
Staphylococcus spp. Causatives of Infections and Carrier of blaZ, femA, and mecA Genes Associated with Resistance

 , , and
, , and
Abstract
:1. Introduction
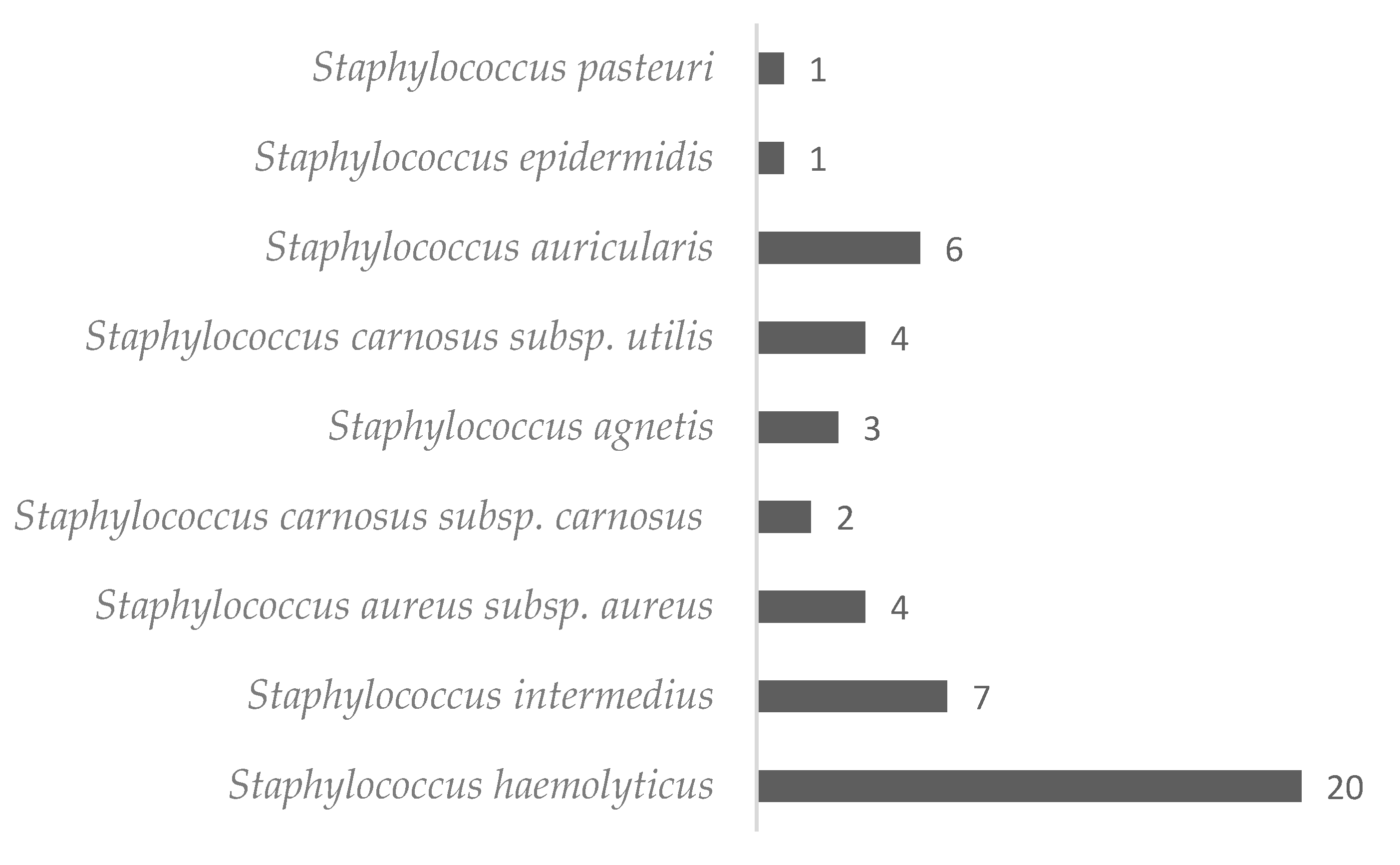
2. Results
3. Discussion
4. Materials and Methods
4.1. Data Collection
4.2. Bacterial Identification
4.3. DNA Extraction and Quantification
4.4. qPCR
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Braga, I.; Campos, P.; Gontijo-Filho, P.; Rjjohi, R. Multi-hospital point prevalence study of healthcare-associated infections in 28 adult intensive care units in Brazil. J. Hosp. Infect. 2018, 99, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, E.; Costa, T.C.D.; Silva, E.F.D. Prevenção e controle de infecção em unidade de terapia intensiva neonatal. Rev. Gaúcha Enferm. 2013, 34, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.C.; Paula, A.O.; Iquiapaza, R.A.; Lacerda, A.C.D.S. Infecções relacionadas à assistência em saúde e gravidade clínica em uma unidade de terapia intensiva. Rev. Gaúcha Enferm. 2012, 33, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, A.L.P.; Junior, A.C.S.S. Resistência bacteriana aos antibióticos e Saúde Pública: Uma breve revisão de literatura. Estação Científica 2017, 7, 45–57. [Google Scholar] [CrossRef]
- Amaral, L.d.S.; Godinho, S.M. Principais Fatores Causais de Infecções Relacionadas à Assistência à Saúde em Unidades de Terapia Intensiva: Uma Revisão Integrativa. 2020. Available online: https://dspace.uniceplac.edu.br/bitstream/123456789/289/1/Luisa%20Amaral_0003714_Silma%20Godinho_0003022.pdf (accessed on 19 February 2023).
- Andrew, E.F.; Friday, U.N.; Andrew, E.K.; Isaiah, L.N.; Silas, E.E.; Unah, U.V. Prevalence and antimicrobial susceptibility profile of bacterial isolates from infected caesarean sites in three federal capital territory hospitals, abuja nigeria. Am. J. Biomed. Life Sci. 2018, 6, 90–95. [Google Scholar] [CrossRef]
- Pittet, D.; Allegranzi, B.; Sax, H.; Dharan, S.; Pessoa-Silva, C.L.; Donaldson, L.; Boyce, J.M. Evidence-based model for hand transmission during patient care and the role of improved practices. Lancet Infect. Dis. 2006, 6, 641–652. [Google Scholar] [CrossRef]
- Santos, A.L.D.; Santos, D.O.; Freitas CCd Ferreira, B.L.A.; Afonso, I.F.; Rodrigues, C.R.; Castro, H.C. Staphylococcus aureus: Visiting a strain of clinical importance. J. Bras. Patol. Med. Lab. 2007, 43, 413–423. [Google Scholar] [CrossRef]
- Mandell, G.L. Catalase, superoxide dismutase, and virulence of Staphylococcus aureus. In vitro and in vivo studies with emphasis on staphylococcal--leukocyte interaction. J. Clin. Investig. 1975, 55, 561–566. [Google Scholar] [CrossRef]
- Bes, T.M.; Martins, R.R.; Perdigão, L.; Mongelos, D.; Moreno, L.; Moreno, A.; De Oliveira, G.S.; Costa, S.F.; Levin, A. Prevalence of methicillin-resistant Staphylococcus aureus colonization in individuals from the community in the city of Sao Paulo, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2018, 60, e58. [Google Scholar] [CrossRef]
- Appelbaum, P.C.; Bozdogan, B. Vancomycin resistance in Staphylococcus aureus. Clin. Lab. Med. 2004, 24, 381–402. [Google Scholar] [CrossRef]
- Foster, T. Staphylococcus. In Medical Microbiology, 4th ed.; Baron, S., Ed.; Chapter 12; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. Available online: https://pubmed.ncbi.nlm.nih.gov/21413338/ (accessed on 19 February 2023).
- Czekaj, T.; Ciszewski, M.; Szewczyk, E.M. Staphylococcus haemolyticus—An emerging threat in the twilight of the antibiotics age. Microbiology 2015, 161, 2061–2068. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C.; Ziebuhr, W.; Becker, K. Are coagulase-negative staphylococci virulent? Clin. Microbiol. Infect. 2019, 25, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Sizar, O.; Unakal, C.G. Gram Positive Bacteria; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Rossi, C.C.; Pereira, M.F.; Giambiagi-deMarval, M.J.G.; Biology, M. Underrated Staphylococcus species and their role in antimicrobial resistance spreading. Genet. Mol. Biol. 2020, 43, e20190065. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, E.C.; Marques, V.F.; Melo, D.A.; Alencar, T.A.; Coelho, I.d.S.; Coelho, S.M.; Souza, M. Caracterização fenogenotípica da resistência antimicrobiana em Staphylococcus spp. isolados de mastite bovina. Pesqui. Veterinária Bras. 2012, 32, 859–864. [Google Scholar] [CrossRef]
- El Feghaly, R.E.; Stamm, J.E.; Fritz, S.A.; Burnham, C.A.D. Presence of the blaZ beta-lactamase gene in isolates of Staphylococcus aureus that appear penicillin susceptible by conventional phenotypic methods. Diagn. Microbiol. Infect. Dis. 2012, 74, 388–393. [Google Scholar] [CrossRef]
- Gelatti, L.C.; Bonamigo, R.R.; Becker, A.P.; d’Azevedo, P.A. Staphylococcus aureus resistentes à meticilina: Disseminação emergente na comunidade. An. Bras. Dermatol. 2009, 84, 501–506. [Google Scholar] [CrossRef]
- Berger-Bächi, B.; Barberis-Maino, L.; Strässle, A.; Kayser, F.H. FemA, a host-mediated factor essential for methicillin resistance in Staphylococcus aureus: Molecular cloning and characterization. Mol. Gen. Genet. MGG 1989, 219, 263–269. [Google Scholar] [CrossRef]
- Andrade, C.R.; Filho, A.R.G.; Costa, A.C.M.; De Oliveira, T.A.; Carneiro, L.C.; Avelino, M.A.G. Identificação de Bactérias Causadoras de Infecção Hospitalar Utilizando Fenotipagem Clássica / Identification of Hospital Infectious Bacteria Using Classical Phenotyping. Braz. J. Dev. 2021, 7, 54446–54463. [Google Scholar] [CrossRef]
- Aguayo-Reyes, A.; Quezada-Aguiluz, M.; Mella, S.; Riedel, G.; Opazo-Capurro, A.; Bello-Toledo, H.; Domínguez, M.; González-Rocha, G. Bases moleculares de la resistencia a meticilina en Staphylococcus aureus. Rev. Chil. Infectol. 2018, 35, 7–14. [Google Scholar] [CrossRef]
- Deekshit, V.K.; Srikumar, S. ‘To be, or not to be’—The dilemma of ‘silent’ antimicrobial resistance genes in bacteria. J. Appl. Microbiol. 2022, 133, 2902–2914. [Google Scholar] [CrossRef] [PubMed]
- Laura, D.M.; Scott, N.L.; Vanner, E.A.; Miller, D.; Flynn, H.W., Jr. Genotypic and phenotypic antibiotic resistance in Staphylococcus epidermidis endophthalmitis. Ophthalmic Surg. Lasers Imaging Retin. 2020, 51, S13–S16. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed]
- d’Azevedo, P.A.; Secchi, C.; Antunes, A.L.; Sales, T.; Silva, F.M.; Tranchesi, R.; Pignatari, A.C.C. Oxacilin-resistant Coagulase-negative staphylococci (CoNS) bacteremia in a general hospital at São Paulo city, Brasil. Braz. J. Microbiol. 2008, 39, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Koksal, F.; Yasar, H.; Samasti, M. Antibiotic resistance patterns of coagulase-negative staphylococcus strains isolated from blood cultures of septicemic patients in Turkey. Microbiol. Res. 2009, 164, 404–410. [Google Scholar] [CrossRef]
- Medis, S.; Dissanayake, T.; Kottahachchi, J.; Namali, D.; Gunasekara, S.; Wijesinghe, G.; Dilrukshi, N.; Weerasekera, M. Biofilm formation and antibiotic resistance among Coagulase Negative Staphylococcus species isolated from central venous catheters of intensive care unit patients. Indian J. Med. Microbiol. 2023, 42, 71–76. [Google Scholar] [CrossRef]
- Shrestha, L.B.; Bhattarai, N.R.; Rai, K.; Khanal, B. Antibiotic resistance and mecA gene characterization of coagulase-negative staphylococci isolated from clinical samples in Nepal. Infect. Drug Resist. 2020, 13, 3163–3169. [Google Scholar] [CrossRef]
- Shah, P.; Dhungel, B.; Bastola, A.; Banjara, M.R.; Rijal, K.R.; Ghimire, P. Methicillin resistant Staphylococcus aureus in health care workers of a tertiary care infectious disease hospital in Nepal. Tribhuvan Univ. J. Microbiol. 2020, 7, 19–30. [Google Scholar] [CrossRef]
- Shwani, A.; Adkins, P.R.F.; Ekesi, N.S.; Alrubaye, A.; Calcutt, M.J.; Middleton, J.R.; Rhoads, D.D. Whole-Genome Comparisons of Staphylococcus agnetis Isolates from Cattle and Chickens. Appl. Environ. Microbiol. 2020, 86, e00484-20. [Google Scholar] [CrossRef]
- Szafraniec, G.; Szeleszczuk, P.; Dolka, B. A Review of Current Knowledge on Staphylococcus agnetis in Poultry. Animals 2020, 10, 1421. [Google Scholar] [CrossRef]
- Schleifer, K.H.; Fischer, U. Description of a New Species of the Genus Staphylococcus: Staphylococcus carnosus. Int. J. Syst. Evol. Microbiol. 1982, 32, 153–156. [Google Scholar] [CrossRef]
- Löfblom, J.; Rosenstein, R.; Nguyen, M.-T.; Ståhl, S.; Götz, F. Staphylococcus carnosus: From starter culture to protein engineering platform. Appl. Microbiol. Biotechnol. 2017, 101, 8293–8307. [Google Scholar] [CrossRef]
- Ramnarain, J.; Yoon, J.; Runnegar, N. Staphylococcus pasteuri infective endocarditis: A case report. IDCases 2019, 18, e00656. [Google Scholar] [CrossRef]
- Ha, E.T.; Heitner, J.F. Staphylococcus Auricularis Endocarditis: A Rare Cause of Subacute Prosthetic Valve Endocarditis with Severe Aortic Stenosis. Cureus 2021, 13, e2738. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.J.; Brown, G.D.; A Lombardo, F. Early-onset sepsis with Staphylococcus auricularis in an extremely low-birth weight infant–an uncommon pathogen. J. Perinatol. 2007, 27, 519–520. [Google Scholar] [CrossRef] [PubMed]
- Vuong, C.; Otto, M. Staphylococcus epidermidis infections. Microbes Infect. 2002, 4, 481–489. [Google Scholar] [CrossRef]
- Oliveira, W.; Silva, P.; Silva, R.; Silva, G.; Machado, G.; Coelho, L.; Correia, M. Staphylococcus aureus and Staphylococcus epidermidis infections on implants. J. Hosp. Infect. 2018, 98, 111–117. [Google Scholar] [CrossRef]
- Al-Amara, S.S. Comparison between phenotype and molecular resistance characteristic in Staphylococcus epidermidis isolates from wound infections in Al-Basrah province, Iraq. Period. Eng. Nat. Sci. (PEN) 2021, 9, 897–903. [Google Scholar] [CrossRef]
- Al-Essa, L.; Abunaja, M.H.; Hamadneh, L.; Abu-Sini, M. Mapping the intensive care unit environment and health care workers for methicillinresistant Staphylococcus aureus with mecA gene confirmation and antibacterial resistance pattern identification in a district hospital in Amman. Kuwait Med. J. 2021, 53, 265–270. [Google Scholar]
- Fernandes, L.F.; Souza, G.A.D.; De Almeida, A.C.; Cardoso, L.; Xavier, M.A.D.S.; Pinheiro, T.P.P.; Da Cruz, G.H.S.; Dourado, H.F.S.; Silva, W.S.; Xavier, A.R.E.D.O. Identification and characterization of methicillin-resistant Staphylococcus spp. isolated from surfaces near patients in an intensive care unit of a hospital in southeastern Brazil. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200244. [Google Scholar] [CrossRef]
- Al-Sultany, Z.K.; Al-Charrakh, A.H. Antibiotic resistance patterns of coagulase negative Staphylococcus (CoNS) strains isolated from blood stream infections in Babylon province, Iraq. Ann. Trop. Med. Public Health 2020, 23. [Google Scholar] [CrossRef]
- Asante, J.; Hetsa, B.A.; Amoako, D.G.; Abia, A.L.K.; Bester, L.A.; Essack, S.Y. Multidrug-resistant coagulase-negative staphylococci isolated from bloodstream in the uMgungundlovu District of KwaZulu-Natal Province in South Africa: Emerging pathogens. Antibiotics 2021, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; Torres, C. Actualización en la resistencia antibiótica en Gram positivos. Inf. Artículo 2017, 35, 2–8. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Monteiro, R.F.; dos Santos, V.R.R.; Ferreira, A.A.C.T.; de Abreu, J.R.G. O uso indiscriminado de antimicrobianos para o desenvolvimento de micro-organismos resistentes. Rev. Eletrônica Acervo Saúde 2020, 53, e3597. [Google Scholar] [CrossRef]
- Oliveira, M.; Pereira, K.D.S.P.S.; Zamberlam, C.R. Resistência Bacteriana Pelo Uso Indiscriminado De Antibióticos: Uma Questão De Saúde Pública. Rev. Ibero-Am. Humanid. Ciênc. Educ. 2020, 6, 183–201. [Google Scholar] [CrossRef]
- Bannoehr, J.; Ben Zakour, N.L.; Waller, A.S.; Guardabassi, L.; Thoday, K.L.; van den Broek, A.H.; Fitzgerald, J.R. Population genetic structure of the Staphylococcus intermedius group: Insights into agr diversification and the emergence of methicillin-resistant strains. J. Bacteriol. 2007, 189, 8685–8692. [Google Scholar] [CrossRef]
- Teixeira, J.P.; Silva, N.; da Fonseca, L.M.; da Costa, G.M. Uso de PCR Duplex para detecção dos genes femA e mecA e determinação da concentração inibitória mínima (CIM) em Staphylococcus aureus isolados de leite cru. Rev. Inst. Adolfo Lutz 2014, 73, 272–279. [Google Scholar] [CrossRef]
- Pearson, K. X. On the criterion that a given system of deviations from the probable in the case of a correlated system of variables is such that it can be reasonably supposed to have arisen from random sampling. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1900, 50, 157–175. [Google Scholar] [CrossRef]


{kind=link}
| S. haemolyticus | S. intermedius | S. aureus subsp. aureus | S. carnosus subsp. carnosus | S. agnetis | S. auricularis | S. epidermidis | S. carnosus subsp. utilis | S. pasteuri | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chromosomal DNA | blaZ | Total | 12 | 7 | 3 | 2 | 2 | 2 | 0 | 2 | 1 |
| Positive | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| % | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| femA | Total | 10 | 6 | 3 | 2 | 1 | 2 | 1 | 1 | 1 | |
| Positive | 3 | 6 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | ||
| % | 30 | 100 | 0 | 50 | 0 | 50 | 100 | 100 | 100 | ||
| mecA | Total | 13 | 5 | 3 | 1 | 2 | 4 | 0 | 3 | 1 | |
| Positive | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| % | 7.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Plasmid DNA | blaZ | Total | 10 | 7 | 2 | 2 | 1 | 2 | 1 | 3 | 1 |
| Positive | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | ||
| % | 10 | 14.3 | 0 | 50 | 100 | 50 | 0 | 33.3 | 0 | ||
| femA | Total | 12 | 7 | 3 | 2 | 0 | 1 | 0 | 3 | 1 | |
| Positive | 5 | 3 | 1 | 1 | 0 | 1 | 0 | 2 | 0 | ||
| % | 41.6 | 42.5 | 33.3 | 50 | 0 | 100 | 0 | 66.6 | 0 | ||
| mecA | Total | 12 | 7 | 3 | 2 | 1 | 2 | 0 | 3 | 1 | |
| Positive | 0 | 1 | 2 | 1 | 1 | 0 | 0 | 1 | 0 | ||
| % | 0 | 14.3 | 66.6 | 50 | 100 | 0 | 0 | 33.3 | 0 |
| Chromosomal | Plasmid | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | blaZ | % | femA | % | mecA | % | blaZ | % | femA | % | mecA | % |
| S. haemolyticus | 0 | 0 | 2 | 66.6 | 1 | 100 | 0 | 0 | 2 | 40 | 0 | 0 |
| S. intermedius | 0 | 0 | 4 | 66.6 | 0 | 0 | 1 | 100 | 2 | 66.6 | 1 | 100 |
| S. aureus subsp. aureus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. carnosus subsp. carnosus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. agnetis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 100 | 0 | 0 | 1 | 100 |
| S. auricularis | 0 | 0 | 1 | 100 | 0 | 0 | 1 | 100 | 1 | 100 | 0 | 0 |
| S. epidermidis | 0 | 0 | 1 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. carnosus subsp. utilis | 0 | 0 | 1 | 100 | 0 | 0 | 1 | 100 | 2 | 100 | 1 | 100 |
| S. pasteuri | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chromosomal | Plasmid | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | blaZ | % | femA | % | mecA | % | blaZ | % | femA | % | mecA | % |
| S. haemolyticus | 0 | 0 | 2 | 66.6 | 0 | 0 | 0 | 0 | 2 | 40 | 0 | 0 |
| S. intermedius | 0 | 0 | 3 | 50 | 0 | 0 | 1 | 100 | 2 | 66.6 | 0 | 0 |
| S. aureus subsp. aureus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 100 | 2 | 100 |
| S. carnosus subsp. carnosus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. agnetis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 100 | 0 | 0 | 1 | 100 |
| S. auricularis | 0 | 0 | 1 | 100 | 0 | 0 | 1 | 100 | 1 | 100 | 0 | 0 |
| S. epidermidis | 0 | 0 | 1 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| S. carnosus subsp. utilis | 0 | 0 | 1 | 100 | 0 | 0 | 1 | 100 | 2 | 100 | 1 | 100 |
| S. pasteuri | 0 | 0 | 1 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimenta, L.K.L.; Rodrigues, C.A.; Filho, A.R.G.; Coelho, C.J.; Goes, V.; Estrela, M.; de Souza, P.; Avelino, M.A.G.; Vieira, J.D.G.; Carneiro, L. Staphylococcus spp. Causatives of Infections and Carrier of blaZ, femA, and mecA Genes Associated with Resistance. Antibiotics 2023, 12, 671. https://doi.org/10.3390/antibiotics12040671
Pimenta LKL, Rodrigues CA, Filho ARG, Coelho CJ, Goes V, Estrela M, de Souza P, Avelino MAG, Vieira JDG, Carneiro L. Staphylococcus spp. Causatives of Infections and Carrier of blaZ, femA, and mecA Genes Associated with Resistance. Antibiotics. 2023; 12(4):671. https://doi.org/10.3390/antibiotics12040671
Chicago/Turabian StylePimenta, Laryssa Ketelyn Lima, Carolina Andrade Rodrigues, Arlindo Rodrigues Galvão Filho, Clarimar José Coelho, Viviane Goes, Mariely Estrela, Priscila de Souza, Melissa Ameloti Gomes Avelino, José Daniel Gonçalves Vieira, and Lilian Carneiro. 2023. "Staphylococcus spp. Causatives of Infections and Carrier of blaZ, femA, and mecA Genes Associated with Resistance" Antibiotics 12, no. 4: 671. https://doi.org/10.3390/antibiotics12040671