
Efficient AntiMycolata Agents by Increasing the Lipophilicity of Known Antibiotics through Multicomponent Reactions

 ,
, 
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results & Discussion
2.1. Chemical Synthesis
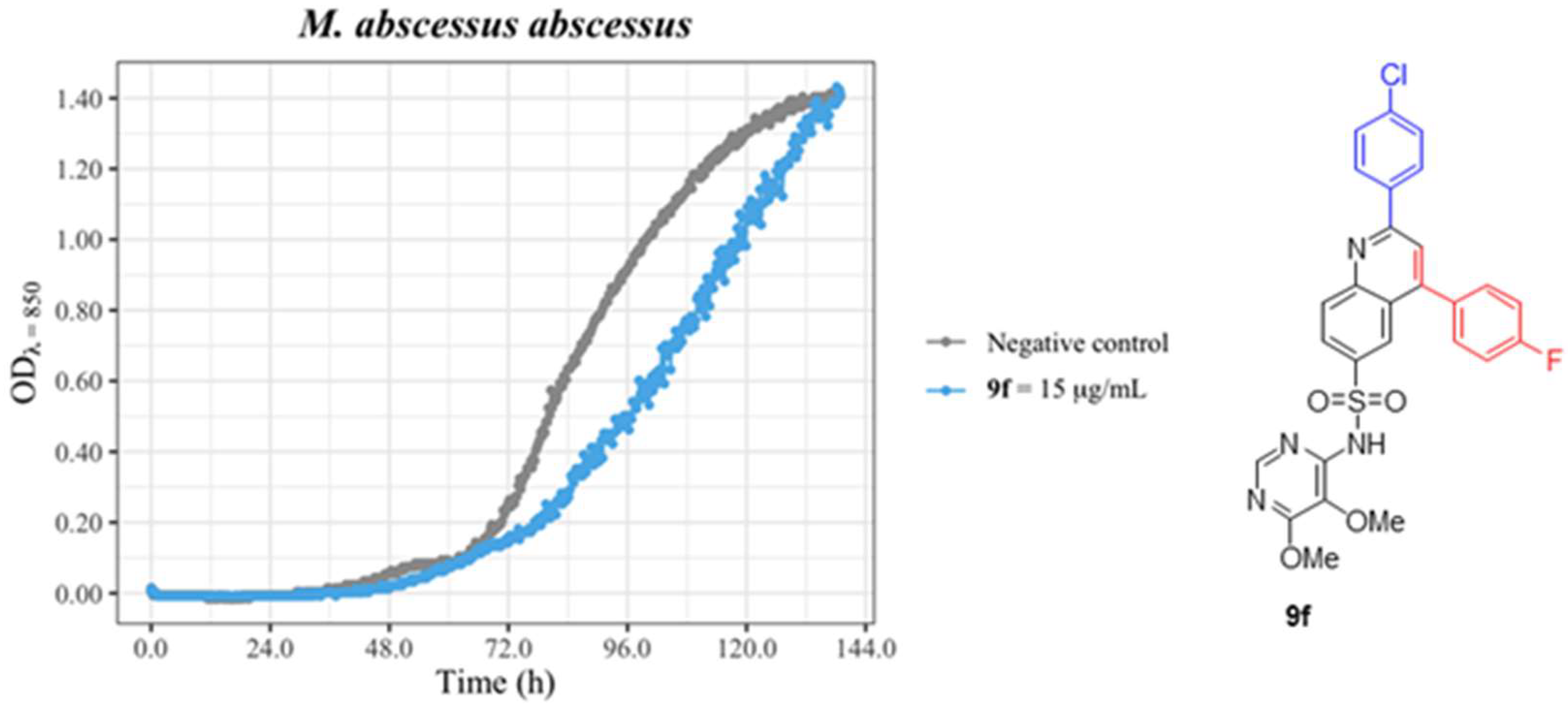
2.2. Microbiological Studies
3. Materials and Methods
3.1. Chemical Synthesis
3.2. Culture Conditions
3.3. Antimicrobial Compounds
3.4. Susceptibility Measurements
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brennan, P.J.; Nikaido, H. The envelope of mycobacteria. Annu. Rev. Biochem. 1995, 64, 29–63. [Google Scholar] [CrossRef] [PubMed]
- Lichtinger, T.; Burkovski, A.; Niederweis, M.; Krämer, R.; Benz, R. Biochemical and Biophysical Characterization of the Cell Wall Porin of Corynebacterium glutamicum: The Channel Is Formed by a Low Molecular Mass Polypeptide. Biochemistry 1998, 37, 15024–15032. [Google Scholar] [CrossRef]
- Rieß, F.G.; Lichtinger, T.; Yassin, A.F.; Schaal, K.P.; Benz, R. The cell wall porin of the gram-positive bacterium Nocardia asteroides forms cation-selective channels that exhibit asymmetric voltage dependence. Arch. Microbiol. 1999, 171, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Jarlier, V.; Nikaido, H. Mycobacterial cell wall: Structure and role in natural resistance to antibiotics. FEMS Microbiol. Lett. 1994, 123, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Jarlier, V. Permeability of the mycobacterial cell wall. Res. Microbiol. 1991, 142, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Iida, S.; Taniguchi, H.; Kageyama, A.; Yazawa, K.; Chibana, H.; Murata, S.; Nomura, F.; Kroppenstedt, R.M.; Mikami, Y. Gordonia otitidis sp. nov., isolated from a patient with external otitis. Int. J. Syst. Evol. Microbiol. 2005, 55, 1871–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenskötter, M.; Bröker, D.; Steinbüchel, A. Biology of the metabolically diverse genus Gordonia. Appl. Environ. Microbiol. 2004, 70, 3195–3204. [Google Scholar] [CrossRef] [Green Version]
- Drzyzga, O. The strengths and weaknesses of Gordonia: A review of an emerging genus with increasing biotechnological potential. Crit. Rev. Microbiol. 2012, 38, 300–316. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Rainey, F.A.; Ward-Rainey, N.L. Proposal for a New Hierarchic Classification System, Actinobacteria classis nov. Int. J. Syst. Bacteriol. 1997, 47, 479–491. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, N.; Toumeh, A.; Georgescu, C. Tibial osteomyelitis caused by Gordonia bronchialis in an immunocompetent patient. J. Clin. Microbiol. 2012, 50, 3119–3121. [Google Scholar] [CrossRef]
- Werno, A.M.; Anderson, T.P.; Chambers, S.T.; Laird, H.M.; Murdoch, D.R. Recurrent breast abscess caused by Gordonia bronchialis in an immunocompetent patient. J. Clin. Microbiol. 2005, 43, 3009–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Miguel, T.; Sieiro, C.; Poza, M.; Villa, T.G. Isolation and taxonomic study of a new canthaxanthin-containing bacterium, Gordonia jacobaea MV-1 sp. nov. Int. Microbiol. 2000, 3, 107–111. [Google Scholar] [PubMed]
- Jiménez-Galisteo, G.; Villa, T.G.; Vinuesa, T.; Vinas, M.; Domínguez, A.; Muñoz, E. Draft Genome Sequence of the Bacterium Gordonia jacobaea, a New Member of the Gordonia Genus. Genome Announc. 2015, 3, e00995-15. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Galisteo, G.; Fusté, E.; Muñoz, E.; Vinuesa, T.; Villa, T.G.; Benz, R.; Domínguez, A.; Viñas, M. Identification and characterization of a cell wall porin from Gordonia jacobaea. J. Gen. Appl. Microbiol. 2017, 63, 266–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiraud, J.; Lescure, M.; Faganello, D.; Bebear, C.; Pereyre, S.; Menard, A. A case of prosthetic joint septic arthritis caused by Gordonia jacobaea. J. Microbiol. Immunol. Infect. 2022, 55, 355–357. [Google Scholar] [CrossRef]
- Kageyama, A.; Hoshino, Y.; Yazawa, K.; Poonwan, N.; Takeshita, N.; Maki, S.; Mikami, Y. Nocardia cyriacigeorgica is a significant pathogen responsible for nocardiosis in Japan and Thailand. Mycopathologia 2005, 160, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Portolá, O.; Guitart, R.; Gómez, F.; Olona, M.; Vidal, F.; Castro, A. Epidemiología y manifestaciones clínicas de la infección por especies de Nocardia en Tarragona, 1997 a 2008: Nocardia cyriacigeorgica es un patógeno emergente. Enferm. Infecc. Microbiol. Clin. 2009, 27, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Schlaberg, R.; Huard, R.C.; Della-Latta, P. Nocardia cyriacigeorgica, an emerging pathogen in the United States. J. Clin. Microbiol. 2008, 46, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Fatahi-Bafghi, M. Nocardiosis from 1888 to 2017. Microb. Pathog. 2018, 114, 369–384. [Google Scholar] [CrossRef]
- Honda, J.R.; Virdi, R.; Chan, E.D. Global Environmental Nontuberculous Mycobacteria and Their Contemporaneous Man-Made and Natural Niches. Front. Microbiol. 2018, 9, 2029. [Google Scholar] [CrossRef]
- Nessar, R.; Cambau, E.; Reyrat, J.M.; Murray, A.; Gicquel, B. Mycobacterium abscessus: A new antibiotic nightmare. J. Antimicrob. Chemother. 2012, 67, 810–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cernak, T.; Dykstra, K.D.; Tyagarajan, S.; Vachal, P.; Krska, S.W. The Medicinal Chemist’s Toolbox for Late-Stage Function-alization of Drug-like Molecules. Chem. Soc. Rev. 2016, 45, 546–576. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the sustainable discovery and development of new antibiotics. Nat. Rev. Chem. 2021, 5, 726–749. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Weng, X.; Li, Y.; Su, M.; Wen, Z.; Ji, X.; Ren, N.; Shen, B.; Duan, Y.; Huang, Y. Late-Stage Functionalization of Platensimycin Leading to Multiple Analogues with Improved Antibacterial Activity In Vitro and In Vivo. J. Med. Chem. 2019, 62, 6682–6693. [Google Scholar] [CrossRef]
- Dailler, D.; Dorst, A.; Schäfle, D.; Sander, P.; Gademann, K. Novel fidaxomicin antibiotics through site-selective catalysis. Commun. Chem. 2021, 4, 59. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, Z.-P.; Lin, Z.; Wei, G.; Wen, X.; Li, S.; Yang, X.; Zhang, Q.; Jing, C.; Dai, Y.; et al. Drug Repurposing by Siderophore Conjugation: Synthesis and Biological Evaluation of Siderophore-Methotrexate Conjugates as Antibiotics. Angew. Chem. Int. Ed. 2022, 61, e202204139. [Google Scholar]
- Zhu, J.; Bienaymé, H. (Eds.) Multicomponent Reactions; Wiley-VCH: Weinheim, Germany, 2005. [Google Scholar]
- Boltjes, A.; Dömling, A. The Groebke-Blackburn-Bienaymé Reaction. Eur. J. Org. Chem. 2019, 2019, 7007–7049. [Google Scholar] [CrossRef]
- Pedrola, M.; Jorba, M.; Jardas, E.; Jardi, F.; Ghashghaei, O.; Viñas, M.; Lavilla, R. Multicomponent Reactions Upon the Known Drug Trimethoprim as a Source of Novel Antimicrobial Agents. Front. Chem. 2019, 7, 475. [Google Scholar] [CrossRef]
- Jorba, M.; Pedrola, M.; Ghashghaei, O.; Herráez, R.; Campos-Vicens, L.; Luque, F.J.; Lavilla, R.; Viñas, M. New Trimethoprim-Like Molecules: Bacteriological Evaluation and Insights into Their Action. Antibiotics 2021, 10, 709. [Google Scholar] [CrossRef]
- Lipinski, C.; Hopkins, A. Navigating Chemical Space for Biology and Medicine. Nature 2004, 432, 855–861. [Google Scholar] [CrossRef]
- Dömling, A.; Wang, W.; Wang, K. Chemistry and Biology of Multicomponent Reactions. Chem. Rev. 2012, 112, 3083–3135. [Google Scholar] [CrossRef] [PubMed]
- Domling, A.; Ugi, I. Multicomponent reactions with isocyanides. Angew. Chem. Int. Ed. 2000, 39, 3168–3210. [Google Scholar] [CrossRef]
- Kouznetsov, V. Recent Synthetic Developments in a Powerful Imino Diels-Alder Reaction (Povarov Reaction): Application to the Synthesis of N-Polyheterocycles and Related Alkaloids. Tetrahedron 2008, 65, 2721–2750. [Google Scholar] [CrossRef]
- Ghashghaei, O.; Masdeu, C.; Alonso, C.; Palacios, F.; Lavilla, R. Recent Advances of the Povarov Reaction in Medicinal Chemistry. Drug Discov. Today Technol. 2018, 29, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Fuertes, M.; Martín-Encinas, E.; Selas, A.; Rubiales, G.; Tesauro, C.; Knudsen, B.R.; Palacios, F. Novel topoisomerase I inhibitors. Syntheses and biological evaluation of phosphorus substituted quinoline derivates with antiproliferative activity. Eur. J. Med. Chem. 2018, 149, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Selas, A.; Fuertes, M.; Melcón-Fernández, E.; Pérez-Pertejo, Y.; Reguera, R.M.; Balaña-Fouce, R.; Knudsen, B.R.; Palacios, F.; Alonso, C. Hybrid Quinolinyl Phosphonates as Heterocyclic Carboxylate Isosteres: Synthesis and Biological Evaluation against Topoisomerase 1B (TOP1B). Pharmaceuticals 2021, 14, 784. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Ishitani, H.; Nagayama, S. Ln(OTf)3- or Sc(OTf)3-catalyzed Three components coupling reactions between al-dehydes, amines, and dienes or alkenes. Efficient syntheses of pyridine and quinoline derivatives. Chem. Lett. 1995, 24, 423–424. [Google Scholar] [CrossRef]
- Yamamoto, S.; Zhou, Z.Y.; Hiruta, G.; Takeuchi, K.; Choi, J.-C.; Yasuda, T.; Kanbara, T.; Kuwabara, J.J. One-Pot Synthesis of Triazatriphenylene Using the Povarov Reaction. J. Org. Chem. 2021, 86, 7920–7927. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, Z. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Kaplan, D.; Pruim, R. ggformula: Formula Interface to the Grammar of Graphics, R Package Version 0.10.2 edn. Available online: https://rdrr.io/cran/ggformula/ (accessed on 7 December 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 9783319242774. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | G. jacobaea | N. cyriacigeorgica | M. abscessus |
|---|---|---|---|
| 1a | - | - | - |
| 1b | - | - | - |
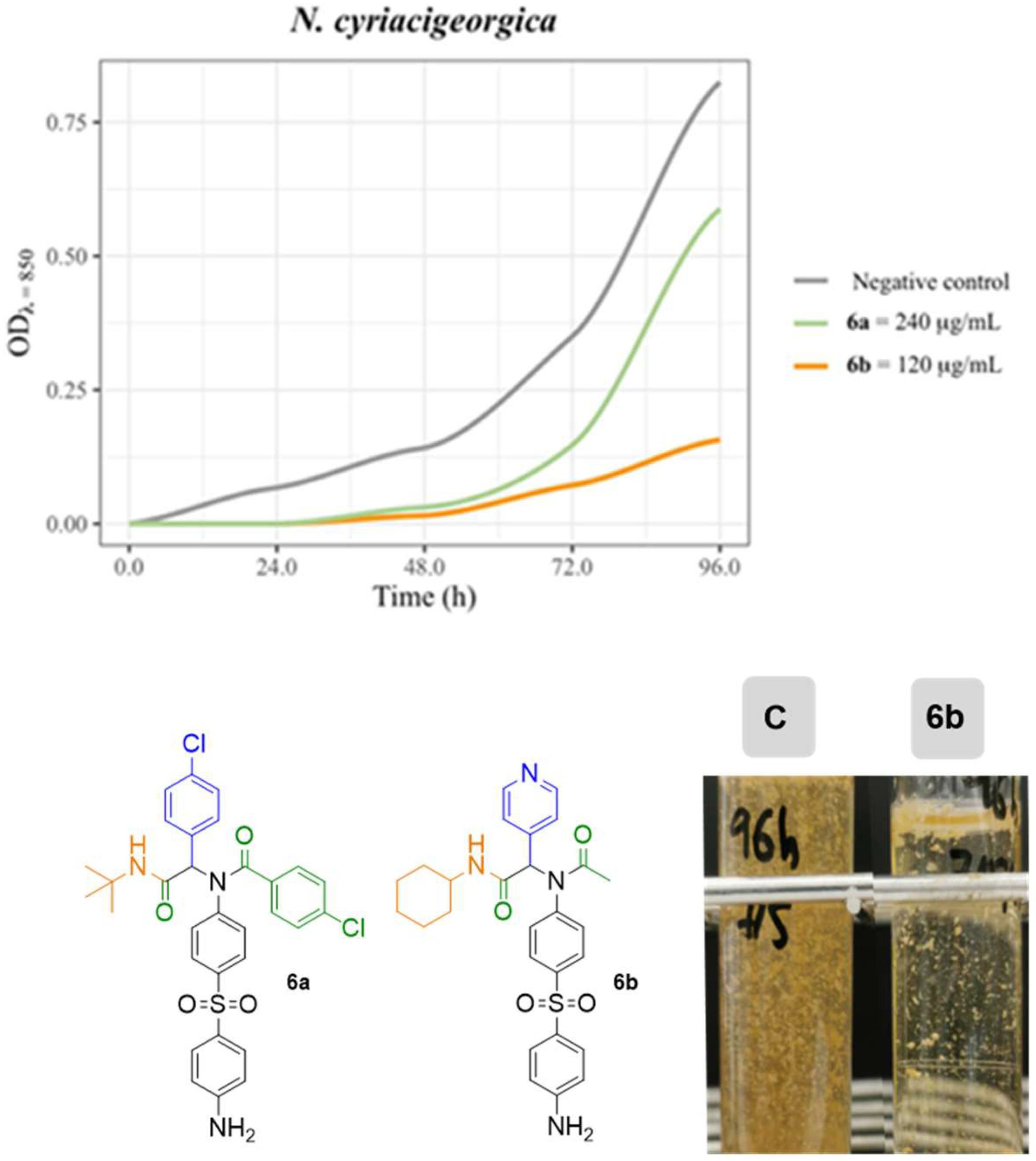
| 6a | 9.0 mm | 11.0 mm | - |
| 6b | - | - | - |
| 6c | - | - | - |
| 7a | - | 10.0 mm | - |
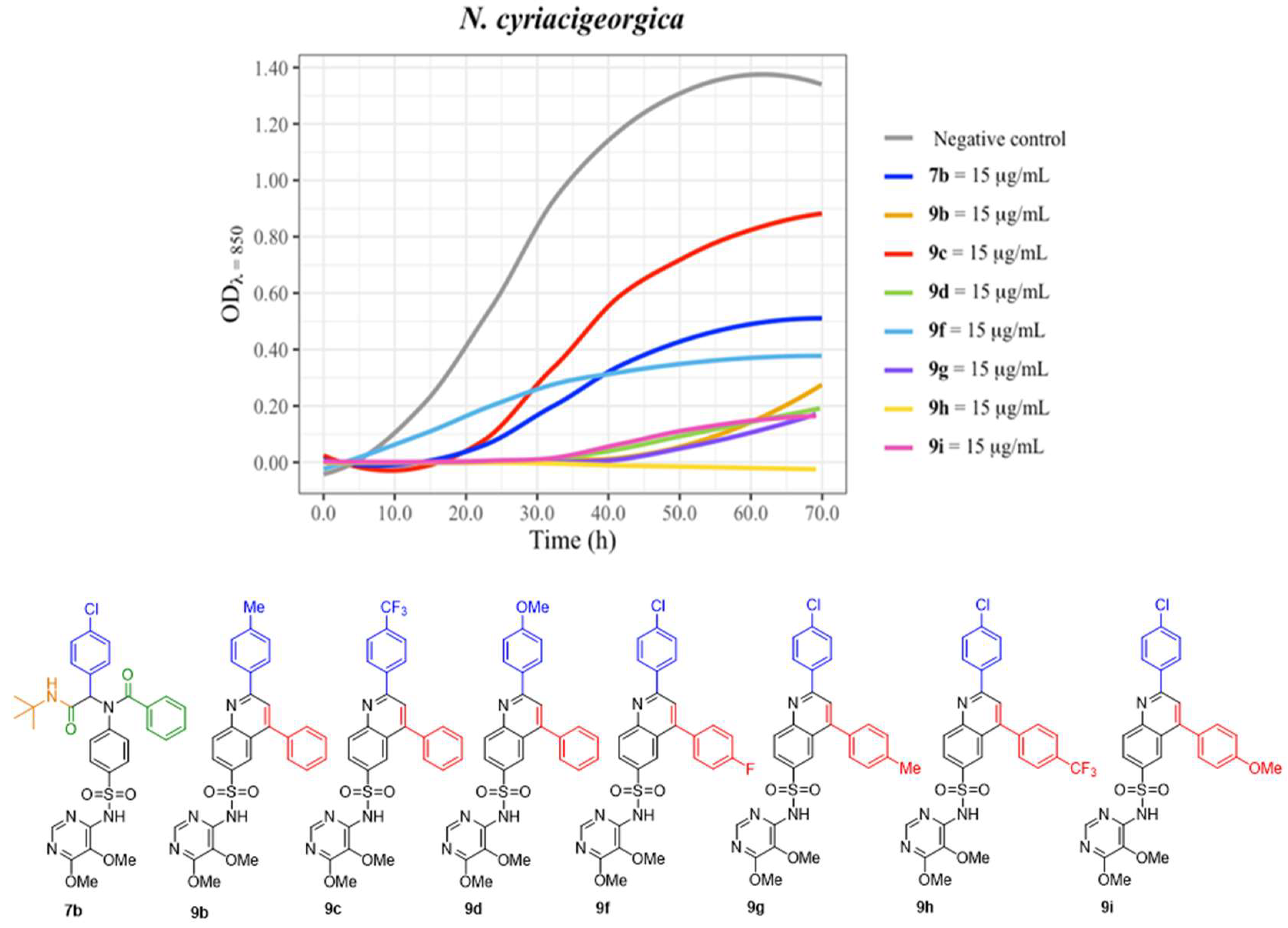
| 7b | - | 10.0 mm | - |
| 9a | - | - | - |
| 9b | - | 13.0 mm | - |
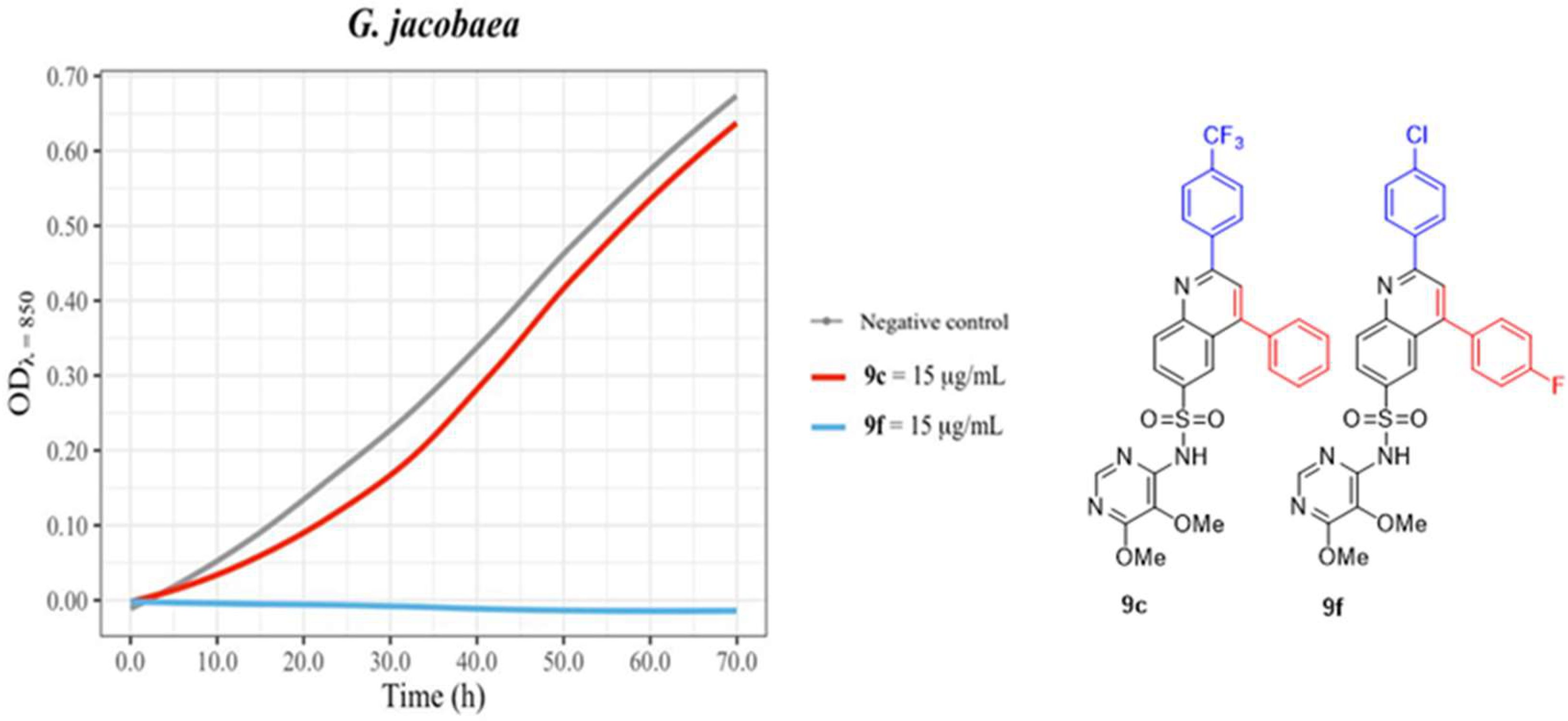
| 9c | - | 15.0 mm | - |
| 9d | 11.0 mm | - | |
| 9e | - | - | - |
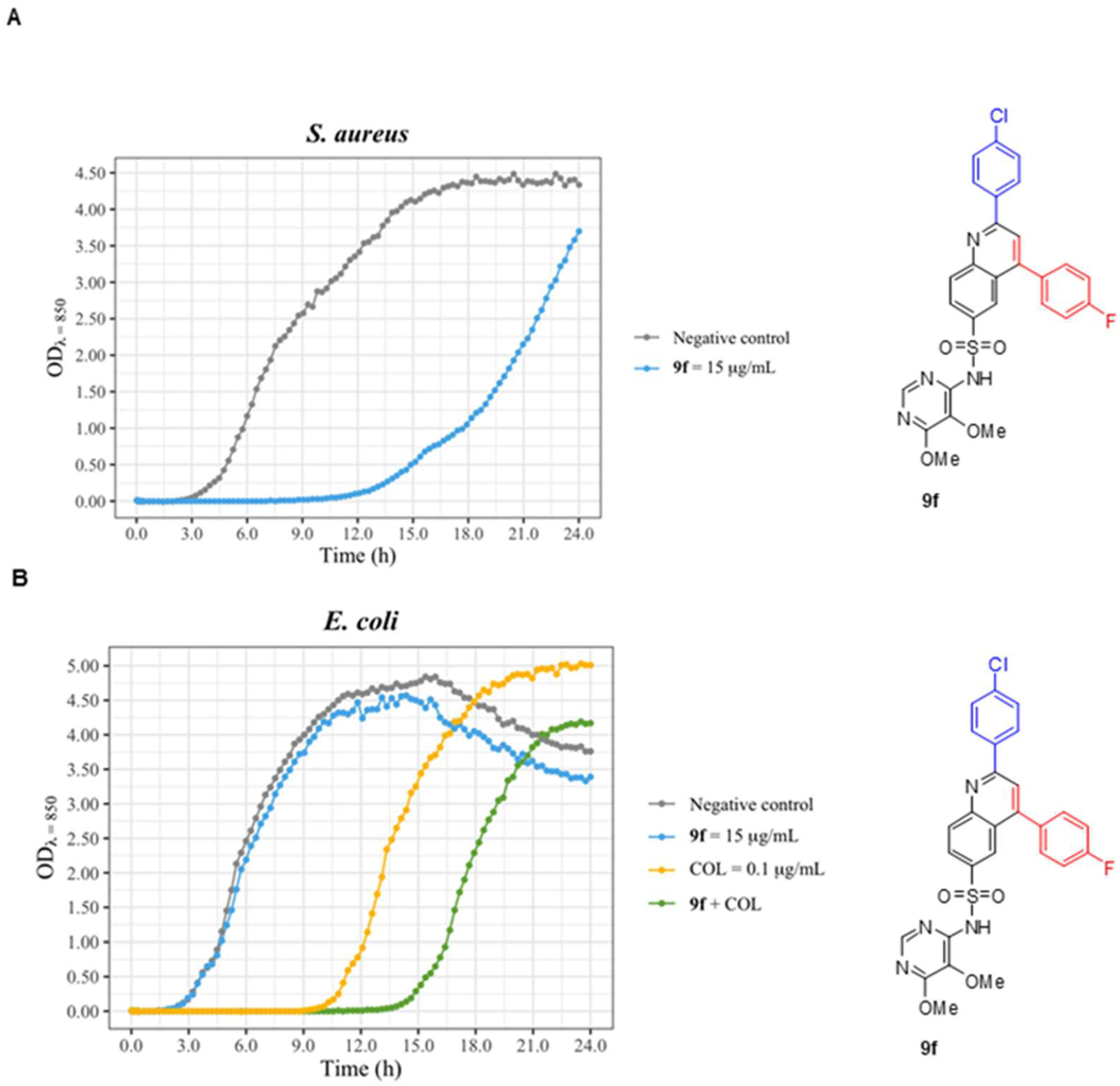
| 9f | - | 18.0 mm | - |
| 9g | - | 11.5 mm | - |
| 9h | - | 8.5 mm | - |
| 9i | - | 14.0 mm | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trejo, A.; Masdeu, C.; Serrano-Pérez, I.; Pedrola, M.; Juanola, N.; Ghashghaei, O.; Jiménez-Galisteo, G.; Lavilla, R.; Palacios, F.; Alonso, C.; et al. Efficient AntiMycolata Agents by Increasing the Lipophilicity of Known Antibiotics through Multicomponent Reactions. Antibiotics 2023, 12, 83. https://doi.org/10.3390/antibiotics12010083
Trejo A, Masdeu C, Serrano-Pérez I, Pedrola M, Juanola N, Ghashghaei O, Jiménez-Galisteo G, Lavilla R, Palacios F, Alonso C, et al. Efficient AntiMycolata Agents by Increasing the Lipophilicity of Known Antibiotics through Multicomponent Reactions. Antibiotics. 2023; 12(1):83. https://doi.org/10.3390/antibiotics12010083
Chicago/Turabian StyleTrejo, Angela, Carme Masdeu, Irene Serrano-Pérez, Marina Pedrola, Narcís Juanola, Ouldouz Ghashghaei, Guadalupe Jiménez-Galisteo, Rodolfo Lavilla, Francisco Palacios, Concepción Alonso, and et al. 2023. "Efficient AntiMycolata Agents by Increasing the Lipophilicity of Known Antibiotics through Multicomponent Reactions" Antibiotics 12, no. 1: 83. https://doi.org/10.3390/antibiotics12010083