
Co-Selection of Bacterial Metal and Antibiotic Resistance in Soil Laboratory Microcosms


 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Heavy Metal Content of Soil and Microcosm Characterisation
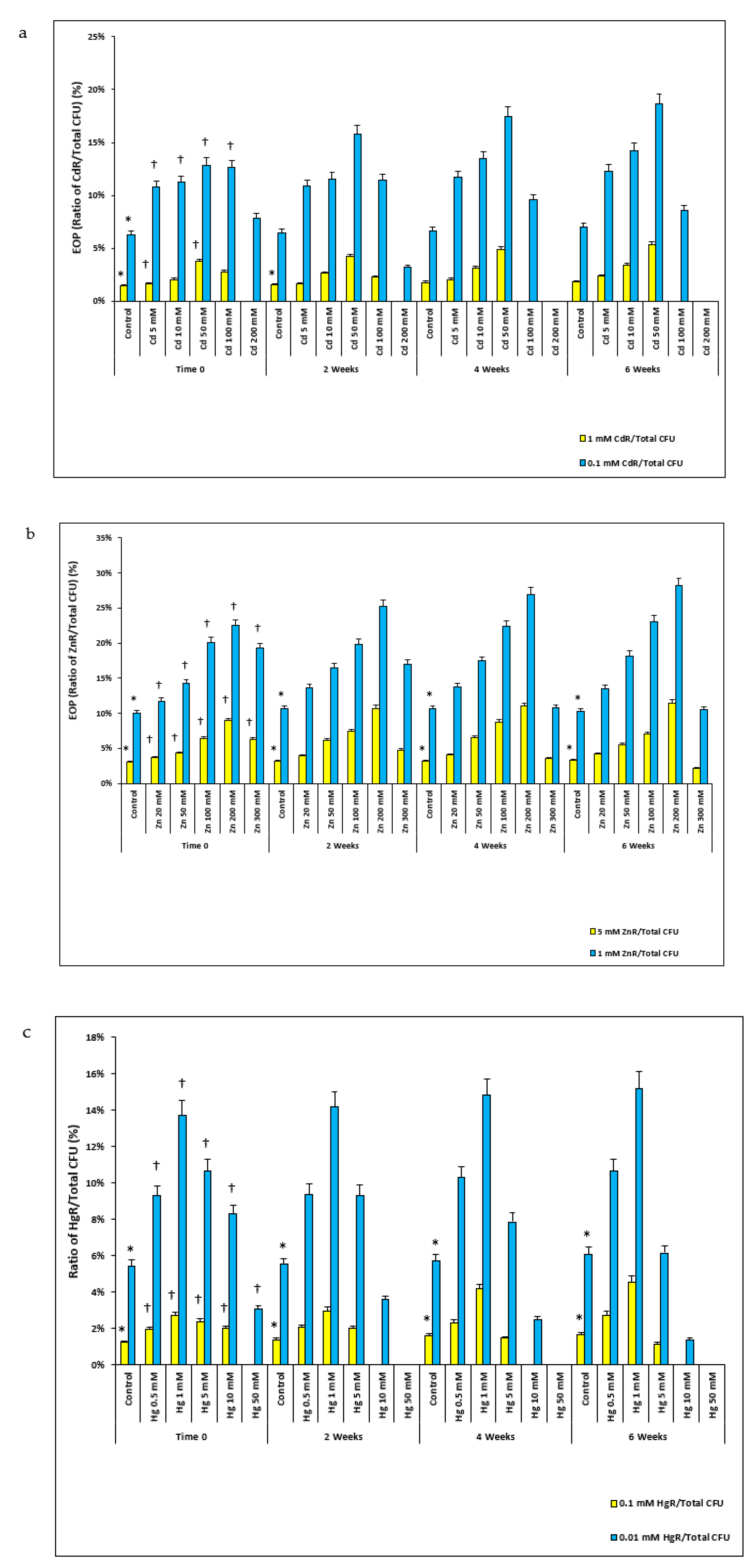
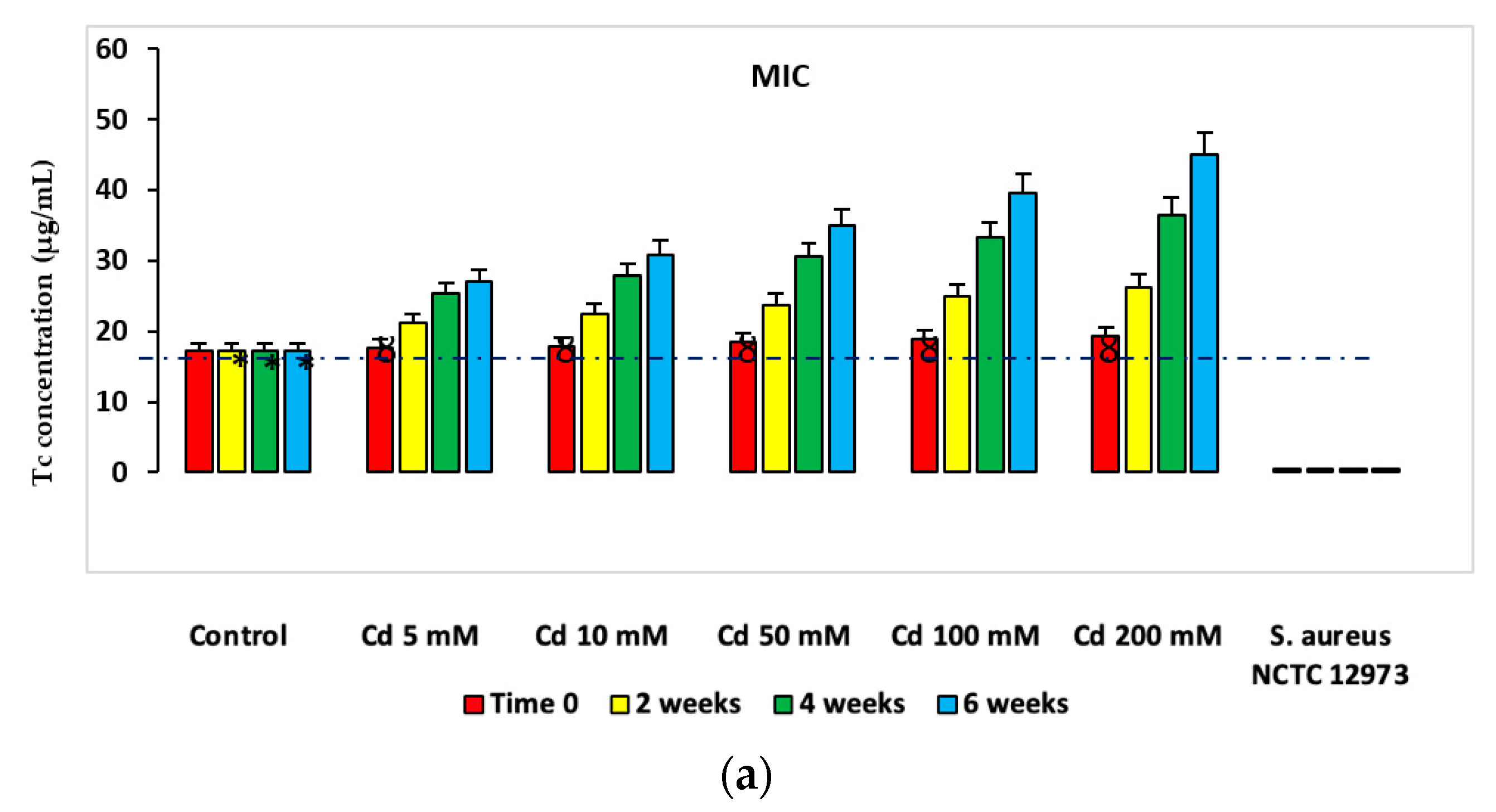
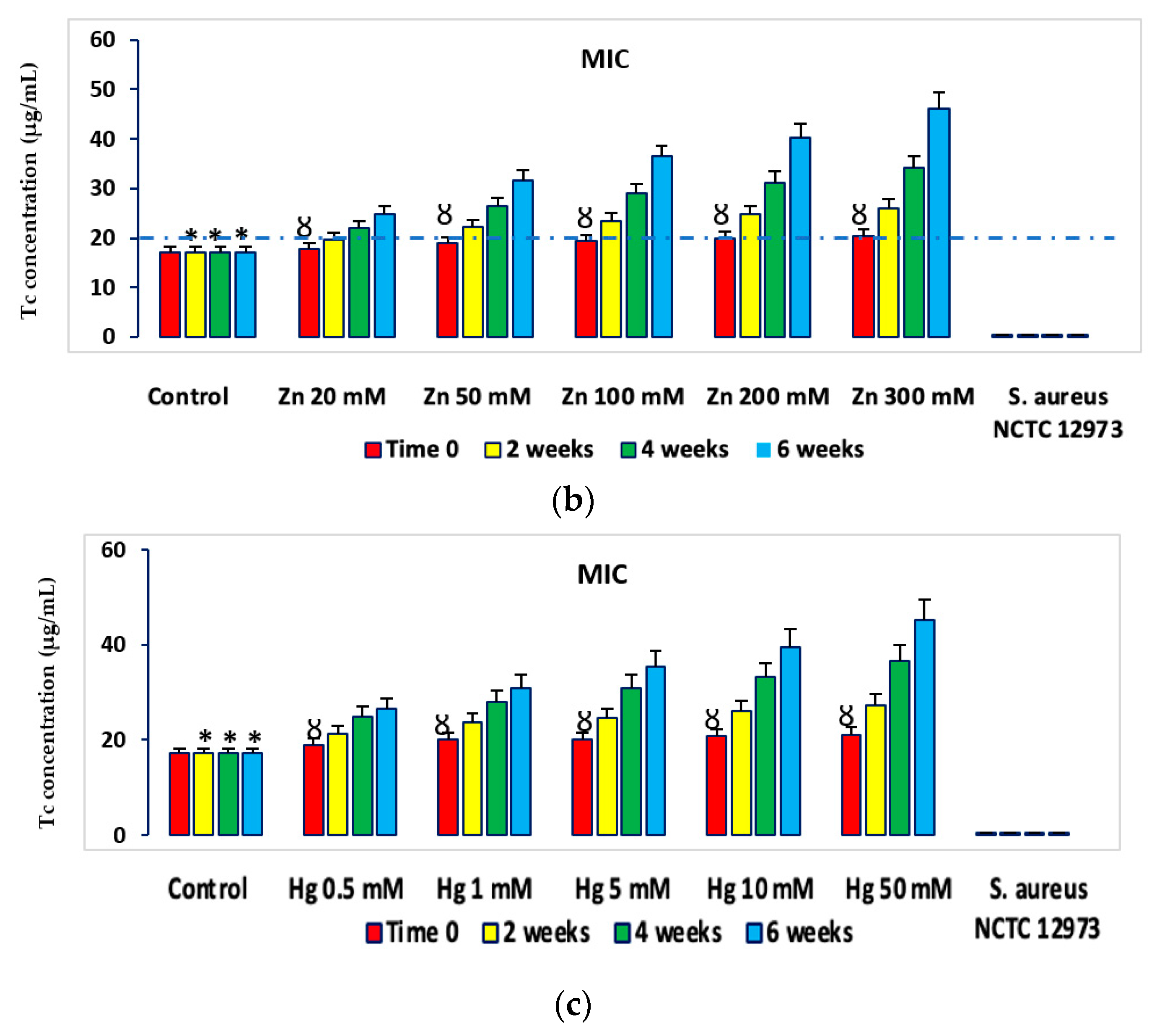
2.2. HM Resistance in Microcosms
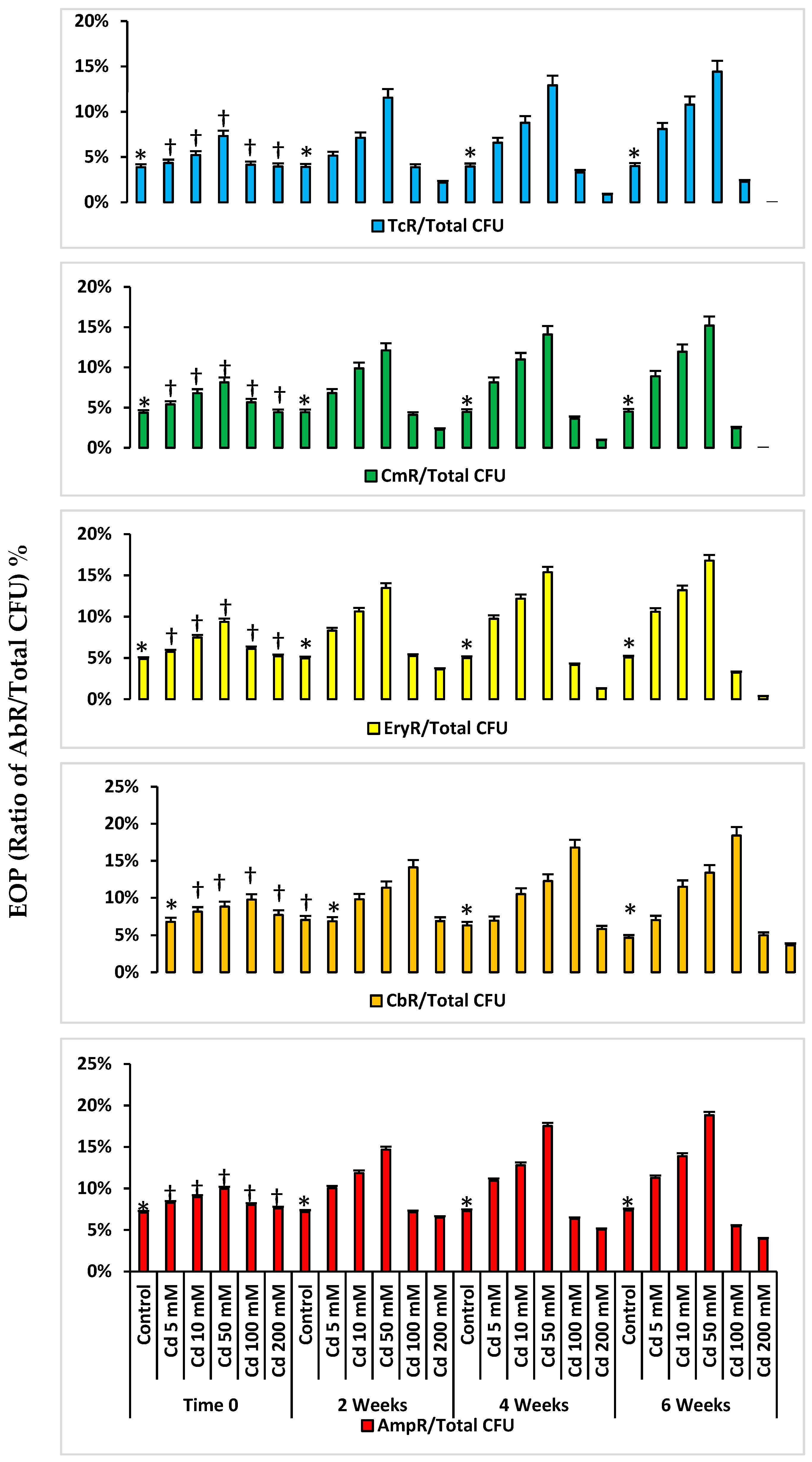
2.3. Antibiotic Resistance in Microcosms
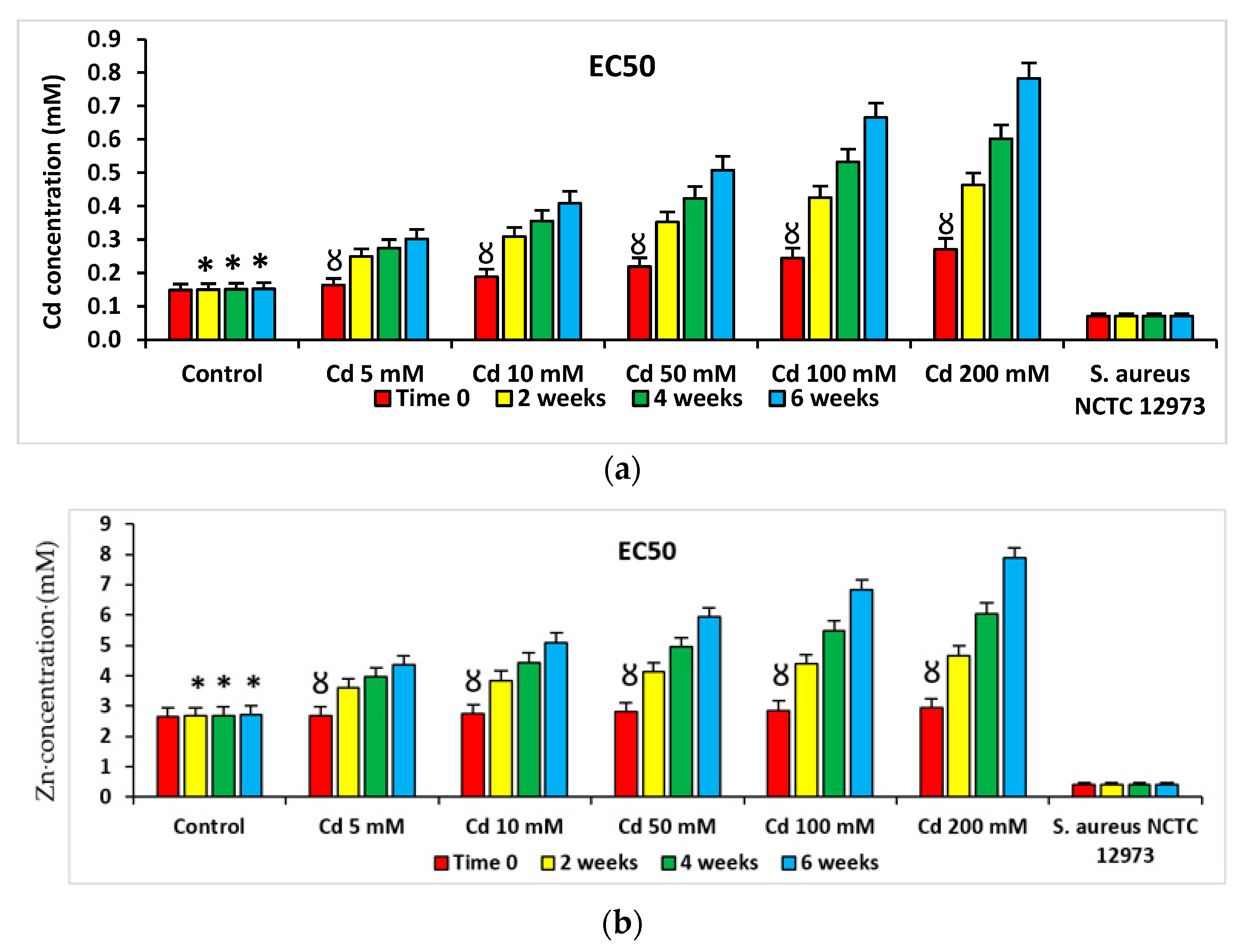
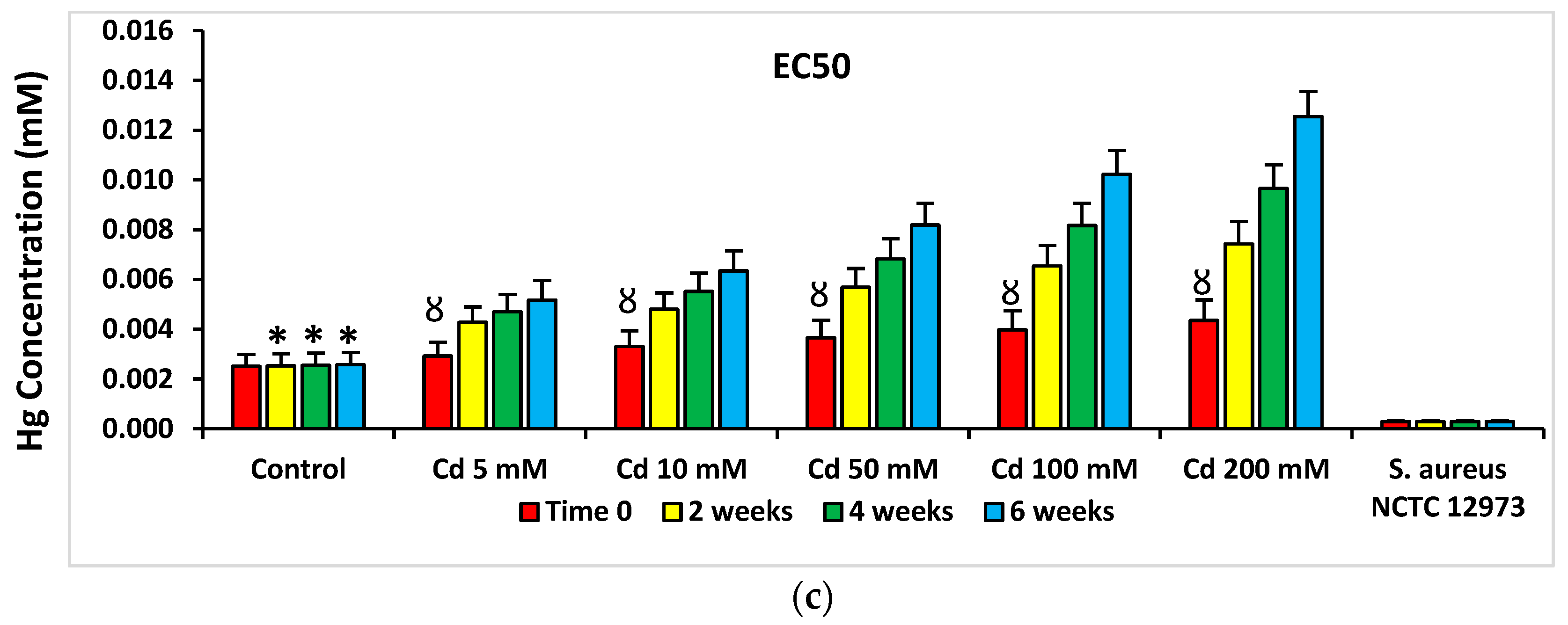
2.4. Pollution-Induced Community Tolerance (PICT) Analysis of Microcosms
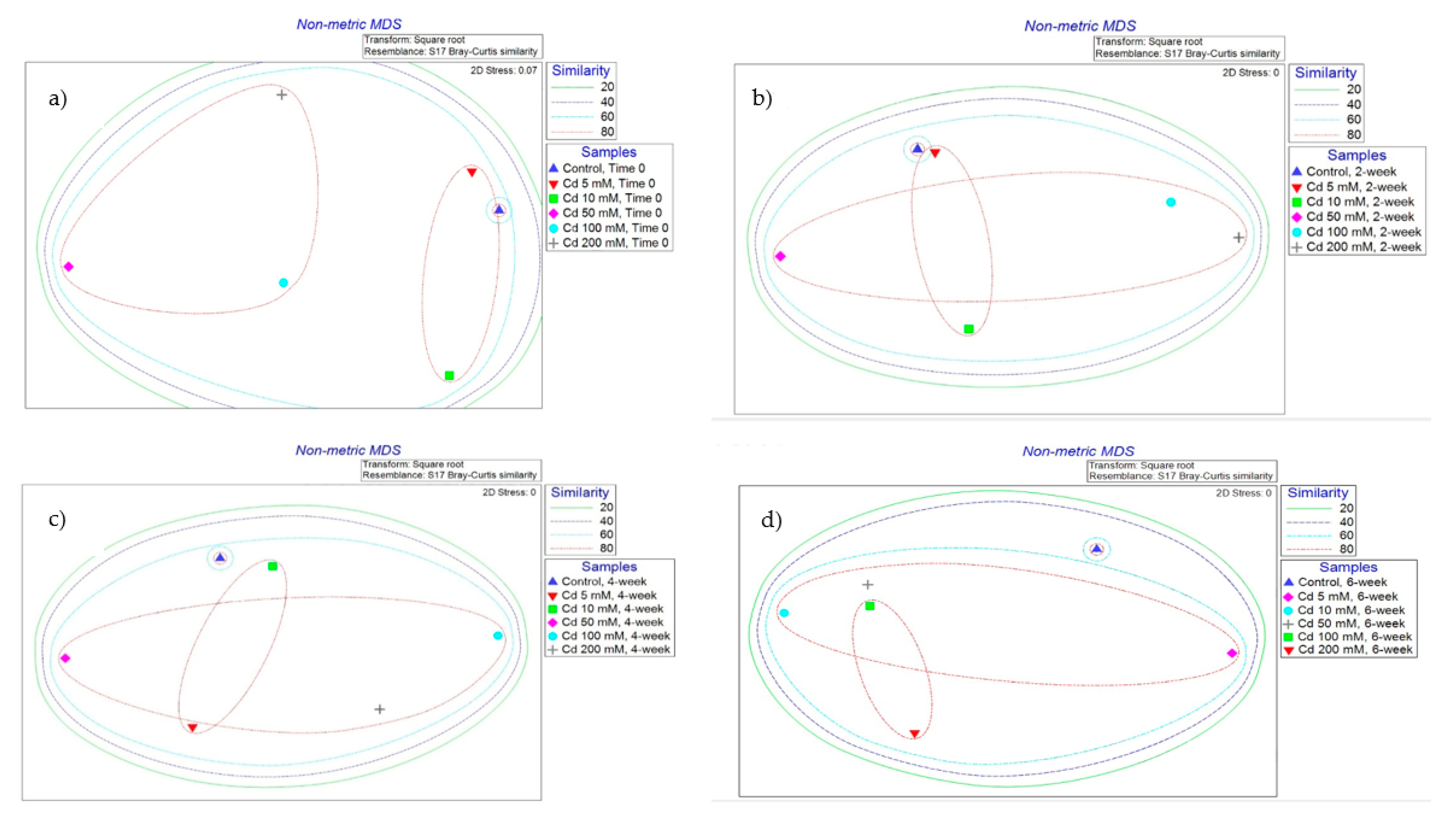
2.5. Terminal Restriction Fragment Length Polymorphism (TRFLP) Analysis of Soil DNA from Microcosms
2.5.1. Cd-Spiked Microcosms
2.5.2. Zn-Spiked Microcosms
2.5.3. Hg-Spiked Microcosms
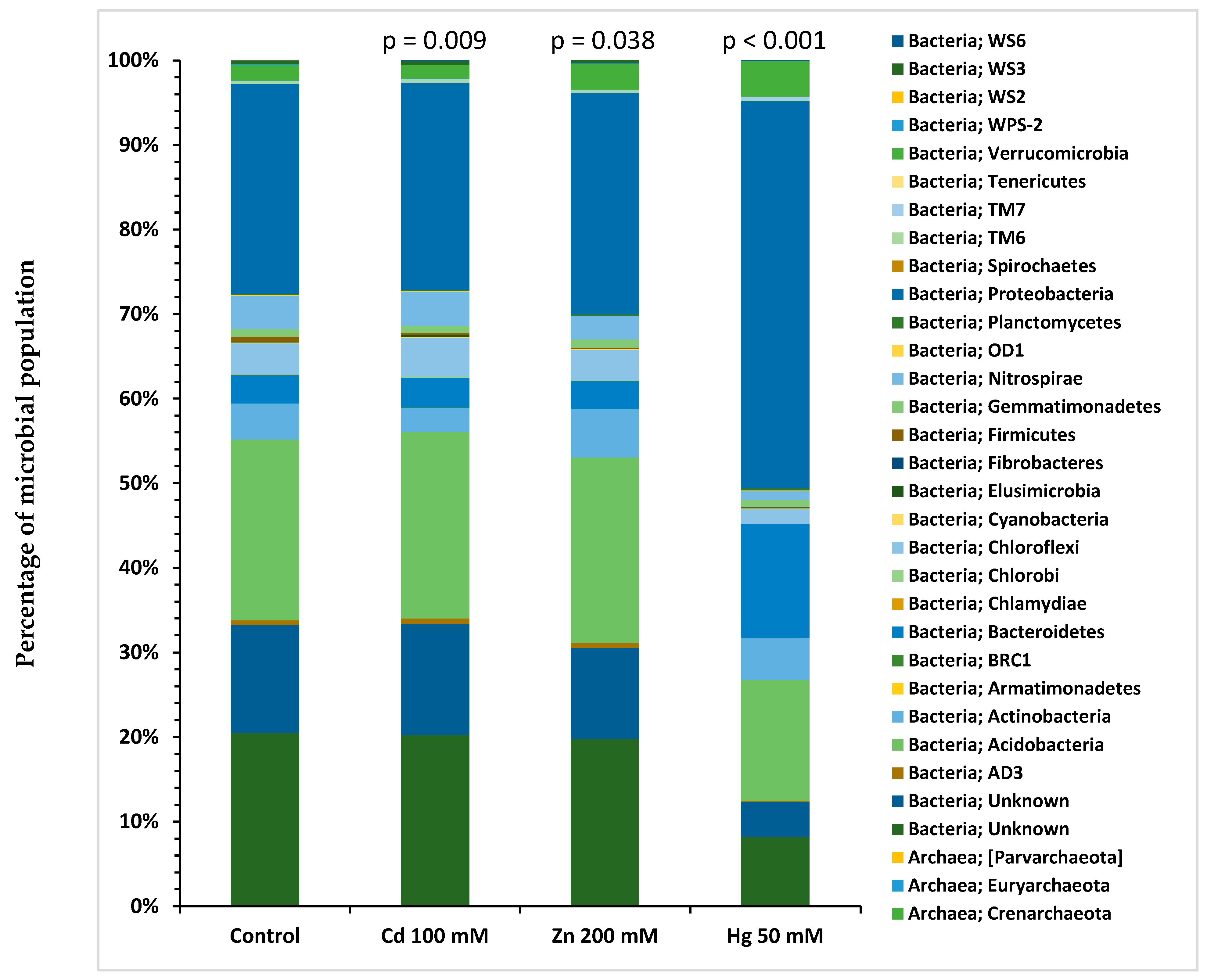
2.6. 16S rRNA Gene Analysis of Microbial Community Structure
3. Discussion
4. Materials and Methods
4.1. Physicochemical Properties of the Soil Sampling Sites
4.2. Microcosms’ Leachate HM Concentration Analysis
4.3. Microcosms
4.4. Plate Culture
4.5. Pollution-Induced Community Tolerance (PICT) Analysis
4.6. Soil DNA Extraction
4.7. Terminal Restriction Fragment Length Polymorphism (TRFLP)
4.8. 16S rDNA Sequencing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teitzel, G.M.; Parsek, M.R. Heavy metal resistance of biofilm and planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Bartl, B.; Hartl, W.; Horak, O. Long-term application of biowaste compost versus mineral fertilization: Effects on the nutrient and heavy metal contents of soil and plants. J. Plant Nutr. Soil Sci. 2002, 165, 161–165. [Google Scholar] [CrossRef]
- Heydari, A.; Kim, N.D.; Horswell, J.; Gielen, G.; Siggins, A.; Taylor, M.; Bromhead, C.; Palmer, B.R. Co-selection of Heavy Metal and Antibiotic Resistance in Soil Bacteria from Agricultural Soils in New Zealand. Sustainability 2022, 14, 1790. [Google Scholar] [CrossRef]
- Garau, G.; Castaldi, P.; Santona, L.; Deiana, P.; Melis, P. Influence of red mud, zeolite and lime on heavy metal immobilization, culturable heterotrophic microbial populations and enzyme activities in a contaminated soil. Geoderma 2007, 142, 47–57. [Google Scholar] [CrossRef]
- Tack, F.M.G. Trace Elements: General Soil Chemistry, Principles and Processes. In Trace Elements in Soils; Wiley: Hoboken, NJ, USA, 2010; pp. 9–37. [Google Scholar]
- Harrison, J.J.; Ceri, H.; Turner, R.J. Multimetal resistance and tolerance in microbial biofilms. Nat. Rev. Microbiol. 2007, 5, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Xie, X.M. Effect of heavy metals on substrate utilization pattern, biomass, and activity of microbial communities in a reclaimed mining wasteland of red soil area. Ecotoxicol. Environ. Saf. 2007, 66, 217–223. [Google Scholar] [CrossRef]
- Hay, S.I.; Rao, P.C.; Dolecek, C.; Day, N.P.J.; Stergachis, A.; Lopez, A.D.; Murray, C.J.L. Measuring and mapping the global burden of antimicrobial resistance. BMC Med. 2018, 16, 78. [Google Scholar] [CrossRef]
- Stanton, I.C.; Bethel, A.; Leonard, A.F.C.; Gaze, W.H.; Garside, R. What is the research evidence for antibiotic resistance exposure and transmission to humans from the environment? A systematic map protocol. Environ. Evid. 2020, 9, 12. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. MIC and Zone Diameter Distributions and ECOFFs. Available online: http://www.eucast.org/mic_distributions_and_ecoffs/ (accessed on 6 April 2023).
- D’Costa, V.M.; Griffiths, E.; Wright, G.D. Expanding the soil antibiotic resistome: Exploring environmental diversity. Curr. Opin. Microbiol. 2007, 10, 481–489. [Google Scholar] [CrossRef]
- Walsh, F. Investigating antibiotic resistance in non-clinical environments. Front. Microbiol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Chu, D. Effects of heavy metals on soil microbial community. IOP Conf. Ser. Earth Environ. Sci. 2018, 113, 012009. [Google Scholar] [CrossRef]
- Caliz, J.; Montserrat, G.; Martí, E.; Sierra, J.; Cruañas, R.; Garau, M.A.; Triadó-Margarit, X.; Vila, X. The exposition of a calcareous Mediterranean soil to toxic concentrations of Cr, Cd and Pb produces changes in the microbiota mainly related to differential metal bioavailability. Chemosphere 2012, 89, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Wang, C.; Zhao, Q.; Wang, Y.; Huo, M.; Wang, J.; Wang, S. Prevalence and dissemination of antibiotic resistance genes and coselection of heavy metals in Chinese dairy farms. J. Hazard. Mater. 2016, 320, 10–17. [Google Scholar] [CrossRef]
- Kim, N.D.; Taylor, M.D.; Drewry, J.J. Anthropogenic fluorine accumulation in the Waikato and Bay of Plenty regions of New Zealand: Comparison of field data with projections. Environ. Earth Sci. 2016, 75, 147. [Google Scholar] [CrossRef]
- Lighthart, B. Enrichment of cadmium-mediated antibiotic-resistant bacteria in a Douglas-fir (Pseudotsuga menziesii) litter microcosm. Appl. Environ. Microbiol. 1979, 37, 859–861. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef]
- Li, L.G.; Xia, Y.; Zhang, T. Co-occurrence of antibiotic and metal resistance genes revealed in complete genome collection. ISME J. 2017, 11, 651–662. [Google Scholar] [CrossRef]
- Zhao, Y.; Cocerva, T.; Cox, S.; Tardif, S.; Su, J.Q.; Zhu, Y.G.; Brandt, K.K. Evidence for co-selection of antibiotic resistance genes and mobile genetic elements in metal polluted urban soils. Sci. Total Environ. 2019, 656, 512–520. [Google Scholar] [CrossRef]
- Nakahara, H.; Ishikawa, T.; Sarai, Y.; Kondo, I.; Kozukue, H.; Silver, S. Linkage of mercury, cadmium, and arsenate and drug resistance in clinical isolates of Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1977, 33, 975–976. [Google Scholar] [CrossRef]
- Schipper, L.A.; Sparling, G.P.; Fisk, L.M.; Dodd, M.B.; Power, I.L.; Littler, R.A. Rates of accumulation of cadmium and uranium in a New Zealand hill farm soil as a result of long-term use of phosphate fertilizer. Agric. Ecosyst. Environ. 2011, 144, 95–101. [Google Scholar] [CrossRef]
- Hermans, S.M.; Taylor, M.; Grelet, G.; Curran-Cournane, F.; Buckley, H.L.; Handley, K.M.; Lear, G. From pine to pasture: Land use history has long-term impacts on soil bacterial community composition and functional potential. FEMS Microbiol. Ecol. 2020, 96, fiaa041. [Google Scholar] [CrossRef] [PubMed]
- Hermans, S.M.; Buckley, H.L.; Case, B.S.; Curran-Cournane, F.; Taylor, M.; Lear, G. Using soil bacterial communities to predict physico-chemical variables and soil quality. Microbiome 2020, 8, 79. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, Y.; Gao, S.; Chen, X. Copper exposure to soil under single and repeated application: Selection for the microbial community tolerance and effects on the dissipation of antibiotics. J. Hazard. Mater. 2017, 325, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhu, Y.; Dong, H.; Li, J.; Zhang, W.; Shao, Y.; Shao, Y. Effects of heavy metals and antibiotics on antibiotic resistance genes and microbial communities in soil. Process Saf. Environ. Prot. 2023, 169, 418–427. [Google Scholar] [CrossRef]
- Hewitt, A.E. New Zealand Soil Classification; Manaaki Whenua Press: Lincoln, New Zealand, 2010. [Google Scholar]
- Baillie, I.C. Soil Survey Staff 1999, Soil Taxonomy. Soil Use Manag. 2001, 17, 57–60. [Google Scholar] [CrossRef]
- Ong, G.H.; Wong, L.S.; Tan, A.L.; Yap, C.K. Effects of metal-contaminated soils on the accumulation of heavy metals in gotu kola (Centella asiatica) and the potential health risks: A study in Peninsular Malaysia. Environ. Monit. Assess. 2016, 188, 40. [Google Scholar] [CrossRef]
- Llobet, J.M.; Schuhmacher, M.; Domingo, J.L. Spatial distribution and temporal variation of metals in the vicinity of a municipal solid waste incinerator after a modernization of the flue gas cleaning systems of the facility. Sci. Total Environ. 2002, 284, 205–214. [Google Scholar] [CrossRef]
- Ozbek, N.; Akman, S. Molecule formation mechanisms of strontium mono fluoride in high-resolution continuum source electrothermal atomic absorption spectrometry. Anal. Sci. Int. J. Jpn. Soc. Anal. Chem. 2013, 29, 741–746. [Google Scholar] [CrossRef]
- MacLean, R.C. Adaptive radiation in microbial microcosms. J. Evol. Biol. 2005, 18, 1376–1386. [Google Scholar] [CrossRef]
- Yang, J.; Yang, F.; Yang, Y.; Xing, G.; Deng, C.; Shen, Y.; Luo, L.; Li, B.; Yuan, H. A proposal of “core enzyme” bioindicator in long-term Pb-Zn ore pollution areas based on topsoil property analysis. Environ. Pollut. 2016, 213, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Koeppel, A.F.; Wertheim, J.O.; Barone, L.; Gentile, N.; Krizanc, D.; Cohan, F.M. Speedy speciation in a bacterial microcosm: New species can arise as frequently as adaptations within a species. ISME J. 2013, 7, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Rozen, D.E.; Lenski, R.E. Long-Term Experimental Evolution in Escherichia coli. VIII. Dynamics of a Balanced Polymorphism. Am. Nat. 2000, 155, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Rousk, K. Responses of microbial tolerance to heavy metals along a century-old metal ore pollution gradient in a subarctic birch forest. Environ. Pollut. 2018, 240, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Wakelin, S.; Gerard, E.; Black, A.; Hamonts, K.; Condron, L.; Yuan, T.; van Nostrand, J.; Zhou, J.; O’Callaghan, M. Mechanisms of pollution induced community tolerance in a soil microbial community exposed to Cu. Environ. Pollut. 2014, 190, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Musken, M.; Di Fiore, S.; Romling, U.; Haussler, S. A 96-well-plate-based optical method for the quantitative and qualitative evaluation of Pseudomonas aeruginosa biofilm formation and its application to susceptibility testing. Nat. Protoc. 2010, 5, 1460–1469. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Papiernik, S.K.; Gan, J.; Yates, S.R.; Yang, C.H.; Crowley, D.E. Impact of fumigants on soil microbial communities. Appl. Environ. Microbiol. 2001, 67, 3245–3257. [Google Scholar] [CrossRef]
- van Elsas, J.D.; Mäntynen, V.; Wolters, A.C. Soil DNA extraction and assessment of the fate of Mycobacterium chlorophenolicum strain PCP-1 in different soils by 16S ribosomal RNA gene sequence based most-probable-number PCR and immunofluorescence. Biol. Fertil. Soils 1997, 24, 188–195. [Google Scholar] [CrossRef]
- Hauben, L.; Vauterin, L.; Swings, J.; Moore, E. Comparison of 16S ribosomal DNA sequences of all Xanthomonas species. Int. J. Syst. Evol. Microbiol. 1997, 47, 328–335. [Google Scholar] [CrossRef]
- Horswell, J.; Prosser, J.; Siggins, A.; Van Schaik, A.; Ying, L.; Ross, C.; McGill, A.; Northcott, G. Assessing the impacts of chemical cocktails on the soil ecosystem. Soil Biol. Biochem. 2014, 75, 64–72. [Google Scholar] [CrossRef]
- Gielen, G.J.; Clinton, P.W.; Van den Heuvel, M.R.; Kimberley, M.O.; Greenfield, L.G. Influence of sewage and pharmaceuticals on soil microbial function. Environ. Toxicol. Chem. 2011, 30, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | Measurement 1 |
|---|---|
| pH | 5.90 ± 0.065 |
| Total C (%) | 5.10 ± 0.35 |
| Total N (%) | 0.36 ± 0.022 |
| C:N | 14.2 ± 0.97 |
| Olsen P | 40.0 ± 4.72 2 |
| Cd | 0.13 ± 0.018 2 |
| Hg | 0.05 ± 0.009 2 |
| Zn | 19.4 ± 3.20 2 |
| Fe | 4500 ± 460 2 |
| P | 1090 ± 120 2 |
| Taxonomy Levels | Cd 100 mM Spiked Microcosms | Zn 200 mM Spiked Microcosms | Hg 50 mM Spiked Microcosms |
|---|---|---|---|
| Phylum | p = 0.009 | p = 0.038 | p < 0.001 |
| Class | p = 0.008 | p = 0.030 | p < 0.001 |
| Order | p = 0.007 | p = 0.020 | p < 0.001 |
| Family | p = 0.006 | p = 0.015 | p < 0.001 |
| Genus | p = 0.006 | p = 0.010 | p < 0.001 |
| Species | p = 0.006 | p = 0.010 | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heydari, A.; Kim, N.D.; Biggs, P.J.; Horswell, J.; Gielen, G.J.H.P.; Siggins, A.; Taylor, M.D.; Bromhead, C.; Palmer, B.R. Co-Selection of Bacterial Metal and Antibiotic Resistance in Soil Laboratory Microcosms. Antibiotics 2023, 12, 772. https://doi.org/10.3390/antibiotics12040772
Heydari A, Kim ND, Biggs PJ, Horswell J, Gielen GJHP, Siggins A, Taylor MD, Bromhead C, Palmer BR. Co-Selection of Bacterial Metal and Antibiotic Resistance in Soil Laboratory Microcosms. Antibiotics. 2023; 12(4):772. https://doi.org/10.3390/antibiotics12040772
Chicago/Turabian StyleHeydari, Ali, Nick D. Kim, Patrick J. Biggs, Jacqui Horswell, Gerty J. H. P. Gielen, Alma Siggins, Matthew D. Taylor, Collette Bromhead, and Barry R. Palmer. 2023. "Co-Selection of Bacterial Metal and Antibiotic Resistance in Soil Laboratory Microcosms" Antibiotics 12, no. 4: 772. https://doi.org/10.3390/antibiotics12040772