

Polyphosphate Ester-Type Transporters Improve Antimicrobial Properties of Oxytetracycline
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
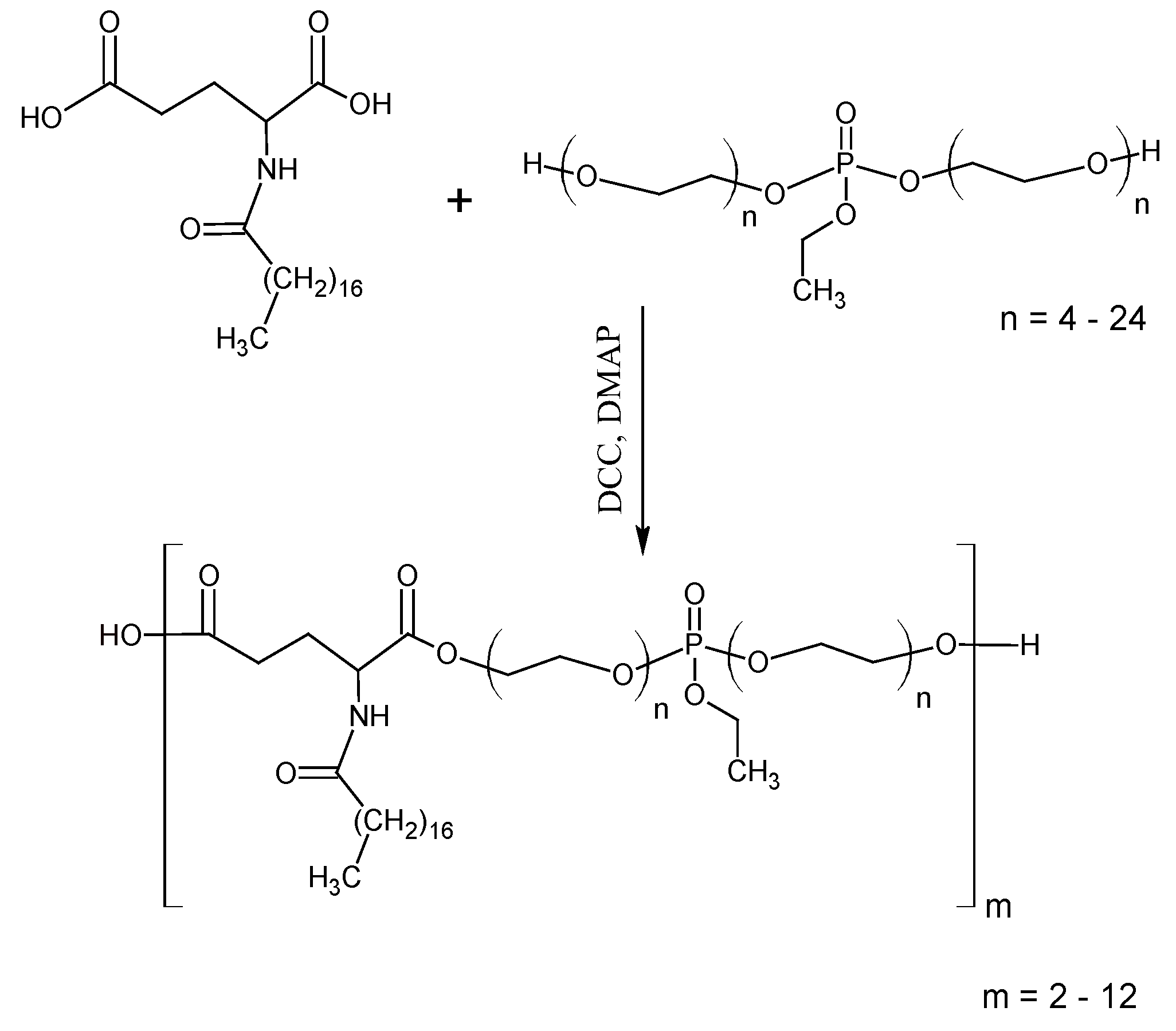
2.1. Synthesis of the Oxytetracycline Complex with a Polyphosphate Ester Type Transporter
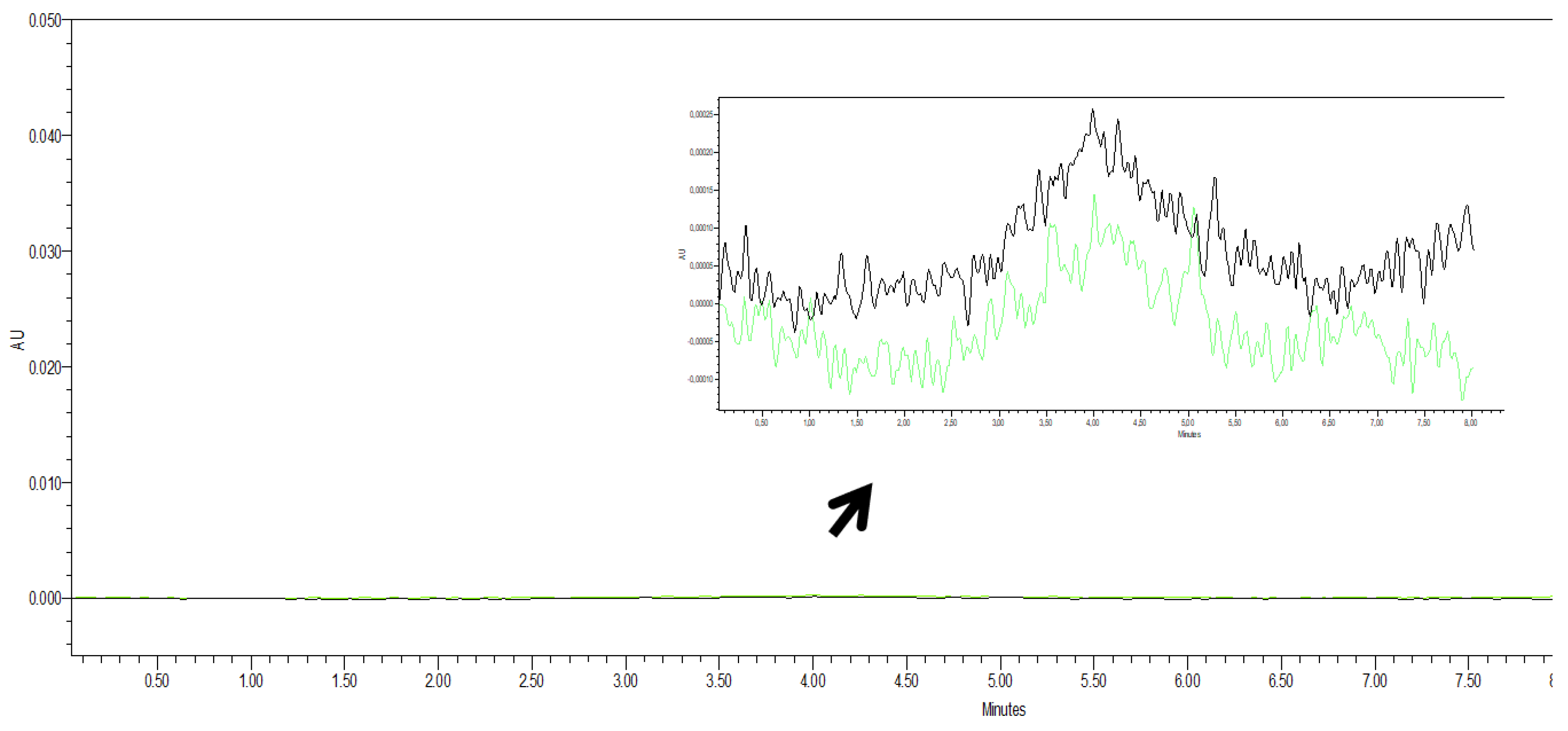
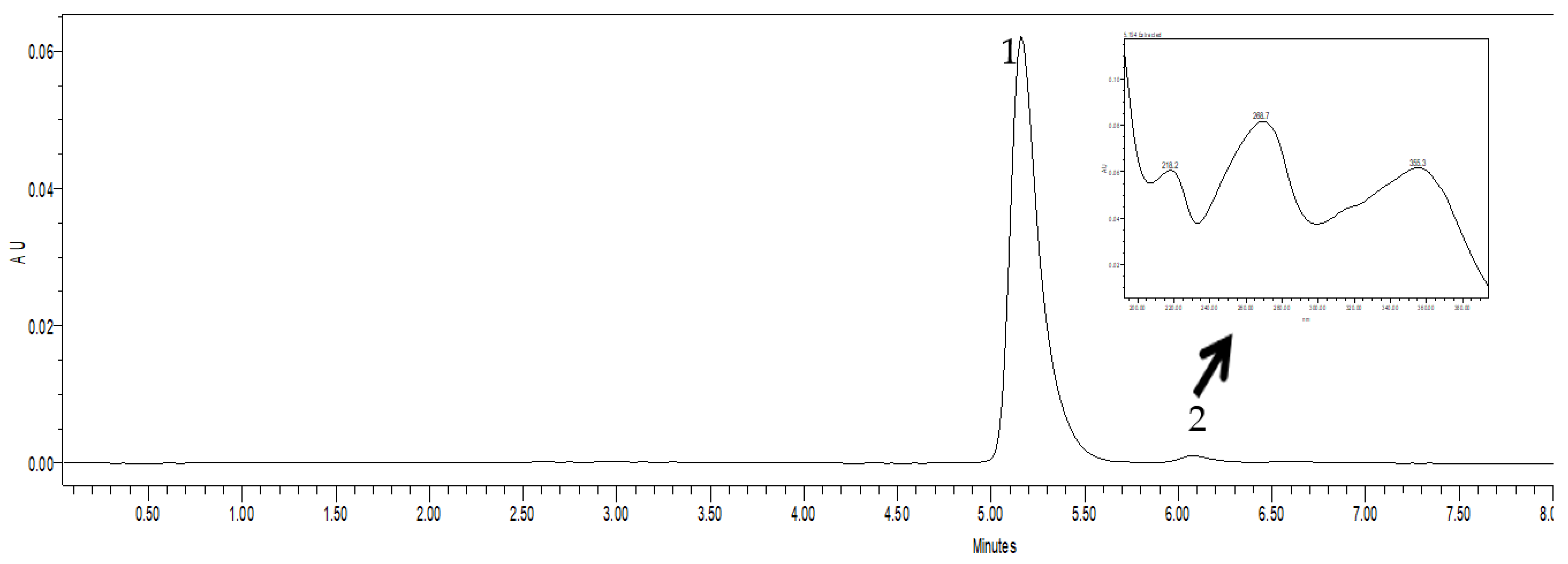
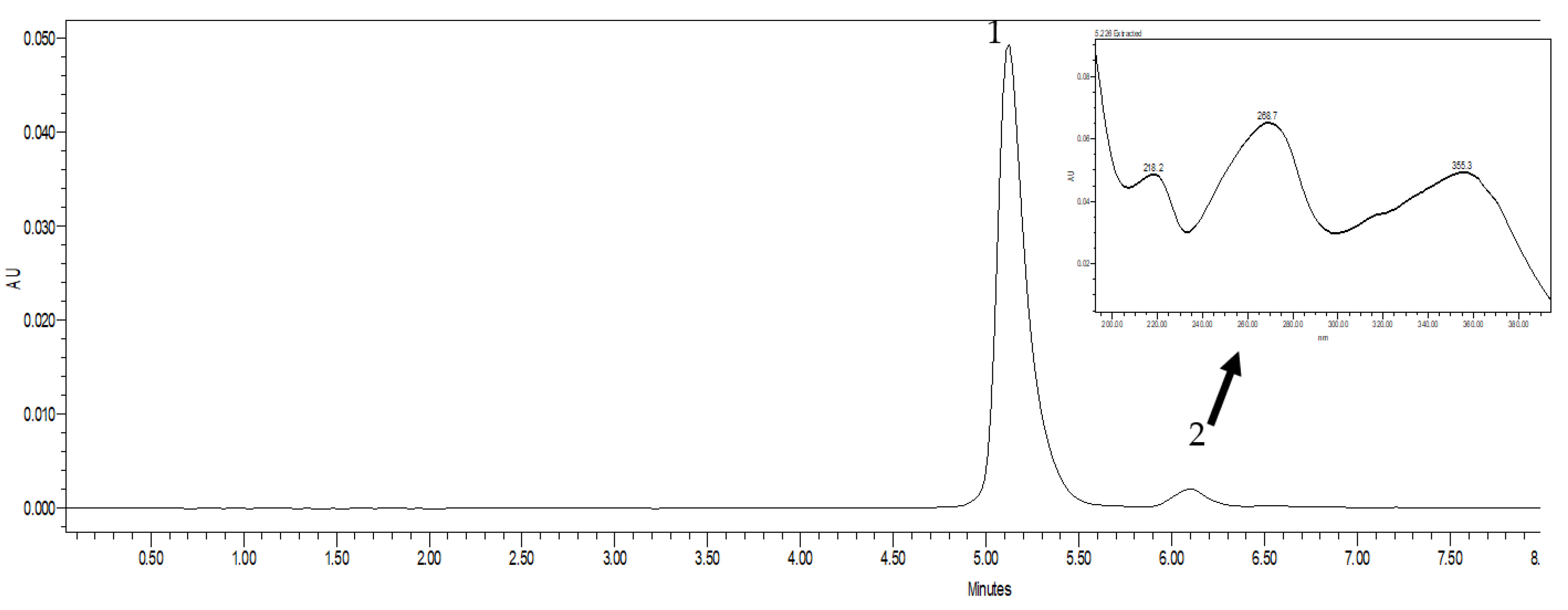
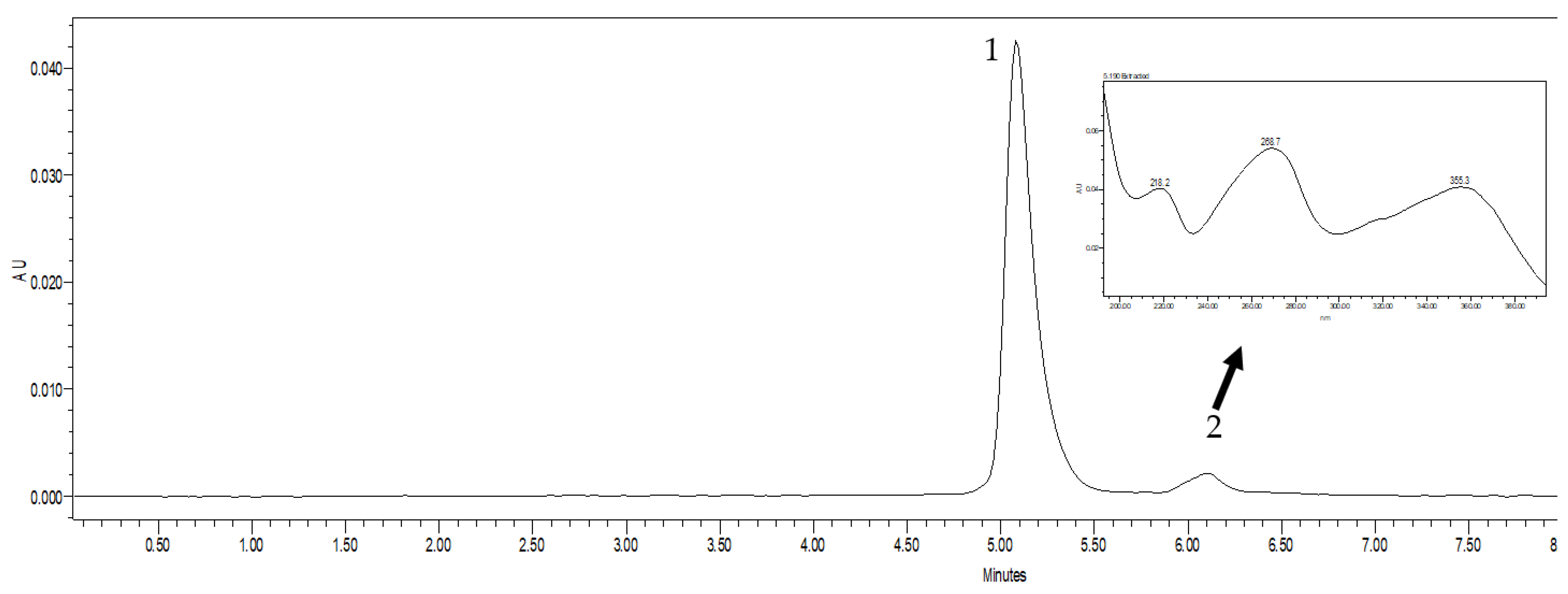
2.2. Chromatography
2.3. Oxytetracycline Susceptibility Testing
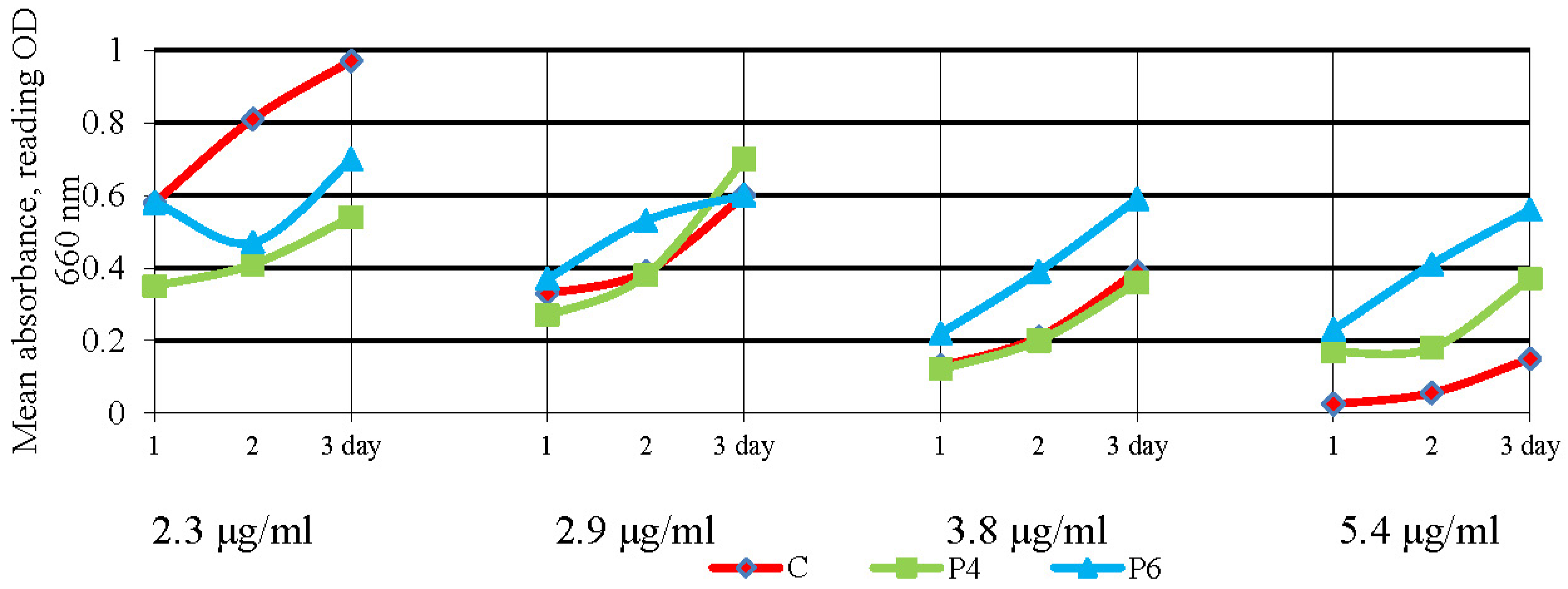
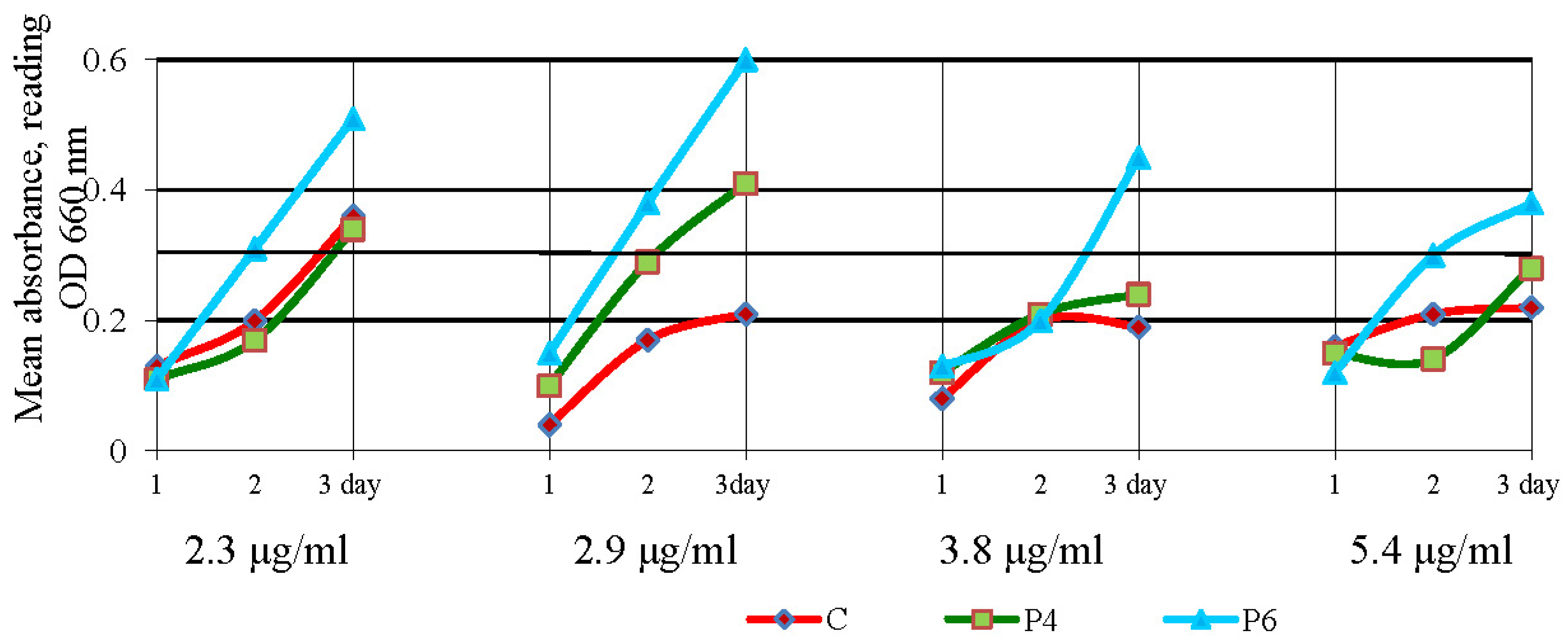
2.4. Oxytetracycline Susceptibility Testing in Liquid Medium
3. Results
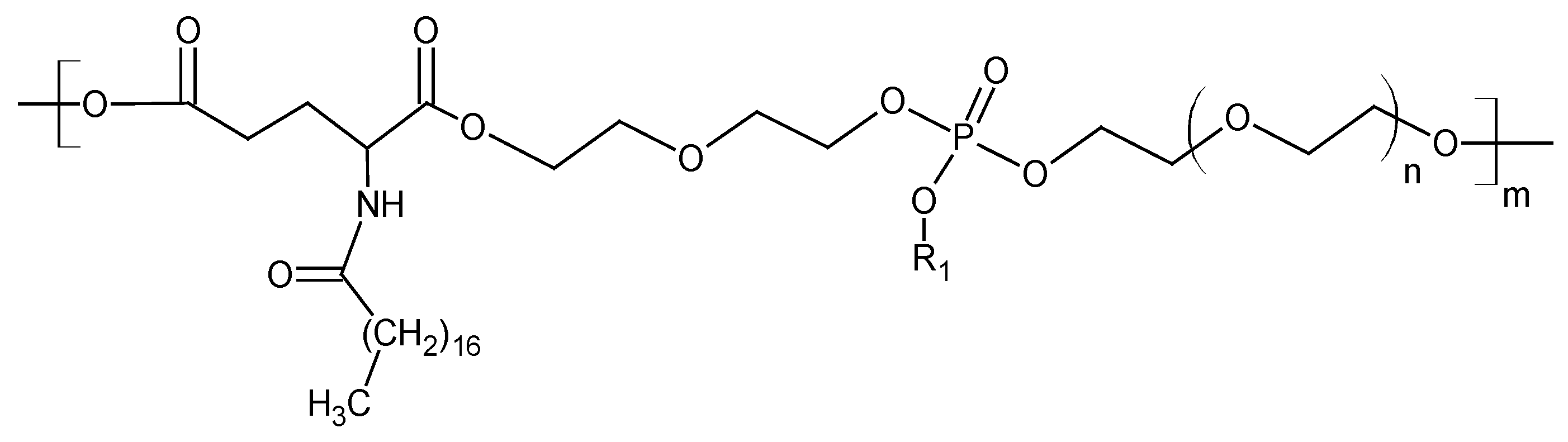
3.1. The Structure of the Polyphosphate Ester
3.2. Chromatography Studies
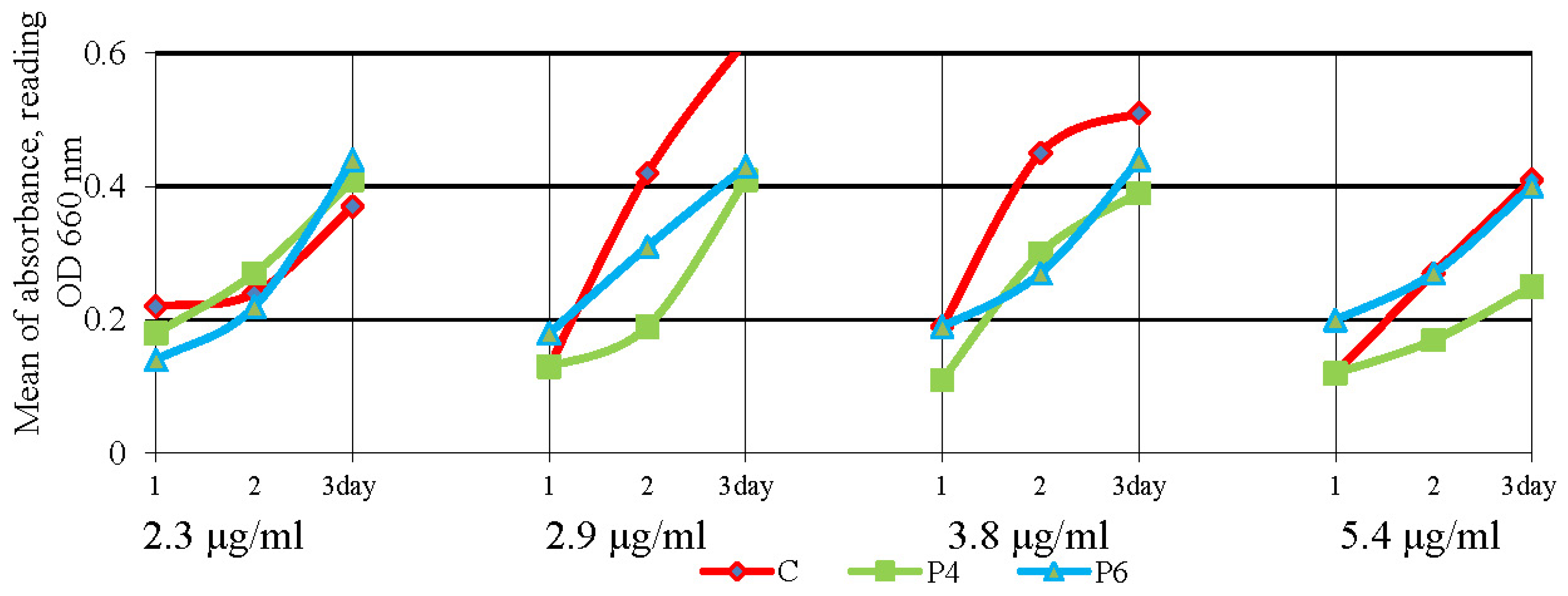
3.3. Antibacterial Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pethick, F.E.; MacFadyen, A.C.; Tang, Z.; Sangal, V.; Liu, T.-T.; Chu, J.; Kosec, G.; Petkovic, H.; Guo, M.; Kirby, R.; et al. Draft genome sequence of the oxytetracycline-producing bacterium Streptomyces rimosus ATCC. Genome Announc. 2013, 1, e00063-13. [Google Scholar] [CrossRef] [Green Version]
- Petković, H.; Cullum, J.; Hranueli, D.; Hunter, I.S.; Perić-Concha, N.; Pigac, J.; Thamchaipenet, A.; Vujaklija, D.; Long, P.F. Genetics of Streptomyces rimosus, the oxytetracycline producer. Microbiol. Mol. Biol. Rev. 2006, 70, 704–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petković, H.; Lukežič, T.; Šušković, J. Biosynthesis of Oxytetracycline by Streptomyces rimosus: Past, Present and Future Directions in the Development of Tetracycline Antibiotics. Food Technol. Biotechnol. 2017, 55, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Pickens, L.B.; Tang, Y. Oxytetracycline biosynthesis. J. Biol. Chem. 2010, 285, 27509–27515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chukwudi, C.U. rRNA Binding Sites and the Molecular Mechanism of Action of the Tetracyclines. Antimicrob. Agents Chemother. 2016, 60, 4433–4441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, M.; Achmon, Y.; Cao, Y.; Liang, X.; Chen, L.; Wang, H.; Siame, B.A.; Leung, K.Y. Distribution of antibiotic resistance genes in the environment. Environ. Pollut. 2021, 15, 117402. [Google Scholar] [CrossRef] [PubMed]
- Cummings, K.; Aprea, V.; Altier, C. Antimicrobial resistance trends among Escherichia coli isolates obtained from dairy cattle in the northeastern United States, 2004–2011. Foodborne Pathog. Dis. 2014, 11, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Breslawec, A.P.; Wang, S.; Monahan, K.N.; Barry, L.L.; Poulin, M.B. The endoglycosidase activity of Dispersin B is mediated through electrostatic interactions with cationic poly-β-(1→6)-N-acetylglucosamine. FEBS J. 2023, 290, 1049–1059. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Breslawec, A.P.; Wang, S.; Li, C.; Poulin, M.B. Anionic amino acids support hydrolysis of poly-β-(1, 6)-N-acetylglucosamine exopolysaccharides by the biofilm dispersing glycosidase Dispersin B. J. Biol. Chem. 2021, 296, 100203. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Breslawec, A.P.; Alvarez, E.; Tyrlik, M.; Li, C.; Poulin, M.B. Differential Recognition of Deacetylated PNAG Oligosaccharides by a Biofilm Degrading Glycosidase. ACS Chem. Biol. 2019, 14, 1998–2005. [Google Scholar] [CrossRef] [PubMed]
- Loayza, F.; Graham, J.P.; Trueba, G. Factors Obscuring the Role of E. coli from Domestic Animals in the Global Antimicrobial Resistance Crisis: An Evidence-Based Review. Int. J. Environ. Res. Public Health 2020, 17, 3061. [Google Scholar] [CrossRef] [PubMed]
- Muzyka, V.P. Study of antimicrobial activity based on the treatment of respiratory fluorfenikol diseases in calves. Sci. Tech. Bull. State Sci. Res. Control Inst. Vet. Med. Prod. Fodd. Addit. Inst. Anim. Biol. 2015, 16, 213–217. [Google Scholar]
- Kabanets, A.S.; Kistsiv, O.S.; Stetsko, T.I. Determination of activity and semi-lethal dose of antibacterial nifulin plus drug a. Sci. Tech. Bull. State Sci. Res. Control Inst. Vet. Med. Prod. Fodd. Addit. Inst. Anim. Biol. 2012, 13, 236–240. [Google Scholar]
- Kucheruk, M.D.; Zasekin, D.A.; Vygovskaya, L.N.; Ushkalov, V.A. Antibiotic resistance of new strains of microorganisms. Biol. Resour. Nat. Manag. 2018, 10, 205–217. [Google Scholar]
- Kalashnikov, V.O. The study baktericidy and bakteriostatic actions some antibiotic on microbe background milks cows. Sci. Rep. NULES Ukr. 2011, 6. Available online: http://www.nbuv.gov.ua/e-journals/Nd/2011_6/11kvo.pdf (accessed on 1 February 2022).
- Di Cerbo, A.; Palatucci, A.T.; Rubino, V.; Centenaro, S.; Giovazzino, A.; Fraccaroli, E.; Cortese, L.; Ruggiero, G.; Guidetti, G.; Canello, S.; et al. Toxicological Implications and Inflammatory Response in Human Lymphocytes Challenged with Oxytetracycline. Biochem. Mol. Toxicol. 2016, 30, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Guidetti, G.; Di Cerbo, A.; Giovazzino, A.; Rubino, V.; Palatucci, A.T.; Centenaro, S.; Fraccaroli, E.; Cortese, L.; Bonomo, M.G.; Ruggiero, G.; et al. In Vitro Effects of Some Botanicals with Anti-Inflammatory and Antitoxic Activity. J. Immunol. Res. 2016, 2016, 5457010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agwuh, K.N.; MacGowan, A. Pharmacokinetics and pharmacodynamics of the tetracyclines including glycylcyclines. J. Antimicrob. Chemother. 2006, 58, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Rani, K.; Paliwal, S.A. Review on targeted drug delivery: Its entire focus on advanced therapeutics and diagnostics. Sch. J. App. Med. Sci. 2014, 2, 328–331. [Google Scholar]
- Makabenta, J.M.V.; Nabawy, A.; Li, C.-H.; Schmidt-Malan, S.; Patel, R.; Rotello, V.M. Nanomaterial-based therapeutics for antibiotic-resistant bacterial infections. Nat. Rev. Microbiol. 2021, 19, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Nautiyal, U.; Kaur, C.; Goel, V.; Piarchand, N. Targeted drug delivery system: Current and novel approach. Int. J. Pharm. Med. Res. 2017, 5, 448–454. [Google Scholar]
- Stasiuk, A.V.; Fihurka, N.V.; Tarnavchyk, I.; Nosova, N.G.; Pasetto, P.; Varvarenko, S.M.; Samaryk, V.Y. Influence of structure and nature of pseudo-poly(amino acid)s on size and morphology of their particle in self-stabilized aqueous dispersions. Appl. Nanosci. 2022. [Google Scholar] [CrossRef]
- Varvarenko, S.; Samaryk, V.; Vlizlo, V.; Ostapiv, D.; Nosova, N.; Tarnachyk, I.; Fihurka, N.; Ferens, M.; Nagornyak, M.; Taras, R.; et al. Fluorescein-containing theranostics based on the pseudo-poly(amino acid)s for monitoring of drug delivery and release. Polym. J. 2015, 37, 193–199. [Google Scholar] [CrossRef]
- Stasiuk, A.; Fihurka, N.; Vlizlo, V.; Prychak, S.; Ostapiv, D.; Varvarenko, S.; Samaryk, V. Synthesis and properties of phosphorus-containing pseudo-poly(amino acid)s of polyester type based on n-derivatives of glutaminic acid. Chem. Chem. Technol. 2022, 16, 51–58. [Google Scholar] [CrossRef]
- Stasiuk, A.; Figurka, N.; Dron, I.; Bukartyk, N.; Prytchak, S.; Lyach, V.; Ostapiv, D.; Nosova, N.; Varvarenko, S.; Vlizlo, V.; et al. Phosphorus-Containing Polyester Eters Derivatives of Dibasic Natural α-Amino Acids and Polyethylene Glycols. Patent of Ukraine UA 124918 C2. 2021, 49. Available online: https://iprop-ua.com/inv/pdf/4ym0e3pt-pub-description.pdf (accessed on 4 August 2021).
- Determination of the Sensitivity of Microorganisms to Antibacterial Drugs: [Guidance] Order of the Ministry of Health of Ukraine: 05.04.2007, 167. Available online: http://mozdocs.kiev.ua/view.php?id=6958 (accessed on 5 April 2007).
- Chekh, B.; Ferens, M.; Susol, N.; Varvarenko, S.; Ostapiv, D.; Vlizlo, V. Nanopolymer GluLa-DPG-PEG600-F Can Penetrate into Cells and Deposit in Rats Body. Sci. Bull. Lesya Ukr. Volyn Natl. Univ. 2016, 12, 138–142. [Google Scholar]
- Chekh, B.O.; Ferens, M.V.; Ostapiv, D.D.; Samaryk, V.Y.; Varvarenko, S.M.; Vlizlo, V.V. Characteristics of novel polymer based on pseudo-polyamino acids GluLa-DP G-PE G600: Binding of albumin, biocompatibility, biodistribution and potential crossing the blood-brain barrier in rats. Ukr. Biochem. J. 2017, 89, 13–21. [Google Scholar]
- Chekh, B.O.; Dron, I.A.; Vynnytska, S.I.; Oleksa, V.V.; Atamaniuk, I.E.; Vlizlo, V.V. Antibacterial activity of complex of enrofloxacin with nanopolymer GluLa-DPG-PEG. Bìol. Tvarin. 2017, 19, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.C.; van der Donk, W.A. The many roles of glutamate in metabolism. J. Ind. Microbiol. Biotechnol. 2016, 43, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Jeljaszewicz, J.; Hawiger, J. The resistance to antibiotics of strains of Streptococcus viridans, Streptococcus faecalis, Escherichia coli, Pseudomonas aeruginosa, Proteus and Klebsiella isolated in Poland. Bull. World Health Organ. 1966, 35, 243–246. [Google Scholar] [PubMed]
- Kotsyumbas, I.Y.; Musyka, V.P.; Stetsko, T.I. The state of antibiotic resistance of microorganisms—Pathogens of bacterial diseases of young cattle and pigs. Sci. Bull. Vet. Med. 2014, 13, 117–120. [Google Scholar]
- Ghosh, A.; Jayaraman, N.; Chatterji, D. Small-molecule inhibition of bacterial biofilm. ACS Omega 2020, 5, 3108–3115. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxytetracycline, μg/Disk | Area of Growth Inhibition, cm2 | |||
|---|---|---|---|---|
| Control (Oxytetracycline, Hydrochloride) | Oxytetracycline + P4 | Oxytetracycline + P6 | η2 | |
| 5.4 | 3.00 ± 0.25 | 3.60 ± 0.18 | 3.33 ± 0.24 | 0.232 |
| 3.8 | 2.90 ± 0.12 | 2.95 ± 0.12 | 3.28 ± 0.21 | 0.759 |
| 2.9 | 1.88 ± 0.17 | 2.78 ± 0.16 *** | 1.90 ± 0.06 | 0.776 |
| 2.3 | 1.58 ± 0.07 | 2.40 ± 0.11 ** | 1.58 ± 0.06 | 0.836 |
| Oxytetracycline, μg/Disk | Area of Growth Inhibition, cm2 | |||
|---|---|---|---|---|
| Control (Oxytetracycline, Hydrochloride) | Oxytetracycline + P4 | Oxytetracycline + P6 | η2 | |
| 20 | 2.23 ± 0.216 | 2.39 ± 0.125 | 1.90 ± 0.093 | 0.369 |
| 14 | 1.85 ± 0.102 | 2.46 ± 0.163 | 1.84 ± 0.135 | 0.605 |
| 11 | 1.76 ± 0.109 | 1.75 ± 0.051 | 1.46 ± 0.031 | 0.559 |
| 8 | 1.20 ± 0.036 | 1.69 ± 0.128 * | 1.55 ± 0.026 *** | 0.695 |
| Oxytetracycline, μg/Disk | Area of Growth Inhibition, cm2 | |||
|---|---|---|---|---|
| Control (Oxytetracycline, Hydrochloride) | Oxytetracycline + P4 | Oxytetracycline + P6 | η2 | |
| 20 | 3.10 ± 0.12 | 2.10 ± 0.12 | 1.80 ± 0.05 | 0.903 |
| 14 | 2.63 ± 0.31 | 1.87 ± 0.09 | 1.67 ± 0.12 | 0.587 |
| 11 | 1.97 ± 0.14 | 1.73 ± 0.09 | 1.60 ± 0.08 | 0.383 |
| 8 | 1.20 ± 0.05 | 1.33 ± 0.07 | 1.27 ± 0.07 | 0.190 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozak, M.; Stasiuk, A.; Vlizlo, V.; Ostapiv, D.; Bodnar, Y.; Kuz’mina, N.; Figurka, N.; Nosova, N.; Ostapiv, R.; Kotsumbas, I.; et al. Polyphosphate Ester-Type Transporters Improve Antimicrobial Properties of Oxytetracycline. Antibiotics 2023, 12, 616. https://doi.org/10.3390/antibiotics12030616
Kozak M, Stasiuk A, Vlizlo V, Ostapiv D, Bodnar Y, Kuz’mina N, Figurka N, Nosova N, Ostapiv R, Kotsumbas I, et al. Polyphosphate Ester-Type Transporters Improve Antimicrobial Properties of Oxytetracycline. Antibiotics. 2023; 12(3):616. https://doi.org/10.3390/antibiotics12030616
Chicago/Turabian StyleKozak, Mariya, Anna Stasiuk, Vasyl Vlizlo, Dmytro Ostapiv, Yulia Bodnar, Nataliia Kuz’mina, Natalia Figurka, Natalia Nosova, Roman Ostapiv, Igor Kotsumbas, and et al. 2023. "Polyphosphate Ester-Type Transporters Improve Antimicrobial Properties of Oxytetracycline" Antibiotics 12, no. 3: 616. https://doi.org/10.3390/antibiotics12030616