. The funders had no role in the collection, analysis, and interpretation of the data.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data related to the manuscript were available in the main manuscript.
Acknowledgments
We are grateful for the assistance of Señor de Sipán University for the technical, translation, and logistical support provided for this research. We thank Luis Orlando Sánchez Suclupe, for his support with the hives for the extraction of bee venom, Karl Weiss Cruz Verde—Íllimo Bee-keeping Association. We thank the Thematic Network for the development of antiviral and antimicrobial peptides for multiresistant strains (RED BUDEPAV-AM) and for participating as an exhibitor at the international symposium on antimicrobial and antiviral peptides.
Conflicts of Interest
The authors declare that there are no conflict of interest with respect to the publication of this article.
References
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; ISBN 978-92-4-156474-8. [Google Scholar]
- Pollitt, E.J.G.; Szkuta, P.T.; Burns, N.; Foster, S.J. Staphylococcus Aureus Infection Dynamics. PLoS Pathog. 2018, 14, e1007112. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Safdari, H.; Aryan, E.; Sadeghian, H.; Shams, S.F.; Aganj, M. Frequency of Methicillin-Resistant Staphylococcus Aureus (MRSA) in Nose and Cellular Phone of Medical and Non-Medical Personnel of Emergency Departments of Ghaem Hospital in Mashhad City. Clin. Epidemiol. Glob. Health 2020, 8, 1043–1046. [Google Scholar] [CrossRef]
- Belayhun, C.; Tilahun, M.; Seid, A.; Shibabaw, A.; Sharew, B.; Belete, M.A.; Demsiss, W. Asymptomatic Nasopharyngeal Bacterial Carriage, Multi-Drug Resistance Pattern and Associated Factors among Primary School Children at Debre Berhan Town, North Shewa, Ethiopia. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 9. [Google Scholar] [CrossRef]
- Akanbi, O.E.; Njom, H.A.; Fri, J.; Otigbu, A.C.; Clarke, A.M. Antimicrobial Susceptibility of Staphylococcus Aureus Isolated from Recreational Waters and Beach Sand in Eastern Cape Province of South Africa. Int. J. Environ. Res. Public Health 2017, 14, 1001. [Google Scholar] [CrossRef]
- Qodrati, M.; SeyedAlinaghi, S.; Dehghan Manshadi, S.A.; Abdollahi, A.; Dadras, O. Antimicrobial Susceptibility Testing of Staphylococcus Aureus Isolates from Patients at a Tertiary Hospital in Tehran, Iran, 2018–2019. Eur. J. Med. Res. 2022, 27, 152. [Google Scholar] [CrossRef]
- Nikbakht, M.; Ahangarzadeh Rezaee, M.; Hasani, A.; Nahaei, M.R.; Sadeghi, J.; Jedari Seifi, S. Molecular Characterization and Antimicrobial Susceptibility Patterns of Methicillin-Resistant Staphylococcus Aureus Isolates in Tabriz, Northwest of Iran. Arch. Pediatr. Infect. Dis. 2017, in press. [CrossRef]
- Guo, Y.; Song, G.; Sun, M.; Wang, J.; Wang, Y. Prevalence and Therapies of Antibiotic-Resistance in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2020, 10, 107. [Google Scholar] [CrossRef]
- Hu, P.; Chen, J.; Chen, Y.; Zhou, T.; Xu, X.; Pei, X. Molecular Epidemiology, Resistance, and Virulence Properties of Pseudomonas aeruginosa Cross-Colonization Clonal Isolates in the Non-Outbreak Setting. Infect. Genet. Evol. 2017, 55, 288–296. [Google Scholar] [CrossRef]
- Nasser, M.; Gayen, S.; Kharat, A.S. Prevalence of β-Lactamase and Antibiotic-Resistant Pseudomonas aeruginosa in the Arab Region. J. Glob. Antimicrob. Resist. 2020, 22, 152–160. [Google Scholar] [CrossRef]
- Hoque, M.N.; Jahan, M.I.; Hossain, M.A.; Sultana, M. Genomic Diversity and Molecular Epidemiology of a Multidrug-Resistant Pseudomonas aeruginosa DMC30b Isolated from a Hospitalized Burn Patient in Bangladesh. J. Glob. Antimicrob. Resist. 2022, 31, 110–118. [Google Scholar] [CrossRef]
- Coșeriu, R.L.; Vintilă, C.; Mare, A.D.; Ciurea, C.N.; Togănel, R.O.; Cighir, A.; Simion, A.; Man, A. Epidemiology, Evolution of Antimicrobial Profile and Genomic Fingerprints of Pseudomonas aeruginosa before and during COVID-19: Transition from Resistance to Susceptibility. Life 2022, 12, 2049. [Google Scholar] [CrossRef]
- Asadpour, L. Antimicrobial Resistance, Biofilm-Forming Ability and Virulence Potential of Pseudomonas aeruginosa Isolated from Burn Patients in Northern Iran. J. Glob. Antimicrob. Resist. 2018, 13, 214–220. [Google Scholar] [CrossRef]
- Schiavano, G.F.; Carloni, E.; Andreoni, F.; Magi, S.; Chironna, M.; Brandi, G.; Amagliani, G. Prevalence and Antibiotic Resistance of Pseudomonas aeruginosa in Water Samples in Central Italy and Molecular Characterization of OprD in Imipenem Resistant Isolates. PLoS ONE 2017, 12, e0189172. [Google Scholar] [CrossRef]
- Alotaibi, B.S.; Tantry, B.A.; Farhana, A.; Alammar, M.A.; Shah, N.N.; Mohammed, A.H.; Wani, F.; Bandy, A. Resistance Pattern in Mostly Gram-Negative Bacteria Causing Urinary Tract Infections. Infect. Disord. Drug Targets 2023, 23, 56–64. [Google Scholar] [CrossRef]
- Umpiérrez, A.; Ernst, D.; Fernández, M.; Oliver, M.; Casaux, M.L.; Caffarena, R.D.; Schild, C.; Giannitti, F.; Fraga, M.; Zunino, P. Virulence Genes of Escherichia coli in Diarrheic and Healthy Calves. Rev. Argent. Microbiol. 2021, 53, 34–38. [Google Scholar] [CrossRef]
- Sarjana Safain, K.; Bhuyan, G.S.; Hassan Hasib, S.; Islam, M.S.; Mahmud-Un-Nabi, M.A.; Sultana, R.; Tasnim, S.; Noor, F.A.; Sarker, S.K.; Islam, M.T.; et al. Genotypic and Phenotypic Profiles of Antibiotic-resistant Bacteria Isolated from Hospitalised Patients in Bangladesh. Trop. Med. Int. Health 2021, 26, 720–729. [Google Scholar] [CrossRef]
- Jang, J.; Hur, H.-G.; Sadowsky, M.J.; Byappanahalli, M.N.; Yan, T.; Ishii, S. Environmental Escherichia coli.: Ecology and Public Health Implications—A Review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef]
- Edmond, T.; Yehouenou, L.C.; Malick, Z.F.; Arsene, K.A.; Rene, K.K.; Diouara, A.A.M.; Tonde, I.; Bankole, H.S.; Wilfried, B.K.; Marius, E.A.; et al. Antimicrobial Susceptibility of Community Acquired Escherichia coli in Urinary Tract Infections (UTI) in Benin for Eleven Years (2005–2015). Am. J. Infect. Dis. 2017, 13, 21–27. [Google Scholar] [CrossRef]
- Grados, M.C.; Thuissard, I.J.; Alós, J.-I. Stratification by Demographic and Clinical Data of the Antibiotic Susceptibility of Escherichia Coli from Urinary Tract Infections of the Community. Atención Primaria 2019, 51, 494–498. [Google Scholar] [CrossRef]
- Amanati, A.; Sajedianfard, S.; Khajeh, S.; Ghasempour, S.; Mehrangiz, S.; Nematolahi, S.; Shahhosein, Z. Bloodstream Infections in Adult Patients with Malignancy, Epidemiology, Microbiology, and Risk Factors Associated with Mortality and Multi-Drug Resistance. BMC Infect. Dis. 2021, 21, 636. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Mahzari, M.; Banar, M.; Pourakbari, B.; Haghi Ashtiani, M.T.; Mohammadi, M.; Keshavarz Valian, S.; Mamishi, S. Antimicrobial Resistance Patterns of Gram-Negative Bacteria Isolated from Bloodstream Infections in an Iranian Referral Paediatric Hospital: A 5.5-Year Study. J. Glob. Antimicrob. Resist. 2017, 11, 17–22. [Google Scholar] [CrossRef]
- Shi, N.; Kang, J.; Wang, S.; Song, Y.; Yin, D.; Li, X.; Guo, Q.; Duan, J.; Zhang, S. Bacteriological Profile and Antimicrobial Susceptibility Patterns of Gram-Negative Bloodstream Infection and Risk Factors Associated with Mortality and Drug Resistance: A Retrospective Study from Shanxi, China. Infect. Drug Resist. 2022, 15, 3561–3578. [Google Scholar] [CrossRef]
- Esposito, S.; Maglietta, G.; Di Costanzo, M.; Ceccoli, M.; Vergine, G.; La Scola, C.; Malaventura, C.; Falcioni, A.; Iacono, A.; Crisafi, A.; et al. Retrospective 8-Year Study on the Antibiotic Resistance of Uropathogens in Children Hospitalised for Urinary Tract Infection in the Emilia-Romagna Region, Italy. Antibiotics 2021, 10, 1207. [Google Scholar] [CrossRef]
- Boschetti, G.; Sgarabotto, D.; Meloni, M.; Bruseghin, M.; Whisstock, C.; Marin, M.; Ninkovic, S.; Pinfi, M.; Brocco, E. Antimicrobial Resistance Patterns in Diabetic Foot Infections, an Epidemiological Study in Northeastern Italy. Antibiotics 2021, 10, 1241. [Google Scholar] [CrossRef]
- Obakiro, S.B.; Kiyimba, K.; Paasi, G.; Napyo, A.; Anthierens, S.; Waako, P.; Royen, P.V.; Iramiot, J.S.; Goossens, H.; Kostyanev, T. Prevalence of Antibiotic-Resistant Bacteria among Patients in Two Tertiary Hospitals in Eastern Uganda. J. Glob. Antimicrob. Resist. 2021, 25, 82–86. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Tawfik, M.M.; Bertelsen, M.; Abdel-Rahman, M.A.; Strong, P.N.; Miller, K. Scorpion Venom Antimicrobial Peptides Induce Siderophore Biosynthesis and Oxidative Stress Responses in Escherichia coli. mSphere 2021, 6, e00267-21. [Google Scholar] [CrossRef]
- Pérez-Delgado, O.; Rincon-Cortés, C.A.; Vega-Castro, N.A.; Reyes-Montaño, E.A.; Gómez-Garzón, M. Purificación Parcial de Péptidos Del Veneno de Escorpión Hadruroides Charcasus (Karsch, 1879) Con Actividad Antimicrobiana. Bionat. Lat. Am. J. Biotechnol. Life Sci. 2021, 6, 1917–1923. [Google Scholar] [CrossRef]
- Zhao, Z.; Ma, Y.; Dai, C.; Zhao, R.; Li, S.; Wu, Y.; Cao, Z.; Li, W. Imcroporin, a New Cationic Antimicrobial Peptide from the Venom of the Scorpion Isometrus maculates. Antimicrob. Agents Chemother. 2009, 53, 3472–3477. [Google Scholar] [CrossRef]
- de Barros, E.; Gonçalves, R.M.; Cardoso, M.H.; Santos, N.C.; Franco, O.L.; Cândido, E.S. Snake Venom Cathelicidins as Natural Antimicrobial Peptides. Front. Pharmacol. 2019, 10, 1415. [Google Scholar] [CrossRef]
- Lee, B.; Shin, M.K.; Yoo, J.S.; Jang, W.; Sung, J.-S. Identifying Novel Antimicrobial Peptides from Venom Gland of Spider Pardosa Astrigera by Deep Multi-Task Learning. Front. Microbiol. 2022, 13, 971503. [Google Scholar] [CrossRef]
- Ko, S.J.; Park, E.; Asandei, A.; Choi, J.-Y.; Lee, S.-C.; Seo, C.H.; Luchian, T.; Park, Y. Bee Venom-Derived Antimicrobial Peptide Melectin Has Broad-Spectrum Potency, Cell Selectivity, and Salt-Resistant Properties. Sci. Rep. 2020, 10, 10145. [Google Scholar] [CrossRef]
- Abd El-Wahed, A.A.; Khalifa, S.A.M.; Sheikh, B.Y.; Farag, M.A.; Saeed, A.; Larik, F.A.; Koca-Caliskan, U.; AlAjmi, M.F.; Hassan, M.; Wahabi, H.A.; et al. Bee Venom Composition: From Chemistry to Biological Activity. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; Volume 60, pp. 459–484. ISBN 978-0-444-64181-6. [Google Scholar]
- Amin, M.A.; Abdel-Raheem, I.T. Accelerated Wound Healing and Anti-Inflammatory Effects of Physically Cross Linked Polyvinyl Alcohol–Chitosan Hydrogel Containing Honey Bee Venom in Diabetic Rats. Arch. Pharm. Res. 2014, 37, 1016–1031. [Google Scholar] [CrossRef]
- Ward, R.; Coffey, M.; Kavanagh, K. Proteomic Analysis of Summer and Winter Apis mellifera Workers Shows Reduced Protein Abundance in Winter Samples. J. Insect Physiol. 2022, 139, 104397. [Google Scholar] [CrossRef]
- Kwon, N.-Y.; Sung, S.-H.; Sung, H.-K.; Park, J.-K. Anticancer Activity of Bee Venom Components against Breast Cancer. Toxins 2022, 14, 460. [Google Scholar] [CrossRef]
- Kim, H.; Park, S.-Y.; Lee, G. Potential Therapeutic Applications of Bee Venom on Skin Disease and Its Mechanisms: A Literature Review. Toxins 2019, 11, 374. [Google Scholar] [CrossRef]
- Hegazi, A.; Abdou, A.M.; El-Moez, S.I.A.; Allah, F.A. Evaluation of the Antibacterial Activity of Bee Venom from Different Sources. World Appl. Sci. J. 2014, 30, 266–270. [Google Scholar]
- Nguyen, C.D.; Yoo, J.; Hwang, S.-Y.; Cho, S.-Y.; Kim, M.; Jang, H.; No, K.O.; Shin, J.C.; Kim, J.-H.; Lee, G. Bee Venom Activates the Nrf2/HO-1 and TrkB/CREB/BDNF Pathways in Neuronal Cell Responses against Oxidative Stress Induced by Aβ1–42. Int. J. Mol. Sci. 2022, 23, 1193. [Google Scholar] [CrossRef]
- Tanner, C.M.; Kamel, F.; Ross, G.W.; Hoppin, J.A.; Goldman, S.M.; Korell, M.; Marras, C.; Bhudhikanok, G.S.; Kasten, M.; Chade, A.R.; et al. Rotenone, Paraquat, and Parkinson’s Disease. Environ. Health Perspect. 2011, 119, 866–872. [Google Scholar] [CrossRef]
- Khalil, W.K.B.; Assaf, N.; ElShebiney, S.A.; Salem, N.A. Neuroprotective Effects of Bee Venom Acupuncture Therapy against Rotenone-Induced Oxidative Stress and Apoptosis. Neurochem. Int. 2015, 80, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Socarras, K.; Theophilus, P.; Torres, J.; Gupta, K.; Sapi, E. Antimicrobial Activity of Bee Venom and Melittin against Borrelia Burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef]
- Zolfagharian, H.; Mohajeri, M.; Babaie, M. Bee Venom (Apis mellifera) an Effective Potential Alternative to Gentamicin for Specific Bacteria Strains: Bee Venom an Effective Potential for Bacteria. J. Pharmacopunct. 2016, 19, 225–230. [Google Scholar] [CrossRef]
- Han, S.; Kim, J.; Hong, I.; Woo, S.; Kim, S.; Jang, H.; Pak, S. Antibacterial Activity and Antibiotic-Enhancing Effects of Honeybee Venom against Methicillin-Resistant Staphylococcus Aureus. Molecules 2016, 21, 79. [Google Scholar] [CrossRef]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.C.; Cunha, W.R.; de Almeida, R.; Martins, C.H.G. Antimicrobial Activity of Apitoxin, Melittin and Phospholipase A2 of Honey Bee (Apis mellifera) Venom against Oral Pathogens. Ann. Acad. Bras. Ciênc. 2015, 87, 147–155. [Google Scholar] [CrossRef] [PubMed]
- El-Didamony, S.E.; Kalaba, M.H.; El-Fakharany, E.M.; Sultan, M.H.; Sharaf, M.H. Antifungal and Antibiofilm Activities of Bee Venom Loaded on Chitosan Nanoparticles: A Novel Approach for Combating Fungal Human Pathogens. World J. Microbiol. Biotechnol. 2022, 38, 244. [Google Scholar] [CrossRef]
- Uddin, M.B.; Lee, B.-H.; Nikapitiya, C.; Kim, J.-H.; Kim, T.-H.; Lee, H.-C.; Kim, C.G.; Lee, J.-S.; Kim, C.-J. Inhibitory Effects of Bee Venom and Its Components against Viruses In Vitro and In Vivo. J. Microbiol. 2016, 54, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Fenard, D.; Lambeau, G.; Valentin, E.; Lefebvre, J.C.; Lazdunski, M.; Doglio, A. Secreted Phospholipases A(2), a New Class of HIV Inhibitors That Block Virus Entry into Host Cells. J. Clin. Investig. 1999, 104, 611–618. [Google Scholar] [CrossRef]
- de Brito, J.C.M.; Bastos, E.M.A.F.; Heneine, L.G.D.; de Souza Figueiredo, K.C. Fractionation of Apis mellifera Venom by Means of Ultrafiltration: Removal of Phospholipase A2. Braz. J. Chem. Eng. 2018, 35, 229–236. [Google Scholar] [CrossRef]
- El-Seedi, H.; Abd El-Wahed, A.; Yosri, N.; Musharraf, S.G.; Chen, L.; Moustafa, M.; Zou, X.; Al-Mousawi, S.; Guo, Z.; Khatib, A.; et al. Antimicrobial Properties of Apis mellifera’s Bee Venom. Toxins 2020, 12, 451. [Google Scholar] [CrossRef] [PubMed]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A Venom-Derived Peptide with Promising Anti-Viral Properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Kurek-Górecka, A.; Komosinska-Vassev, K.; Rzepecka-Stojko, A.; Olczyk, P. Bee Venom in Wound Healing. Molecules 2020, 26, 148. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.-E.; Seo, B.-K.; Lee, J.-W.; Park, Y.-C.; Baek, Y.-H. Analgesic Effects of Diluted Bee Venom Acupuncture Mediated by δ-Opioid and A2-Adrenergic Receptors in Osteoarthritic Rats. Altern. Ther. Health Med. 2018, 24, 28–35. [Google Scholar] [PubMed]
- Maitip, J.; Mookhploy, W.; Khorndork, S.; Chantawannakul, P. Comparative Study of Antimicrobial Properties of Bee Venom Extracts and Melittins of Honey Bees. Antibiotics 2021, 10, 1503. [Google Scholar] [CrossRef]
- Haktanir, I.; Masoura, M.; Mantzouridou, F.T.; Gkatzionis, K. Mechanism of Antimicrobial Activity of Honeybee (Apis mellifera) Venom on Gram-Negative Bacteria: Escherichia coli and Pseudomonas spp. AMB Expr. 2021, 11, 54. [Google Scholar] [CrossRef]
- Bakhiet, E.K.; Hussien, H.A.M.; Elshehaby, M. Apis mellifera Venom Inhibits Bacterial and Fungal Pathogens In Vitro. Pak. J. Biol. Sci. 2022, 25, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Babaie, M.; Ghaem panah, A.; Mehrabi, Z.; Mollaei, A.; Sima Khalilifard, B. Partial Purification and Characterization of Antimicrobial Effects from Snake (Echis carinatus), Scorpion (Mesosobuthus epues) and Bee (Apis mellifera) Venoms. Iran. J. Med. Microbiol. 2020, 14, 460–477. [Google Scholar] [CrossRef]
- Pucca, M.B.; Cerni, F.A.; Oliveira, I.S.; Jenkins, T.P.; Argemí, L.; Sørensen, C.V.; Ahmadi, S.; Barbosa, J.E.; Laustsen, A.H. Bee Updated: Current Knowledge on Bee Venom and Bee Envenoming Therapy. Front. Immunol. 2019, 10, 2090. [Google Scholar] [CrossRef]
- van den Bogaart, G.; Guzmán, J.V.; Mika, J.T.; Poolman, B. On the Mechanism of Pore Formation by Melittin. J. Biol. Chem. 2008, 283, 33854–33857. [Google Scholar] [CrossRef]
- Zarrinnahad, H.; Mahmoodzadeh, A.; Hamidi, M.P.; Mahdavi, M.; Moradi, A.; Bagheri, K.P.; Shahbazzadeh, D. Apoptotic Effect of Melittin Purified from Iranian Honey Bee Venom on Human Cervical Cancer HeLa Cell Line. Int. J. Pept. Res. Ther. 2018, 24, 563–570. [Google Scholar] [CrossRef]
- de Roodt, A.R.; Lanari, L.C.; Lago, N.R.; Bustillo, S.; Litwin, S.; Morón-Goñi, F.; Gould, E.G.; van Grootheest, J.H.; Dokmetjian, J.C.; Dolab, J.A.; et al. Toxicological Study of Bee Venom (Apis mellifera Mellifera) from Different Regions of the Province of Buenos Aires, Argentina. Toxicon 2020, 188, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Viana, G.A.; Freitas, C.I.A.; Almeida, J.G.L.d.; Medeiros, G.V.D.d.; Teófilo, T.d.S.; Rodrigues, V.H.V.; Coelho, W.A.C.; Batista, J.S. Antioxidant, Genotoxic, Antigenotoxic, and Antineoplastic Activities of Apitoxin Produced by Apis mellifera in Northeast, Brazil. Cienc. Rural 2021, 51, e20200545. [Google Scholar] [CrossRef]
- Frangieh, J.; Salma, Y.; Haddad, K.; Mattei, C.; Legros, C.; Fajloun, Z.; El Obeid, D. First Characterization of the Venom from Apis mellifera Syriaca, a Honeybee from the Middle East Region. Toxins 2019, 11, 191. [Google Scholar] [CrossRef] [PubMed]
- Somwongin, S.; Chantawannakul, P.; Chaiyana, W. Antioxidant Activity and Irritation Property of Venoms from Apis Species. Toxicon 2018, 145, 32–39. [Google Scholar] [CrossRef]
- Tanuwidjaja, I.; Svečnjak, L.; Gugić, D.; Levanić, M.; Jurić, S.; Vinceković, M.; Mrkonjić Fuka, M. Chemical Profiling and Antimicrobial Properties of Honey Bee (Apis mellifera L.) Venom. Molecules 2021, 26, 3049. [Google Scholar] [CrossRef]
- Yaacoub, C.; Rifi, M.; El-Obeid, D.; Mawlawi, H.; Sabatier, J.-M.; Coutard, B.; Fajloun, Z. The Cytotoxic Effect of Apis mellifera Venom with a Synergistic Potential of Its Two Main Components—Melittin and PLA2—On Colon Cancer HCT116 Cell Lines. Molecules 2021, 26, 2264. [Google Scholar] [CrossRef]
- Kamel, A.; Suleiman, W.; Elfeky, A.; El-Sherbiny, G.; Elhaw, M. Characterization of Bee Venom and Its Synergistic Effect Combating Antibiotic Resistance of Pseudomonas aeruginosa. Egypt. J. Chem. 2021, 65, 1–2. [Google Scholar] [CrossRef]
- Nehme, H.; Ayde, H.; El Obeid, D.; Sabatier, J.M.; Fajloun, Z. Potential Inhibitory Effect of Apis mellifera’s Venom and of Its Two Main Components—Melittin and PLA2—On Escherichia coli F1F0-ATPase. Antibiotics 2020, 9, 824. [Google Scholar] [CrossRef]
- Sobral, F.; Sampaio, A.; Falcão, S.; Queiroz, M.J.R.P.; Calhelha, R.C.; Vilas-Boas, M.; Ferreira, I.C.F.R. Chemical Characterization, Antioxidant, Anti-Inflammatory and Cytotoxic Properties of Bee Venom Collected in Northeast Portugal. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef]
- Noble, J.E. Quantification of Protein Concentration Using UV Absorbance and Coomassie Dyes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 536, pp. 17–26. ISBN 978-0-12-420070-8. [Google Scholar]
- Pérez-Delgado, O.; Espinoza-Vergara, M.A.; Castro-Vega, N.A.; Reyes-Montaño, E.A. Evaluación Preliminar de Actividad Antibacteriana in Vitro Del Veneno de Escorpión Hadruroides Charcasus (Karsch, 1879) Contra Pseudomonas aeruginosa y Staphylococcus aureus. Rev. Cuerpo Med. HNAAA 2019, 12, 6–12. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Tricine-Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis for the Separation of Proteins in the Range from 1 to 100 KDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Brunelle, J.L.; Green, R. Coomassie Blue Staining. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 541, pp. 161–167. ISBN 978-0-12-420119-4. [Google Scholar]
- Approved Standard M07; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 9th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012; Volume 32, ISBN 1-56238-784-7.
- Oddo, A.; Hansen, P.R. Hemolytic Activity of Antimicrobial Peptides. In Antimicrobial Peptides; Hansen, P.R., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; Volume 1548, pp. 427–435. ISBN 978-1-4939-6735-3. [Google Scholar]
Figure 1.
SDS polyacrylamide gel electrophoresis with Tricine (SDS-PAGE-Tricine) of the purified fraction; M denoted the marker lane (molecular weight 2–40 kDa) (molecular weight marker), PF denotes the protein fractions, and TV denotes total venom.
Figure 1.
SDS polyacrylamide gel electrophoresis with Tricine (SDS-PAGE-Tricine) of the purified fraction; M denoted the marker lane (molecular weight 2–40 kDa) (molecular weight marker), PF denotes the protein fractions, and TV denotes total venom.

Figure 2.
Microbial growth of S. aureus, E. coli, and P. aeruginosa incubated with the purified fraction of Apis mellifera venom.
Figure 2.
Microbial growth of S. aureus, E. coli, and P. aeruginosa incubated with the purified fraction of Apis mellifera venom.

Figure 3.
Hemolytic activity of the purified fraction of Apis mellifera venom.

Figure 4.
(a) beeWhisper 6.0. collector box; (b) electrostimulation of the collector box; (c) recovery of A. mellifera dry venom; (d) storage of the venom.
Figure 4.
(a) beeWhisper 6.0. collector box; (b) electrostimulation of the collector box; (c) recovery of A. mellifera dry venom; (d) storage of the venom.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pérez-Delgado, O.; Espinoza-Culupú, A.O.; López-López, E. Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru. Antibiotics 2023, 12, 779. https://doi.org/10.3390/antibiotics12040779
AMA Style
Pérez-Delgado O, Espinoza-Culupú AO, López-López E. Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru. Antibiotics. 2023; 12(4):779. https://doi.org/10.3390/antibiotics12040779
Chicago/Turabian StylePérez-Delgado, Orlando, Abraham Omar Espinoza-Culupú, and Elmer López-López. 2023. "Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru" Antibiotics 12, no. 4: 779. https://doi.org/10.3390/antibiotics12040779
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.