
Fomitopsis officinalis: Spatial (Pileus and Hymenophore) Metabolomic Variations Affect Functional Components and Biological Activities

 ,
,  ,
,  , , ,
, , ,  ,
,  , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Mushroom Identification
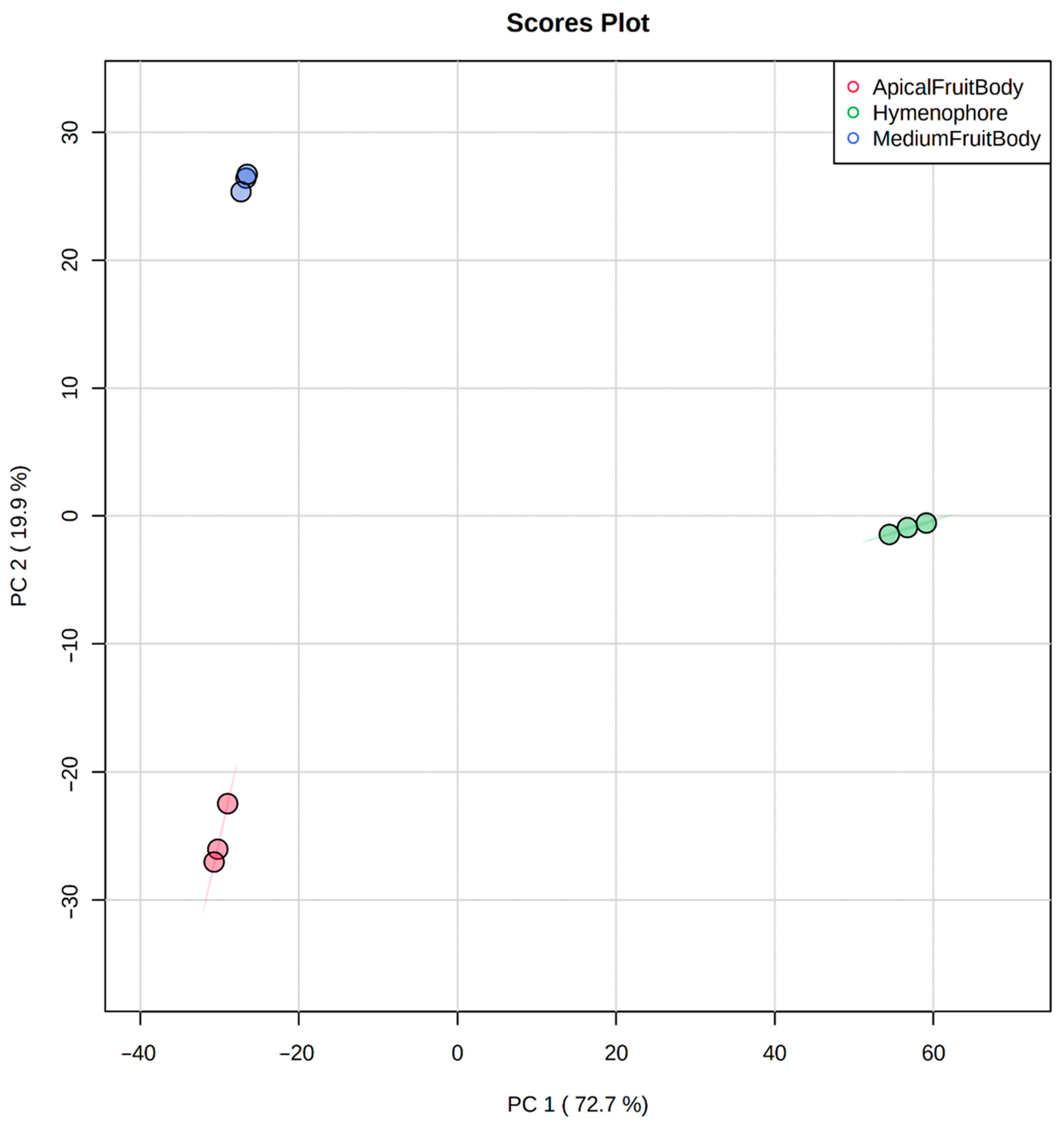
2.2. Untargeted LC–MS/MS-Based Metabolomics
2.3. Statistical Data Analysis
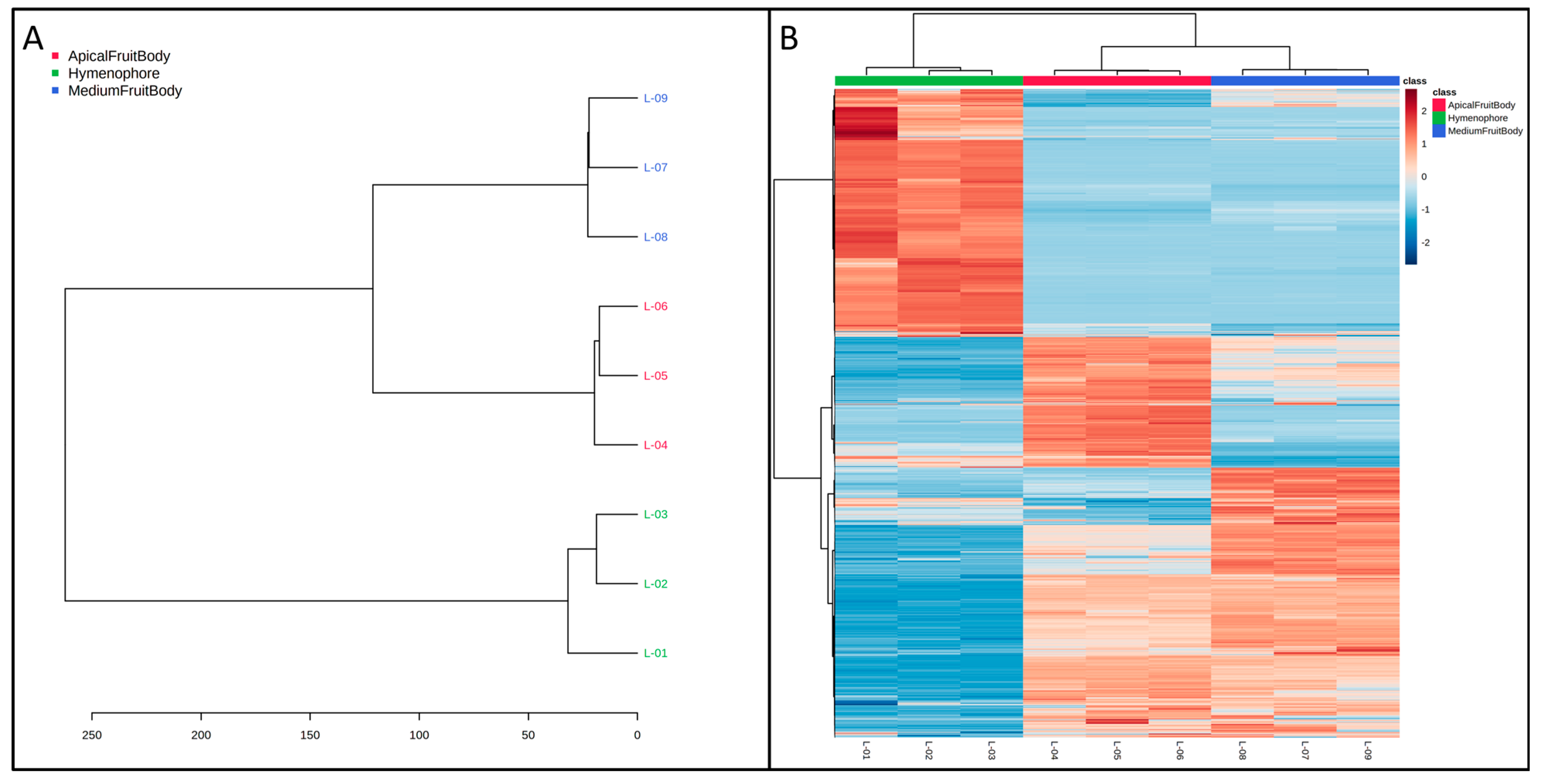
2.4. Cluster Analysis
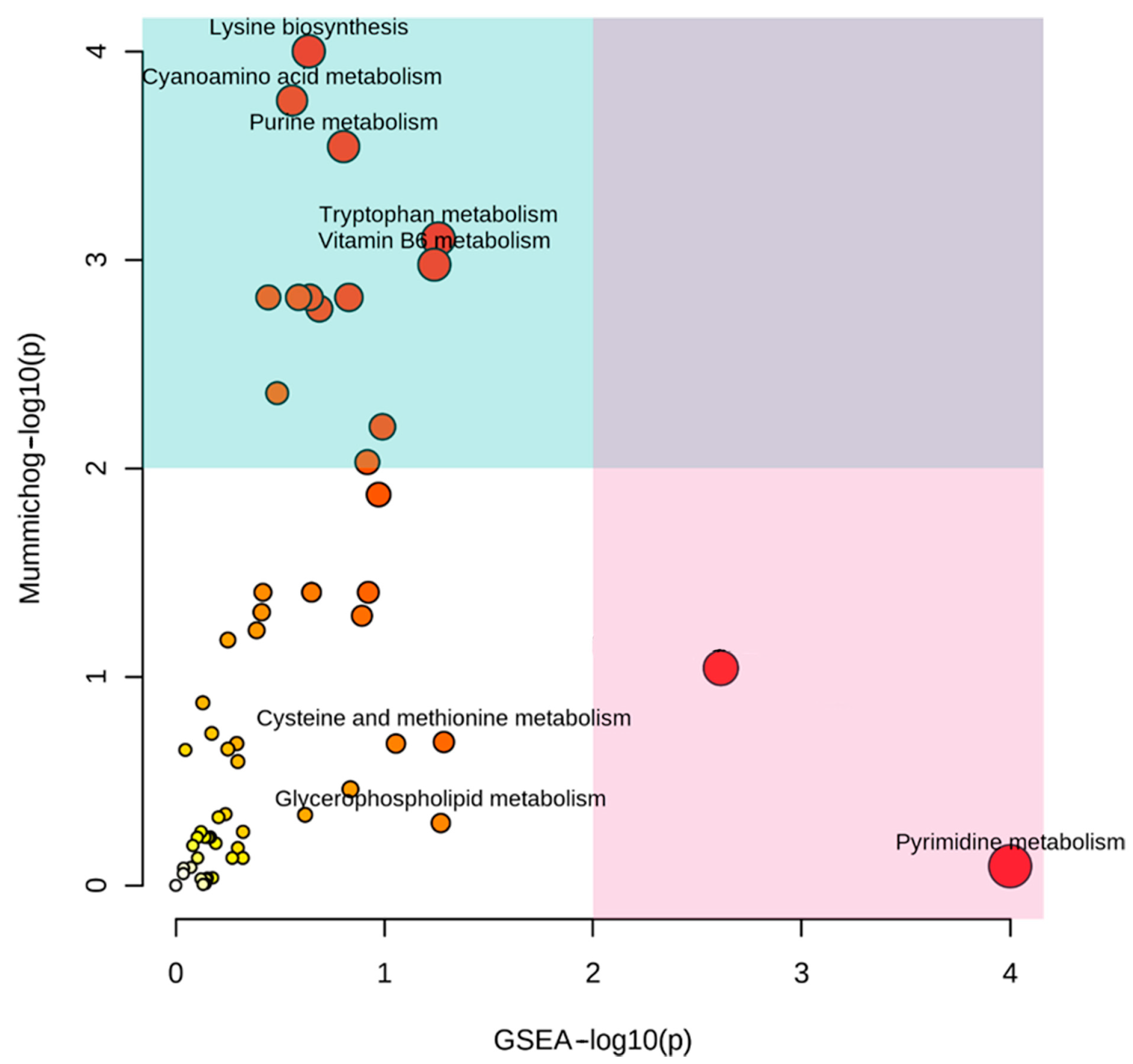
2.5. Functional Analysis
2.6. Pathway Analysis of the Apical Part with Respect to the Middle Part of the Fruiting Body
2.7. Extract Phenolic Composition and Antioxidant Activity
2.8. Antimicrobial Activity
3. Materials and Methods
3.1. Chemical and Reagents
3.2. Mushroom Strain
3.3. Molecular Identification
3.4. Mushroom Extract Preparation
3.5. Spectrophotometric Assays
3.5.1. Determination of Total Phenolic Content
3.5.2. Determination of Antioxidant Activity
3.6. Untargeted LC–MS/MS-Based Metabolomics and Statistical Analysis
3.7. HPLC Determination of Phenolic Compounds
3.8. Antimicrobial Test
Bacterial and Fungal Strains
3.9. Antibacterial Activity
3.10. Antifungal Activity
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernicchia, A. Polyporaceae sl.; Candusso: Alassio Savona, Italy, 2005; pp. 222–224. [Google Scholar]
- Ryvarden, L.; Melo, I. Poroid Fungi of Europe; Fungiflora: Oslo, Norway, 2014. [Google Scholar]
- Girometta, C. Antimicrobial properties of Fomitopsis officinalis in the light of its bioactive metabolites: A review. Mycology 2018, 10, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Hayova, V.P.; Heluta, V.P.; Shevchenko, M.V. Fomitopsis officinalis (Polyporales): Are there any records of the fungus known from Ukraine? Ukr. Bot. J. 2020, 77, 40–43. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Ryvarden, L. North American Polypores. Volume 1: Abortiporus—Lindtneria. 433 S., 209 Abb. Oslo 1986. Fungiflora A/S. J. Basic Microbiol. 1987, 27, 282. [Google Scholar] [CrossRef]
- Grienke, U.; Zöll, M.; Peintner, U.; Rollinger, J.M. European medicinal polypores—A modern view on traditional uses. J. Ethnopharmacol. 2014, 154, 564–583. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, J.-S.; Yan, M. Four New Triterpenes from Fungus of Fomes officinalis. Chem. Pharm. Bull. 2009, 57, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Li, L.; Zhong, J.; Tohtaton, Z.; Ren, Q.; Han, L.; Huang, X.; Yuan, T. Officimalonic acids A−H, lanostane triterpenes from the fruiting bodies of Fomes officinalis. Phytochemistry 2016, 130, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Simi, M.; Prisco, G. Patent Application Publication. U.S. Patent 2018/0250085A1, 2018. [Google Scholar]
- Angelini, P.; Pellegrino, R.M.; Tirillini, B.; Flores, G.A.; Alabed, H.B.; Ianni, F.; Blasi, F.; Cossignani, L.; Venanzoni, R.; Orlando, G. Metabolomic profiling and biological activities of Pleurotus columbinus Quél. cultivated on different agri-food byproducts. Antibiotics 2021, 10, 1245. [Google Scholar] [CrossRef]
- Flores, G.A.; Girometta, C.E.; Cusumano, G.; Angelini, P.; Tirillini, B.; Ianni, F.; Blasi, F.; Cossignani, L.; Pellegrino, R.M.; Emiliani, C. Untargeted Metabolomics Used to Describe the Chemical Composition, Antioxidant and Antimicrobial Effects of Extracts from Pleurotus spp. Mycelium Grown in Different Culture Media. Antibiotics 2022, 11, 1468. [Google Scholar] [CrossRef]
- Pellegrino, R.M.; Blasi, F.; Angelini, P.; Ianni, F.; Alabed, H.B.R.; Emiliani, C.; Venanzoni, R.; Cossignani, L. LC/MS Q-TOF Metabolomic Investigation of Amino Acids and Dipeptides in Pleurotus ostreatus Grown on Different Substrates. J. Agric. Food Chem. 2022, 70, 10371–10382. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Daba, G.M.; Elnahas, M.O.; Thomas, P.W. Fomitopsis officinalis mushroom: Ancient gold mine of functional components and biological activities for modern medicine. Egypt. Pharm. J. 2019, 18, 285. [Google Scholar] [CrossRef]
- Muszyńska, B.; Fijałkowska, A.; Sułkowska-Ziaja, K.; Włodarczyk, A.; Kaczmarczyk, P.; Nogaj, E.; Piętka, J. Fomitopsis officinalis: A Species of Arboreal Mushroom with Promising Biological and Medicinal Properties. Chem. Biodivers. 2020, 17, e2000213. [Google Scholar] [CrossRef]
- Hassan, K.; Kemkuignou, B.M.; Kirchenwitz, M.; Wittstein, K.; Rascher-Albaghdadi, M.; Chepkirui, C.; Matasyoh, J.C.; Decock, C.; Köster, R.W.; Stradal, T.E.B.; et al. Neurotrophic and Immunomodulatory Lanostane Triterpenoids from Wood-Inhabiting Basidiomycota. Int. J. Mol. Sci. 2022, 23, 13593. [Google Scholar] [CrossRef] [PubMed]
- Naranmandakh, S.; Murata, T.; Odonbayar, B.; Suganuma, K.; Batkhuu, J.; Sasaki, K. Lanostane triterpenoids from Fomitopsis officinalis and their trypanocidal activity. J. Nat. Med. 2018, 72, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, J.; Dong, Y. Chemical constituents of Fomes officinalis (I). Chin. Tradit. Herb. Drugs 2005, 36, 811–814. [Google Scholar]
- Wu, X.; Yang, J.; Zhou, L.; Dong, Y. New Lanostane-Type Triterpenes from Fomes officinalis. Chem. Pharm. Bull. 2004, 52, 1375–1377. [Google Scholar] [CrossRef] [PubMed]
- Angelini, P.; Girometta, C.; Tirillini, B.; Moretti, S.; Covino, S.; Cipriani, M.; D’Ellena, E.; Angeles, G.; Federici, E.; Savino, E.; et al. A comparative study of the antimicrobial and antioxidant activities of Inonotus hispidus fruit and their mycelia extracts. Int. J. Food Prop. 2019, 22, 768–783. [Google Scholar] [CrossRef]
- Ritota, M.; Manzi, P. Pleurotus spp. Cultivation on Different Agri-Food by-Products: Example of Biotechnological Application. Sustainability 2019, 11, 5049. [Google Scholar] [CrossRef]
- Bernicchia, A.; Gorjón, S.P.; Arras, L.; Facchini, M.; Porcu, G.; Trichies, G. Polypores of the Mediterranean Region; Romar SRL: Castellammare di Stabia, Italy, 2020. [Google Scholar]
- Ianni, F.; Blasi, F.; Angelini, P.; Simone, S.; Flores, G.A.; Cossignani, L.; Venanzoni, R. Extraction Optimization by Experimental Design of Bioactives from Pleurotus ostreatus and Evaluation of Antioxidant and Antimicrobial Activities. Processes 2021, 9, 743. [Google Scholar] [CrossRef]
- Adebayo, E.A.; Martínez-Carrera, D.; Morales, P.; Sobal, M.; Escudero, H.; Meneses, M.E.; Avila-Nava, A.; Castillo, I.; Bonilla, M. Comparative study of antioxidant and antibacterial properties of the edible mushrooms Pleurotus levis, P. ostreatus, P. pulmonarius and P. tuber-regium. Int. J. Food Sci. Technol. 2018, 53, 1316–1330. [Google Scholar] [CrossRef]
- Sánchez, C. Reactive oxygen species and antioxidant properties from mushrooms. Synth. Syst. Biotechnol. 2017, 2, 13–22. [Google Scholar] [CrossRef]
- López-Vázquez, E.; Prieto-García, F.; Gayosso-Canales, M.; Sánchez, E.O.; Ibarra, J.V. Phenolics acids, flavonoids, ascorbic acid, β-glucans and antioxidant activity in mexican wild edible mushrooms. Ital. J. Food Sci. 2017, 29, 766–774. [Google Scholar]
- Doğan, H.H.; Duman, R.; Özkalp, B.; Aydin, S. Antimicrobial activities of some mushrooms in Turkey. Pharm. Biol. 2013, 51, 707–711. [Google Scholar] [CrossRef]
- Alves, M.J.; Ferreira, I.C.F.R.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A Review on Antimicrobial Activity of Mushroom (Basidiomycetes) Extracts and Isolated Compounds. Planta Med. 2012, 78, 1707–1718. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B.; et al. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Politi, M.; Ferrante, C.; Menghini, L.; Angelini, P.; Flores, G.A.; Muscatello, B.; Braca, A.; De Leo, M. Hydrosols from Rosmarinus officinalis, Salvia officinalis, and Cupressus sempervirens: Phytochemical analysis and bioactivity evaluation. Plants 2022, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Nakabayashi, R.; Mori, T.; Yamada, Y.; Takahashi, M.; Rai, A.; Sugiyama, R.; Yamamoto, H.; Nakaya, T.; Yamazaki, M.; et al. A cheminformatics approach to characterize metabolomes in stable-isotope-labeled organisms. Nat. Methods 2019, 16, 295–298, Erratum in Nat. Methods 2019, 16, 446. [Google Scholar] [CrossRef]
- Li, S.; Park, Y.; Duraisingham, S.; Strobel, F.H.; Khan, N.; Soltow, Q.A.; Jones, D.P.; Pulendran, B. Predicting Network Activity from High Throughput Metabolomics. PLoS Comput. Biol. 2013, 9, e1003123. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Chiavaroli, A.; Libero, M.L.; Di Simone, S.C.; Acquaviva, A.; Nilofar; Recinella, L.; Leone, S.; Brunetti, L.; Cicia, D.; Izzo, A.A.; et al. Adding New Scientific Evidences on the Pharmaceutical Properties of Pelargonium quercetorum Agnew Extracts by Using In Vitro and In Silico Approaches. Plants 2023, 12, 1132. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Approved Standard, 4th ed.; CLSI Document M27-A4; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antifungal Susceptibility Testing of Yeasts; CLSI Supplement M60; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample ID | Country | Base Pair | Correspondence with Genbank Seq. | Identity Percentage | Accession No. |
|---|---|---|---|---|---|---|
| F. officinalis | MicUNIPV | Italy | 597 | F. officinalis | 100 | OL672134 |
| F. officinalis | JFo3619 | Poland | 599 | F. officinalis | 100 | MN534335 |
| F. officinalis | LE-BIN 3560 | Russia | 605 | F. officinalis | 99.81 | MG735354 |
| F. officinalis | 270279 | Russia | 553 | F. officinalis | 99.80 | MF952886 |
| F. officinalis | Stamets F04 | USA | 672 | F. officinalis | 99.62 | EU854437 |
| Pathway Name | Total_Size | Hits | Sig_Hits | Mummichog_Pvals | GSEA_Pvals | Combined_Pvals |
|---|---|---|---|---|---|---|
| Pyrimidine metabolism | 17 | 17 | 4 | 0.937 | 0.01 | 0.05313 |
| One carbon pool by folate | 4 | 4 | 2 | 0.4849 | 0.04938 | 0.1133 |
| Sphingolipid metabolism | 4 | 4 | 2 | 0.4849 | 0.04938 | 0.1133 |
| Tryptophan metabolism | 12 | 12 | 7 | 0.1163 | 0.2347 | 0.1256 |
| Lysine biosynthesis | 15 | 15 | 9 | 0.06231 | 0.48 | 0.1349 |
| Vitamin B6 metabolism | 10 | 10 | 6 | 0.1267 | 0.24 | 0.1366 |
| Purine metabolism | 27 | 27 | 14 | 0.08555 | 0.396 | 0.1485 |
| Cyano amino acid metabolism | 7 | 7 | 5 | 0.07341 | 0.5263 | 0.1643 |
| Pathway Name | Match Status | p Value | −LOG(p) | Holm p | FDR | Impact |
|---|---|---|---|---|---|---|
| Pyrimidine metabolism | 17/34 | 1.13 × 10−6 | 5.946 | 7.47 × 10−5 | 7.47 × 10−5 | 0.56111 |
| Tryptophan metabolism | 12/30 | 2.78 × 10−6 | 5.5561 | 1.81 × 10−4 | 9.13 × 10−5 | 0.42636 |
| Methane metabolism | 13/23 | 6.43 × 10−6 | 5.1918 | 4.12 × 10−4 | 9.13 × 10−5 | 0.42562 |
| Starch and sucrose metabolism | 2/15 | 7.90 × 10−6 | 5.1023 | 4.98 × 10−4 | 9.13 × 10−5 | 0.4374 |
| Folate biosynthesis | 12/23 | 8.51 × 10−6 | 5.0703 | 5.27 × 10−4 | 9.13 × 10−5 | 0.53932 |
| Galactose metabolism | 6/17 | 9.64 × 10−6 | 5.0159 | 5.88 × 10−4 | 9.13 × 10−5 | 0.20513 |
| Pantothenate and CoA biosynthesis | 11/20 | 9.69 × 10−6 | 5.0138 | 5.88 × 10−4 | 9.13 × 10−5 | 0.48294 |
| Phenylalanine, tyrosine, and tryptophan biosynthesis | 15/21 | 1.98 × 10−5 | 4.7036 | 0.001167 | 1.63 × 10−4 | 0.62752 |
| Valine, leucine, and isoleucine biosynthesis | 14/20 | 2.76 × 10−5 | 4.5585 | 0.001603 | 1.96 × 10−4 | 0.61768 |
| Glutathione metabolism | 11/26 | 2.97 × 10−5 | 4.5273 | 0.001693 | 1.96 × 10−4 | 0.28415 |
| Tyrosine metabolism | 10/15 | 3.39 × 10−5 | 4.4697 | 0.001899 | 2.03 × 10−4 | 0 |
| Glycerophospholipid metabolism | 4/32 | 5.24 × 10−5 | 4.2808 | 0.002881 | 2.88 × 10−4 | 0.05355 |
| Citrate (TCA) cycle | 10/20 | 6.61 × 05 | 4.1796 | 0.003571 | 3.32 × 10−4 | 0.46979 |
| Purine metabolism | 29/62 | 8.47 × 10−5 | 4.0719 | 0.004491 | 3.32 × 10−4 | 0.44416 |
| Pyruvate metabolism | 11/23 | 8.48 × 10−5 | 4.0714 | 0.004491 | 3.32 × 10−4 | 0.52672 |
| Cysteine and methionine metabolism | 18/41 | 8.56 × 10−5 | 4.0673 | 0.004491 | 3.32 × 10−4 | 0.60695 |
| Histidine metabolism | 8/18 | 8.64 × 10−5 | 4.0636 | 0.004491 | 3.32 × 10−4 | 0.46939 |
| beta-Alanine metabolism | 4/11 | 9.04 × 10−5 | 4.0436 | 0.004491 | 3.32 × 10−4 | 0.5 |
| Butanoate metabolism | 9/14 | 1.02 × 10−4 | 3.993 | 0.004878 | 3.53 × 10−4 | 0.6 |
| Sulfur metabolism | 7/13 | 1.16 × 10−4 | 3.9364 | 0.005441 | 3.71 × 10−4 | 0.25975 |
| Glycine, serine and threonine metabolism | 16/32 | 1.18 × 10−4 | 3.9278 | 0.005441 | 3.71 × 10−4 | 0.56013 |
| Biotin metabolism | 3/13 | 1.39 × 10−4 | 3.8571 | 0.006253 | 4.17 × 10−4 | 0.21277 |
| Lysine biosynthesis | 8/16 | 1.48 × 10−4 | 3.831 | 0.006493 | 4.23 × 10−4 | 0.52557 |
| Aminoacyl-tRNA biosynthesis | 16/46 | 1.55 × 10−4 | 3.8106 | 0.006651 | 4.25 × 10−4 | 0.16667 |
| Amino sugar and nucleotide sugar metabolism | 4/24 | 1.86 × 10−4 | 3.7315 | 0.007793 | 4.90 × 10−4 | 0.07092 |
| Glyoxylate and dicarboxylate metabolism | 14/26 | 2.03 × 10−4 | 3.6926 | 0.00832 | 5.15 × 10−4 | 0.76807 |
| Glycolysis/gluconeogenesis | 6/24 | 2.17 × 10−4 | 3.6645 | 0.008661 | 5.29 × 10−4 | 0.24445 |
| Pentose phosphate pathway | 7/18 | 2.67 × 10−4 | 3.5737 | 0.010409 | 6.29 × 10−4 | 0.18245 |
| Alanine, aspartate and glutamate metabolism | 14/22 | 3.15 × 10−4 | 3.5012 | 0.011984 | 7.18 × 10−4 | 0.91008 |
| Pentose and glucuronate interconversions | 4/12 | 4.20 × 10−4 | 3.3763 | 0.015555 | 9.25 × 10−4 | 0.27273 |
| Ubiquinone and other terpenoid-quinone biosynthesis | 2/2 | 4.78 × 10−4 | 3.3209 | 0.017195 | 0.001017 | 0 |
| Biosynthesis of unsaturated fatty acids | 11/23 | 5.03 × 10−4 | 3.2988 | 0.017588 | 0.001037 | 0 |
| Arginine biosynthesis | 13/18 | 5.53 × 10−4 | 3.2576 | 0.018788 | 0.001105 | 0.69546 |
| Sphingolipid metabolism | 6/13 | 6.11 × 10−4 | 3.2143 | 0.020149 | 0.001185 | 0.76666 |
| C5-Branched dibasic acid metabolism | 3/4 | 7.42 × 10−4 | 3.1293 | 0.023758 | 0.001397 | 0 |
| Arginine and proline metabolism | 13/25 | 7.62 × 10−4 | 3.1181 | 0.023758 | 0.001397 | 0.59627 |
| Vitamin B6 metabolism | 4/11 | 7.89 × 10−4 | 3.1032 | 0.023758 | 0.001407 | 0.30769 |
| Porphyrin and chlorophyll metabolism | 12/23 | 8.48 × 10−4 | 3.0717 | 0.024588 | 0.001446 | 0.67455 |
| Nicotinate and nicotinamide metabolism | 8/12 | 8.73 × 10−4 | 3.0591 | 0.024588 | 0.001446 | 0.68086 |
| Atrazine degradation | 2/4 | 8.76 × 10−4 | 3.0574 | 0.024588 | 0.001446 | 0.5 |
| One carbon pool by folate | 5/8 | 0.001039 | 2.9835 | 0.027005 | 0.001672 | 0.63939 |
| Fatty acid degradation | 5/30 | 0.001234 | 2.9085 | 0.030861 | 0.00194 | 0.14049 |
| Selenocompound metabolism | 1/12 | 0.001829 | 2.7378 | 0.043894 | 0.002807 | 0.14286 |
| Propanoate metabolism | 2/19 | 0.001898 | 2.7218 | 0.043894 | 0.002846 | 0 |
| Terpenoid backbone biosynthesis | 7/16 | 0.002058 | 2.6865 | 0.045283 | 0.003019 | 0.41189 |
| N-Glycan biosynthesis | 3/31 | 0.002423 | 2.6156 | 0.050884 | 0.003477 | 0.07877 |
| Steroid biosynthesis | 7/30 | 0.002504 | 2.6014 | 0.050884 | 0.003516 | 0.41379 |
| Valine, leucine and isoleucine degradation | 6/18 | 0.002744 | 2.5617 | 0.052128 | 0.003772 | 0 |
| Nitrogen metabolism | 3/5 | 0.002913 | 2.5356 | 0.052437 | 0.003924 | 0 |
| Fructose and mannose metabolism | 2/14 | 0.003126 | 2.505 | 0.053139 | 0.004126 | 0 |
| Lysine degradation | 4/15 | 0.003409 | 2.4674 | 0.054543 | 0.004412 | 0.2 |
| Fatty acid elongation | 3/22 | 0.003517 | 2.4539 | 0.054543 | 0.004464 | 0 |
| Thiamine metabolism | 6/18 | 0.004475 | 2.3492 | 0.06265 | 0.005573 | 0.38119 |
| Riboflavin metabolism | 6/11 | 0.0061 | 2.2147 | 0.079301 | 0.007456 | 0.84849 |
| Monobactam biosynthesis | 2/4 | 0.007354 | 2.1335 | 0.088243 | 0.008824 | 0 |
| Fatty acid biosynthesis | 1/43 | 0.012753 | 1.8944 | 0.14028 | 0.01503 | 0 |
| Phosphatidylinositol signaling system | 2/26 | 0.014143 | 1.8494 | 0.14143 | 0.016094 | 0.08621 |
| Inositol phosphate metabolism | 2/22 | 0.014143 | 1.8494 | 0.14143 | 0.016094 | 0 |
| Glycerolipid metabolism | 1/14 | 0.017911 | 1.7469 | 0.14329 | 0.019943 | 0.07059 |
| Taurine and hypotaurine metabolism | 2/7 | 0.01813 | 1.7416 | 0.14329 | 0.019943 | 0 |
| Sesquiterpenoid and triterpenoid biosynthesis | 1/4 | 0.018708 | 1.728 | 0.14329 | 0.020241 | 0 |
| Cyanoamino acid metabolism | 5/8 | 0.035402 | 1.451 | 0.17701 | 0.037686 | 0 |
| Carbapenem biosynthesis | 1/3 | 0.10926 | 0.96153 | 0.43705 | 0.11447 | 0 |
| Lipoic acid metabolism | 1/6 | 0.15843 | 0.80017 | 0.47528 | 0.16338 | 0 |
| Arachidonic acid metabolism | 2/8 | 0.40465 | 0.39292 | 0.8093 | 0.41088 | 0 |
| Synthesis and degradation of ketone bodies | 1/3 | 0.55021 | 0.25947 | 0.8093 | 0.55021 | 0 |
| TPC mg GAE/100 g | ABTS mg TE/100 g | DPPH mg TE/100 g | FRAP mg TE/100 g | |
|---|---|---|---|---|
| L1–L3 hymenium | 89.61 ± 8.70 | 144.39 ± 4.95 | 18.44 ± 0.28 | 119.97 ± 2.65 |
| L4–L6 apical part | 116.12 ± 3.45 | 170.00 ± 41.70 | 104.06 ± 5.44 | 198.00 ± 1.48 |
| L7–L8 median part | 92.02 ± 2.07 | 157.08 ± 12.60 | 72.60 ± 4.42 | 135.52 ± 1.79 |
| TPC | ABTS | DPPH | FRAP | |
|---|---|---|---|---|
| TPC | - | 0.8216 | 0.6874 | 0.9886 |
| ABTS | 0.8216 | - | 0.9755 | 0.8956 |
| DPPH | 0.6874 | 0.9755 | - | 0.7817 |
| FRAP | 0.9886 | 0.8956 | 0.7817 | - |
| MIC (µg/mL) * | ||||||||
|---|---|---|---|---|---|---|---|---|
| Escherichia coli | Escherichia coli | Escherichia coli | Bacillus cereus | Pseudomonas aeruginosa | Bacillus subtilis | Salmonella typhi | Staphylococcus aureus | |
| Bacteria | (ATCC 10536) | (PeruMycA 2) | (PeruMycA 3) | (ATCC 12826) | (ATCC 15442) | (PeruMycA 6) | (PeruMycA 7) | (ATCC 6538) |
| Samples | ||||||||
| L1–L3 | 79.37 (50–100) | 158.74 (100–200) | 125.99 (100–200) | 158.74 (100–200) | >200 | 1.53–<1.53 | >200 | >200 |
| L4–L6 | 3.86 (3.06–6.12) | 15.53 (12.25-25) | 79.37 (50–100) | 19.71 (12.25–25) | 7.71 (6.12–12.25) | 79.37 (50–100) | 158.74 (100–200) | 31.49 (25–50) |
| L7–L9 | 7.71 (6.12–12.25) | 62.99 (50–100) | 158.74 (100–200) | 19.71 (12.25-25) | 125.99 (100–200) | 2.42 (1.53–3.06) | >200 | 39.68 (25–50) |
| Ciprofloxacin (µg/mL) | 31.49 (25–50) | 9.92 (6.25–12.5) | 79.37 (50–100) | 125.99 (100–200) | 125.99 (100–200) | 125.99 (100–200) | 79.37 (50–100) | 200–>200 |
| MIC (µg/mL) * | ||||
|---|---|---|---|---|
| Candida tropicalis | Candida albicans | Candida parapsilosis | Candida albicans | |
| Yeast Strain | (YEPGA 6184) | (YEPGA 6379) | (YEPGA 6551) | (YEPGA 6183) |
| Samples | ||||
| L1–L3 | 200–>200 | 200–>200 | >200 | >200 |
| L4–L6 | >200 | >200 | 200–>200 | >200 |
| L7–L9 | >200 | >200 | >200 | >200 |
| Fluconazole (µg/mL) | 2 | 1 | 4 | 2 |
| MIC (µg/mL) * | ||||||||
|---|---|---|---|---|---|---|---|---|
| Trichophyton interdigitalis | Trichophyton tonsurans | Trichophyton rubrum | Arthroderma quadrifidum | Trichophyton erinacei | Arthroderma gypseum | Arthroderma currei | Arthroderma insingulare | |
| Dermatophyte | (CCF 4823) | (CCF 4834) | (CCF 4933) | (CCF 5792) | (CCF 5930) | (CCF 6261) | (CCF 5207) | (CCF 5417) |
| Samples | ||||||||
| L1–L3 | 125.99 (100–200) | 200–>200 | 158.74 (100–200) | 125.99 (100–200) | 200–>200 | 200–>200 | 200–>200 | 200–>200 |
| L4–L6 | 31.49 (25–50) | 19.57 (12.25–50) | 125.99 (100–200) | 79.37 (50–100) | 79.37 (50–100) | 31.49 (25–50) | 31.49 (25–50) | 62.99 (50–100) |
| L7–L9 | 31.49 (25–50) | 31.49 (25–50) | 158.74 (100–200) | 125.99 (100–200) | 158.74 (100–200) | 158.74 (100–200) | 39.68 (25–50) | 158.74 (100–200) |
| Griseofulvin (µg/mL) | 2.52 (2–4) | 0.198 (0.125–0.25) | 1.26 (1–2) | >8 | 3.174 (2–4) | 1.587 (1–2) | >8 | >8 |
| Sample ID | Mushroom Sample | Dried Mushroom Weight (mg) | Added EtOH: H2O (mL) | Final Concentration (mg/mL) |
|---|---|---|---|---|
| L1 | Hymenium | 4700 | 50 | 94 |
| L2 | Hymenium | 4700 | 50 | 94 |
| L3 | Hymenium | 4700 | 50 | 94 |
| L4 | Apical part | 4700 | 50 | 94 |
| L5 | Apical part | 4700 | 50 | 94 |
| L6 | Apical part | 4700 | 50 | 94 |
| L7 | Median part | 4700 | 50 | 94 |
| L8 | Median part | 4700 | 50 | 94 |
| L9 | Median part | 4700 | 50 | 94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores, G.A.; Cusumano, G.; Ianni, F.; Blasi, F.; Angelini, P.; Cossignani, L.; Pellegrino, R.M.; Emiliani, C.; Venanzoni, R.; Zengin, G.; et al. Fomitopsis officinalis: Spatial (Pileus and Hymenophore) Metabolomic Variations Affect Functional Components and Biological Activities. Antibiotics 2023, 12, 766. https://doi.org/10.3390/antibiotics12040766
Flores GA, Cusumano G, Ianni F, Blasi F, Angelini P, Cossignani L, Pellegrino RM, Emiliani C, Venanzoni R, Zengin G, et al. Fomitopsis officinalis: Spatial (Pileus and Hymenophore) Metabolomic Variations Affect Functional Components and Biological Activities. Antibiotics. 2023; 12(4):766. https://doi.org/10.3390/antibiotics12040766
Chicago/Turabian StyleFlores, Giancarlo Angeles, Gaia Cusumano, Federica Ianni, Francesca Blasi, Paola Angelini, Lina Cossignani, Roberto Maria Pellegrino, Carla Emiliani, Roberto Venanzoni, Gokhan Zengin, and et al. 2023. "Fomitopsis officinalis: Spatial (Pileus and Hymenophore) Metabolomic Variations Affect Functional Components and Biological Activities" Antibiotics 12, no. 4: 766. https://doi.org/10.3390/antibiotics12040766