
Clonal Dissemination of Extended-Spectrum Cephalosporin-Resistant Enterobacterales between Dogs and Humans in Households and Animal Shelters of Romania
 , ,
, ,
Abstract
:1. Introduction
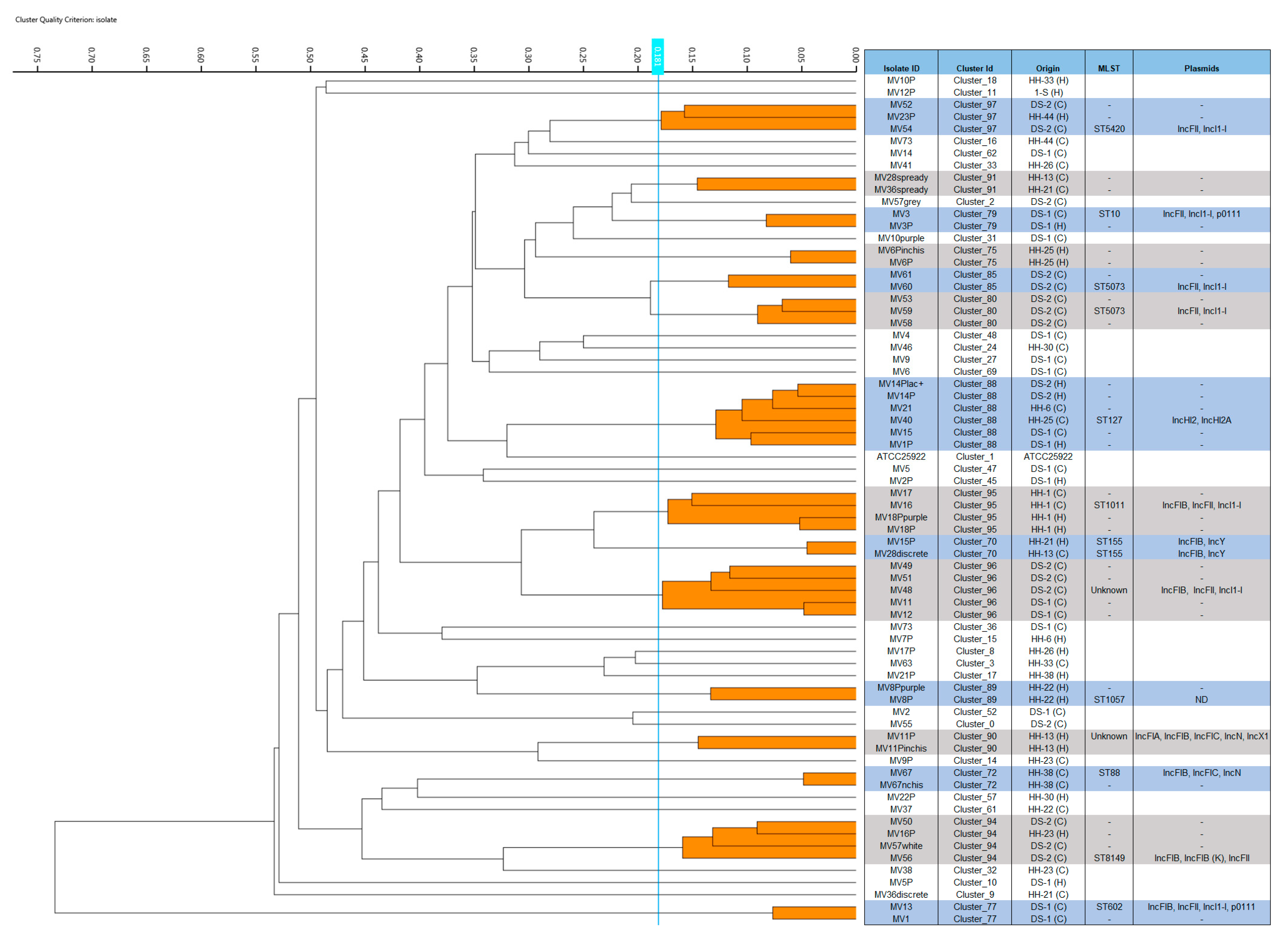
2. Results
2.1. Study Groups and Bacterial Isolates
2.2. Characterization of ESBL and Other Resistance Genes
2.3. Phylogroup and PCR-Based Replicon Typing
2.4. Fourier Transform Infrared Spectroscopy (FTIR) Typing, Multi-Locus Sequence Typing (MLST), ESC-R/ESBL Resistance Genes, and Plasmid Cluster Traits
2.5. Co-Carriage of ESC-R Isolates in Dogs and Humans from Households and Dog Shelters
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Bacterial Isolates
4.3. Antimicrobial Susceptibility Testing
4.4. Characterization of Beta-Lactamase and FQ Resistance Genes
4.5. Molecular Characterization of E. coli Genetic Background and PCR-Based Replicon Typing
4.6. Fourier Transform InfraRed Spectroscopy (FTIR); Sample Preparation and Analysis
4.7. Whole Genome Sequencing (WGS) Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rubin, J.E.; Pitout, J.D.D. Extended-spectrum beta-lactamase, carbapenemase and AmpC producing Enterobacteriaceae in companion animals. Vet. Microbiol. 2014, 170, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Coque, T.M.; Baquero, F.; Canton, R. Increasing prevalence of ESBL-producing Enterobacteriaceae in Europe. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. = Eur. Commun. Dis. Bull. 2008, 13, 19044. [Google Scholar] [CrossRef]
- Salgado-Caxito, M.; Benavides, J.A.; Adell, A.D.; Paes, A.C.; Moreno-Switt, A.I. Global prevalence and molecular characterization of extended-spectrum β-lactamase producing-Escherichia coli in dogs and cats—A scoping review and meta-analysis. One Health 2021, 12, 100236. [Google Scholar] [CrossRef]
- Strahilevitz, J.; Jacoby, G.A.; Hooper, D.C.; Robicsek, A. Plasmid-Mediated Quinolone Resistance: A Multifaceted Threat. Clin. Microbiol. Rev. 2009, 22, 664–689. [Google Scholar] [CrossRef]
- Petty, N.K.; Ben Zakour, N.L.; Stanton-Cook, M.; Skippington, E.; Totsika, M.; Forde, B.M.; Phan, M.D.; Gomes Moriel, D.; Peters, K.M.; Davies, M.; et al. Global dissemination of a multidrug resistant Escherichia coli clone. Proc. Natl. Acad. Sci. USA 2014, 111, 5694–5699. [Google Scholar] [CrossRef]
- Ewers, C.; Bethe, A.; Stamm, I.; Grobbel, M.; Kopp, P.A.; Guerra, B.; Stubbe, M.; Doi, Y.; Zong, Z.; Kola, A.; et al. CTX-M-15-D-ST648 Escherichia coli from companion animals and horses: Another pandemic clone combining multiresistance and extraintestinal virulence? J. Antimicrob. Chemother. 2014, 69, 1224–1230. [Google Scholar] [CrossRef]
- Zogg, A.L.; Zurfluh, K.; Schmitt, S.; Nüesch-Inderbinen, M.; Stephan, R. Antimicrobial resistance, multilocus sequence types and virulence profiles of ESBL producing and non-ESBL producing uropathogenic Escherichia coli isolated from cats and dogs in Switzerland. Vet. Microbiol. 2018, 216, 79–84. [Google Scholar] [CrossRef]
- Dierikx, C.M.; van Duijkeren, E.; Schoormans, A.H.; van Essen-Zandbergen, A.; Veldman, K.; Kant, A.; Huijsdens, X.W.; van der Zwaluw, K.; Wagenaar, J.A.; Mevius, D.J. Occurrence and characteristics of extended-spectrum-β-lactamase- and AmpC-producing clinical isolates derived from companion animals and horses. J. Antimicrob. Chemother. 2012, 67, 1368–1374. [Google Scholar] [CrossRef]
- Melo, L.C.; Oresco, C.; Leigue, L.; Netto, H.M.; Melville, P.A.; Benites, N.R.; Saras, E.; Haenni, M.; Lincopan, N.; Madec, J.Y. Prevalence and molecular features of ESBL/pAmpC-producing Enterobacteriaceae in healthy and diseased companion animals in Brazil. Vet. Microbiol. 2018, 221, 59–66. [Google Scholar] [CrossRef]
- Timofte, D.; Maciuca, I.E.; Williams, N.J.; Wattret, A.; Schmidt, V. Veterinary Hospital Dissemination of CTX-M-15 Extended-Spectrum Beta-Lactamase-Producing Escherichia coli ST410 in the United Kingdom. Microb. Drug Resist. 2016, 22, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Bortolami, A.; Zendri, F.; Maciuca, E.I.; Wattret, A.; Ellis, C.; Schmidt, V.; Pinchbeck, G.; Timofte, D. Diversity, Virulence, and Clinical Significance of Extended-Spectrum β-Lactamase- and pAmpC-Producing Escherichia coli from Companion Animals. Front. Microbiol. 2019, 10, 1260. [Google Scholar] [CrossRef] [PubMed]
- Baede, V.O.; Wagenaar, J.A.; Broens, E.M.; Duim, B.; Dohmen, W.; Nijsse, R.; Timmerman, A.J.; Hordijk, J. Longitudinal study of extended-spectrum-β-lactamase- and AmpC-producing Enterobacteriaceae in household dogs. Antimicrob. Agents Chemother. 2015, 59, 3117–3124. [Google Scholar] [CrossRef] [PubMed]
- Hordijk, J.; Schoormans, A.; Kwakernaak, M.; Duim, B.; Broens, E.; Dierikx, C.; Mevius, D.; Wagenaar, J.A. High prevalence of fecal carriage of extended spectrum β-lactamase/AmpC-producing Enterobacteriaceae in cats and dogs. Front. Microbiol. 2013, 4, 242. [Google Scholar] [CrossRef]
- European Public Health Alliance. In the Red Zone—Antimicrobial Resistance: Lessons from Romania. Brussels, March 2017. Available online: https://epha.org/wp-content/uploads/2017/06/In-the-red-zone-EPHA.pdf (accessed on 1 July 2022).
- FEDIAF. Share of Households Owning at Least One Dog in the European Union in 2020, by Country [Graph]. In Statista. November 23, 2021. Available online: https://www.statista.com/statistics/515475/dog-ownership-european-union-eu-by-country/ (accessed on 19 July 2022).
- European Centre for Disease Prevention and Control; World Health Organization, Regional Office for Europe. Antimicrobial Resistance Surveillance in Europe 2022–2020 Data; World Health Organization, Regional Office for Europe: Copenhagen, Denmark, 2022. [Google Scholar]
- Tuchilus, C.; Mereuta, A.I.; Badescu, A.; Buiuc, D. High Prevalence of CTX-M Group of Extended Spectrum Beta-Lactamases in Enterobacteriaceae strains isolated from the North-East of Romania. Rev. Romana Med. Lab. 2011, 19, 251–257. [Google Scholar]
- Maciuca, I.E.; Williams, N.J.; Tuchilus, C.; Dorneanu, O.; Guguianu, E.; Carp-Carare, C.; Rimbu, C.; Timofte, D. High Prevalence of Escherichia coli-Producing CTX-M-15 Extended-Spectrum Beta-Lactamases in Poultry and Human Clinical Isolates in Romania. Microb. Drug Resist. 2015, 21, 651–662. [Google Scholar] [CrossRef]
- Petca, R.C.; Mareș, C.; Petca, A.; Negoiță, S.; Popescu, R.I.; Boț, M.; Barabás, E.; Chibelean, C.B. Spectrum and Antibiotic Resistance of Uropathogens in Romanian Females. Antibiotics 2020, 9, 472. [Google Scholar] [CrossRef]
- Arbune, M.; Gurau, G.; Niculet, E.; Iancu, A.V.; Lupasteanu, G.; Fotea, S.; Vasile, M.C.; Tatu, A.L. Prevalence of Antibiotic Resistance of ESKAPE Pathogens Over Five Years in an Infectious Diseases Hospital from South-East of Romania. Infect. Drug Resist. 2021, 14, 2369–2378. [Google Scholar] [CrossRef]
- Hritcu, O.M.; Schmidt, V.M.; Salem, S.E.; Maciuca, I.E.; Moraru, R.F.; Lipovan, I.; Mareş, M.; Solcan, G.; Timofte, D. Geographical Variations in Virulence Factors and Antimicrobial Resistance amongst Staphylococci Isolated from Dogs from the United Kingdom and Romania. Front. Vet. Sci. 2020, 7, 414. [Google Scholar] [CrossRef]
- János, D.; Viorel, H.; Ionica, I.; Corina, P.; Tiana, F.; Roxana, D. Carriage of Multidrug Resistance Staphylococci in Shelter Dogs in Timisoara, Romania. Antibiotics 2021, 10, 801. [Google Scholar] [CrossRef]
- Dégi, J.; Moțco, O.-A.; Dégi, D.M.; Suici, T.; Mareș, M.; Imre, K.; Cristina, R.T. Antibiotic Susceptibility Profile of Pseudomonas aeruginosa Canine Isolates from a Multicentric Study in Romania. Antibiotics 2021, 10, 846. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.S.; Song, W.; Park, H.M.; Oh, J.Y.; Chae, J.C.; Shin, S.; Jeong, S.H. Clonal Spread of Extended-Spectrum Cephalosporin-Resistant Enterobacteriaceae between Companion Animals and Humans in South Korea. Front. Microbiol. 2019, 10, 1371. [Google Scholar] [CrossRef] [PubMed]
- Haenni, M.; Saras, E.; Métayer, V.; Médaille, C.; Madec, J.Y. High prevalence of blaCTX-M-1/IncI1/ST3 and blaCMY-2/IncI1/ST2 plasmids in healthy urban dogs in France. Antimicrob. Agents Chemother. 2014, 58, 5358–5362. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Salinas, F.; Díaz-Jiménez, D.; García-Meniño, I.; Lumbreras, P.; López-Beceiro, A.M.; Fidalgo, L.E.; Rodicio, M.R.; Mora, A.; Fernández, J. High Prevalence and Diversity of Cephalosporin-Resistant Enterobacteriaceae Including Extraintestinal Pathogenic E. coli CC648 Lineage in Rural and Urban Dogs in Northwest Spain. Antibiotics 2020, 9, 468. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Betts, J.; La Ragione, R.; van Essen-Zandbergen, A.; Ceccarelli, D.; Petinaki, E.; Koutinas, C.K.; Mevius, D.J. Occurrence and characterization of extended-spectrum cephalosporin-resistant Enterobacteriaceae in healthy household dogs in Greece. J. Med. Microbiol. 2018, 67, 931–935. [Google Scholar] [CrossRef]
- Umeda, K.; Hase, A.; Matsuo, M.; Horimoto, T.; Ogasawara, J. Prevalence and genetic characterization of cephalosporin-resistant Enterobacteriaceae among dogs and cats in an animal shelter. J. Med. Microbiol. 2019, 68, 339–345. [Google Scholar] [CrossRef]
- Verma, A.; Carney, K.; Taylor, M.; Amsler, K.; Morgan, J.; Gruszynski, K.; Erol, E.; Carter, C.; Locke, S.; Callipare, A.; et al. Occurrence of potentially zoonotic and cephalosporin resistant enteric bacteria among shelter dogs in the Central and South-Central Appalachia. BMC Vet. Res. 2021, 17, 313. [Google Scholar] [CrossRef]
- Johansson, V.; Nykäsenoja, S.; Myllyniemi, A.-L.; Rossow, H.; Heikinheimo, A. Genomic characterization of ESBL/AmpC-producing and high-risk clonal lineages of Escherichia coli and Klebsiella pneumoniae in imported dogs with shelter and stray background. J. Glob. Antimicrob. Resist. 2022, 30, 183–190. [Google Scholar] [CrossRef]
- Zogg, A.L.; Simmen, S.; Zurfluh, K.; Stephan, R.; Schmitt, S.N.; Nüesch-Inderbinen, M. High Prevalence of Extended-Spectrum β-Lactamase Producing Enterobacteriaceae among Clinical Isolates from Cats and Dogs Admitted to a Veterinary Hospital in Switzerland. Front. Vet. Sci. 2018, 5, 62. [Google Scholar] [CrossRef]
- Wedley, A.L.; Dawson, S.; Maddox, T.W.; Coyne, K.P.; Pinchbeck, G.L.; Clegg, P.; Nuttall, T.; Kirchner, M.; Williams, N.J. Carriage of antimicrobial resistant Escherichia coli in dogs: Prevalence, associated risk factors and molecular characteristics. Vet. Microbiol. 2017, 199, 23–30. [Google Scholar] [CrossRef]
- Dahmen, S.; Haenni, M.; Châtre, P.; Madec, J.-Y. Characterization of blaCTX-M IncFII plasmids and clones of Escherichia coli from pets in France. J. Antimicrob. Chemother. 2013, 68, 2797–2801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madec, J.Y.; Poirel, L.; Saras, E.; Gourguechon, A.; Girlich, D.; Nordmann, P.; Haenni, M. Non-ST131 Escherichia coli from cattle harbouring human-like bla(CTX-M-15)-carrying plasmids. J. Antimicrob. Chemother. 2012, 67, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Weigel, L.M.; Rasheed, J.K.; McGowan, J.E., Jr.; Raney, P.; Tenover, F.C. Mechanisms of decreased susceptibility to cefpodoxime in Escherichia coli. Antimicrob. Agents Chemother. 2002, 46, 3829–3836. [Google Scholar] [CrossRef] [PubMed]
- van den Bunt, G.; Fluit, A.C.; Spaninks, M.P.; Timmerman, A.J.; Geurts, Y.; Kant, A.; Scharringa, J.; Mevius, D.; Wagenaar, J.A.; Bonten, M.J.M.; et al. Faecal carriage, risk factors, acquisition and persistence of ESBL-producing Enterobacteriaceae in dogs and cats and co-carriage with humans belonging to the same household. J. Antimicrob. Chemother. 2020, 75, 342–350. [Google Scholar] [CrossRef]
- Toombs-Ruane, L.J.; Benschop, J.; French, N.P.; Biggs, P.J.; Midwinter, A.C.; Marshall, J.C.; Chan, M.; Drinković, D.; Fayaz, A.; Baker, M.G.; et al. Carriage of Extended-Spectrum-Beta-Lactamase- and AmpC Beta-Lactamase-Producing Escherichia coli Strains from Humans and Pets in the Same Households. Appl. Environ. Microbiol. 2020, 86, e01613-20. [Google Scholar] [CrossRef]
- AlMasoud, N.; Muhamadali, H.; Chisanga, M.; AlRabiah, H.; Lima, C.A.; Goodacre, R. Discrimination of bacteria using whole organism fingerprinting: The utility of modern physicochemical techniques for bacterial typing. Analyst 2021, 146, 770–788. [Google Scholar] [CrossRef]
- Martak, D.; Valot, B.; Sauget, M.; Cholley, P.; Thouverez, M.; Bertrand, X.; Hocquet, D. Fourier-Transform InfraRed Spectroscopy Can Quickly Type Gram-Negative Bacilli Responsible for Hospital Outbreaks. Front. Microbiol. 2019, 10, 1440. [Google Scholar] [CrossRef]
- Rakovitsky, N.; Frenk, S.; Kon, H.; Schwartz, D.; Temkin, E.; Solter, E.; Paikin, S.; Cohen, R.; Schwaber, M.J.; Carmeli, Y.; et al. Fourier Transform Infrared Spectroscopy Is a New Option for Outbreak Investigation: A Retrospective Analysis of an Extended-Spectrum-Beta-Lactamase-Producing Klebsiella pneumoniae Outbreak in a Neonatal Intensive Care Unit. J. Clin. Microbiol. 2020, 58, e00098-20. [Google Scholar] [CrossRef]
- Sousa, C.; Novais, Â.; Magalhães, A.; Lopes, J.; Peixe, L. Diverse high-risk B2 and D Escherichia coli clones depicted by Fourier Transform Infrared Spectroscopy. Sci. Rep. 2013, 3, 3278. [Google Scholar] [CrossRef]
- Dinkelacker, A.G.; Vogt, S.; Oberhettinger, P.; Mauder, N.; Rau, J.; Kostrzewa, M.; Rossen, J.W.A.; Autenrieth, I.B.; Peter, S.; Liese, J. Typing and Species Identification of Clinical Klebsiella Isolates by Fourier Transform Infrared Spectroscopy and Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2018, 56, e00843-18. [Google Scholar] [CrossRef]
- Vogt, S.; Löffler, K.; Dinkelacker, A.G.; Bader, B.; Autenrieth, I.B.; Peter, S.; Liese, J. Fourier-Transform Infrared (FTIR) Spectroscopy for Typing of Clinical Enterobacter cloacae Complex Isolates. Front. Microbiol. 2019, 10, 2582. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhou, H.; Lu, J.; Sun, Q.; Liu, C.; Zeng, Y.; Zhang, R. Evaluation of the IR Biotyper for Klebsiella pneumoniae typing and its potentials in hospital hygiene management. Microb. Biotechnol. 2021, 14, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.C.; Barbosa, A.V.; Arais, L.R.; Ribeiro, P.F.; Carneiro, V.C.; Cerqueira, A.M.F. Resistance patterns, ESBL genes, and genetic relatedness of Escherichia coli from dogs and owners. Braz. J. Microbiol. 2016, 47, 150–158. [Google Scholar] [CrossRef]
- Elankumaran, P.; Browning, G.F.; Marenda, M.S.; Reid, C.J.; Djordjevic, S.P. Close genetic linkage between human and companion animal extraintestinal pathogenic Escherichia coli ST127. Curr. Res. Microb. Sci. 2022, 3, 100106. [Google Scholar] [CrossRef]
- Johnson, J.R.; Owens, K.; Gajewski, A.; Clabots, C. Escherichia coli Colonization Patterns among Human Household Members and Pets, with Attention to Acute Urinary Tract Infection. J. Infect. Dis. 2008, 197, 218–224. [Google Scholar] [CrossRef]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef]
- Matamoros, S.; van Hattem, J.M.; Arcilla, M.S.; Willemse, N.; Melles, D.C.; Penders, J.; Vinh, T.N.; Thi Hoa, N.; Bootsma, M.C.J.; van Genderen, P.J.; et al. Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci. Rep. 2017, 7, 15364. [Google Scholar] [CrossRef]
- Day, M.J.; Rodríguez, I.; van Essen-Zandbergen, A.; Dierikx, C.; Kadlec, K.; Schink, A.-K.; Wu, G.; Chattaway, M.A.; DoNascimento, V.; Wain, J.; et al. Diversity of STs, plasmids and ESBL genes among Escherichia coli from humans, animals and food in Germany, the Netherlands and the UK. J. Antimicrob. Chemother. 2016, 71, 1178–1182. [Google Scholar] [CrossRef]
- Fooks, A.R.; Johnson, N. Jet set pets: Examining the zoonosis risk in animal import and travel across the European Union. Vet. Med. 2015, 6, 17–25. [Google Scholar] [CrossRef]
- Huang, T.D.; Bogaerts, P.; Berhin, C.; Guisset, A.; Glupczynski, Y. Evaluation of Brilliance ESBL agar, a novel chromogenic medium for detection of extended-spectrum-beta- lactamase-producing Enterobacteriaceae. J. Clin. Microbiol. 2010, 48, 2091–2096. [Google Scholar] [CrossRef]
- McDaniels, A.E.; Rice, E.W.; Reyes, A.L.; Johnson, C.H.; Haugland, R.A.; Stelma, G.N. Confirmational identification of Escherichia coli, a comparison of genotypic and phenotypic assays for glutamate decarboxylase and beta-D-glucuronidase. Appl. Environ. Microbiol. 1996, 62, 3350–3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasi, E.M.; Matthews, B.; Gundogdu, A.; Vollmerhausen, T.L.; Ramos, N.L.; Stratton, H.; Ahmed, W.; Katouli, M. Prevalence and persistence of Escherichia coli strains with uropathogenic virulence characteristics in sewage treatment plants. Appl. Environ. Microbiol. 2010, 76, 5882–5886. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, C.M.; Rhoden, D.L.; Miller, J.M. Reevaluation of the API 20E identification system versus conventional biochemicals for identification of members of the family Enterobacteriaceae: A new look at an old product. J. Clin. Microbiol. 1992, 30, 123–125. [Google Scholar] [CrossRef] [PubMed]
- CLSI Approved Standard M100-S15; Performance Standards for Antimicrobial Susceptibility Testing. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018.
- Dallenne, C.; Da Costa, A.; Decre, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef]
- Park, C.H.; Robicsek, A.; Jacoby, G.A.; Sahm, D.; Hooper, D.C. Prevalence in the United States of aac(6′)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob. Agents Chemother. 2006, 50, 3953–3955. [Google Scholar] [CrossRef]
- Perez-Perez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC beta-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef]
- Robicsek, A.; Strahilevitz, J.; Sahm, D.F.; Jacoby, G.A.; Hooper, D.C. qnr Prevalence in ceftazidime-resistant Enterobacteriaceae isolates from the United States. Antimicrob. Agents Chemother. 2006, 50, 2872–2874. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef]
- Clermont, O.; Dhanji, H.; Upton, M.; Gibreel, T.; Fox, A.; Boyd, D.; Mulvey, M.R.; Nordmann, P.; Ruppe, E.; Sarthou, J.L.; et al. Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 2009, 64, 274–277. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Host | Source | Bacterial Species | No. and Isolate ID (n = 73) | Cluster by FTIR | PG Typing | MLST by WGS | Beta-Lactamase Genes by PCR | Beta-Lactamase Genes by WGS | PMQR Genes | Additional Resistance Determinants by WGS | Inc Type by PBRT |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Human | Dog owner | E. coli | 1 (MV10P) | 18 | A | - | CTX-M-1 group® | - | - | - | FIB; I1 |
| E. coli | 1 (MV16P) | 94 | A | - | CTX-M-1 group® | - | qnrS | - | FIB | ||
| E. coli | 1 (MV22P) | 57 | A | - | blaCTX-M-15, blaTEM, blaOXA | - | aac-(6′)-lb-cr | - | FIB/FIA; F | ||
| E. coli | 1(MV21P) | 17 | B2 | - | blaCTX-M-14 | - | - | - | FIB/FIA; F | ||
| E. coli | 1 (MV18P purple) | 95 | D | - | blaCTX-M-14, blaTEM | - | - | - | FIB; F/I1 | ||
| E. coli | 1 (MV11P) | 90 | B1 | UN | blaSHV, blaTEM | blaTEM-1, blaSHV-134, ampC1, ampH | - | sul3, tet(A), APH(4)-Ia, AAC(3)-IV, aadA, aadA2, APH(3′)-Iia | N | ||
| E. coli | 1 (MV11P-Inc) | 90 | D | - | blaSHV, blaTEM | - | qnrB | - | FIB/FIA/W/K; N/F | ||
| E. coli | 1 (MV23P) | 97 | D | - | blaTEM-1 | - | - | - | P1 | ||
| E. coli | 1 (MV17P) | 8 | B2 | - | blaOXA | - | qnrS; aac-(6′)-lb-cr | - | FIB/FIA; F | ||
| E. coli | 1 (MV8P purple) | 89 | B2 | ST1057 | ampC | ampH | ND | ||||
| E. coli | 1 (MV15P) | 70 | B1 | ST155 | ampC | ampC1, ampH | AAC(2′)-IIa | ||||
| Kennel staff | E. coli | 1 (MV5P) | 10 | A | - | blaCTX-M-1, blaTEM | - | - | - | Y; I1 | |
| E. coli | 1 (MV14P) | 88 | B2 | - | blaCTX-M-1 | - | qnrS | - | HI2 | ||
| E. coli | 1 (MV19P-Inc) | ND | A | - | blaCTX-M-3 | - | - | - | FIB; F | ||
| E. coli | 1 (MV20P-pink) | ND | B2 | - | blaCTX-M-15, blaTEM | - | - | FIB; F/I1 | |||
| E. coli | 1 (MV24P) | ND | B2 | - | CTX-M-1 group, blaTEM | - | - | - | FIB/FIA; L/M | ||
| E. coli | 1 (MV19P) | ND | D | - | blaCTX-M-14, blaTEM | - | - | - | FIB; F/I1 | ||
| E. coli | 1 (MV3P) | 79 | A | - | blaSHV, blaTEM | - | - | - | F/I1 | ||
| E. coli | 1 (MV12P) | 11 | D | - | blaTEM-1 | - | - | - | FIB; F | ||
| E. coli | 1 (MV26P) | ND | B2 | - | blaTEM-55, blaOXA-1 | - | - | - | FIB/FIA; F | ||
| E. coli | 1 (MV1P) | 88 | B2 | - | - | - | qnrS | - | HI2 | ||
| Klebsiella | 1 (MV4P-Kleb) | ND | ND | - | blaSHV | - | - | - | N | ||
| Dog | Households | E. coli | 1 (MV18) | ND | B2 | - | blaCTX-M-1 | - | qnrS | - | HI2 |
| E. coli | 1 (MV21) | 88 | D | - | blaCTX-M-1 | - | qnrS | - | HI2 | ||
| E. coli | 3 (MV19; MV22; MV40) | ND; ND; 88 | B2 | ST127 (MV40) | CTX-M-1 group | blaCTX-M-1 | qnrS | qnrS1, ampH, sul1, tet(A), tet(B), APH(3′)-Ia, APH(6)-Id, APH(3″)-Ib | HI2 | ||
| E. coli | 1 (MV73) | 36 | B2 | - | CTX-M-1 group®, blaTEM | - | qnrS | - | P1; FIB; F | ||
| E. coli | 1 (MV67) | 72 | C | ST88 | blaCTX-M-3 | blaCTX-M-3, ampC1, ampH | - | ND | FIB, N | ||
| E. coli | 1 (MV46) | 24 | B2 | - | blaCTX-M-15 | - | - | - | FIB; F | ||
| E. coli | 1 (MV67-Inch) | 72 | A | - | blaCTX-M-15 | - | - | - | FIB; N | ||
| E. coli | 1 (MV47) | ND | D | - | blaCTX-M-15, blaTEM, blaSHV | - | - | - | F2A; L/M | ||
| E. coli | 1 (MV39) | ND | B1 | - | blaCTX-M-14 | - | - | - | FIB; I1 | ||
| E. coli | 3 (MV20; MV66; MV16) | ND; ND; 95 | D | ST1011 | blaCTX-M-14; blaTEM | blaCTX-M-14, ampC1, TEM-1 | - | sul2, AAC(3)-Iid | FIB; I1 | ||
| E. coli | 1 (MV43) | ND | B1 | - | blaCTX-M-9 | - | - | - | FIB/K; I1 | ||
| E. coli | 1 (MV17) | 95 | D | - | blaCTX-M-9; blaTEM | - | - | - | FIB; I1 | ||
| E. coli | 1 (MV37) | 61 | B1 | - | blaTEM; blaCIT-M | - | - | A/C | |||
| E. coli | 1 (MV29) | ND | B1 | - | blaTEM-1 | - | - | - | FIB/Y/FIA | ||
| E. coli | 1 (MV30) | ND | A | - | blaTEM-1 | - | - | - | FIB; I1 | ||
| E. coli | 1 (MV33) | ND | A | - | blaTEM-1 | - | - | - | FIB; F | ||
| E. coli | 1 (MV62) | ND | B1 | - | blaTEM | - | - | - | FIB | ||
| E. coli | 1 (MV63) | 3 | B2 | - | blaTEM | - | - | - | - | ||
| E. coli | 1 (MV38) | 32 | B1 | - | blaSHV, blaTEM | - | - | - | FIB; I1 | ||
| E. coli | 1 (MV31) | ND | B1 | - | blaSHV, blaTEM, blaOXA | - | - | - | FIB; N/F | ||
| E. coli | 1 (MV71) | ND | A | - | blaTEM-1, blaSHV-52 | - | - | - | FIB; F/I1 | ||
| E. coli | 1 (MV69) | ND | B1 | - | - | - | aac-(6′)-lb-cr | - | FIB; F | ||
| E. coli | 1 (MV28) | ND | B1 | ST155 | ampC | ampC1, ampH | AAC(2′)-IIa | ||||
| Klebsiella | 1 (MV34) | ND | NI | - | blaSHV, blaTEM | - | qnrA, aac-(6′)-lb-cr | - | HI2 | ||
| Klebsiella | 1 (MV71 purple) | ND | NI | - | blaSHV-52 | - | - | - | - | ||
| Klebsiella | 1 (MV68) | ND | NI | - | blaSHV-2 | - | - | - | - | ||
| Kennels | E. coli | 2 (MV53; MV54) | 80; 97 | A | ST5420 | CTX-M-1 group® | blaCTX-M-3, ampH | - | ND | F/I1 | |
| E. coli | 3 (MV59; MV60; MV61) | 80; 85; ND | B1 | ST5073 | CTX-M-1 group® | blaCTX-M-3, ampC1 | - | ND | F/I1 | ||
| E. coli | 1 (MV15) | 88 | B2 | - | blaCTX-M-1 | - | qnrS | - | HI2 | ||
| E. coli | 1 (MV50) | 94 | A | - | blaCTX-M-1 | - | qnrS | - | FIB | ||
| E. coli | 1 (MV55) | 0 | B1 | - | blaCTX-M-1, blaTEM | - | qnrS | - | I1 | ||
| E. coli | 1 (MV56) | 94 | A | CTX-M-1 group® | qnrS | FIB | |||||
| E. coli | 3 (MV48; MV49; MV51) | 96; 96; 96 | A | CTX-M-1 group® | qnrB | FIB; I1 | |||||
| E. coli | 1 (MV58) | 80 | B1 | blaCTX-M-3 | - | F/I1 | |||||
| E. coli | 1 (MV52) | 97 | A | blaCTX-M-15 | - | F; I1 | |||||
| E. coli | 1 (MV57) | 2 | A | blaCTX-M-15 | qnrS | FIB | |||||
| E. coli | 2 (MV1; MV13) | 77; 77 | B1 | blaCTX-M-14 | - | FIB; I1 | |||||
| E. coli | 1 (MV3) | 79 | A | blaSHV, blaTEM | - | F/I1 | |||||
| E. coli | 1 (MV4) | 48 | A | blaSHV, blaTEM | - | P1; FIB; Y; N/I1 | |||||
| E. coli | 1 (MV9) | 27 | A | blaSHV, blaTEM | - | I1 | |||||
| E. coli | 1 (MV14) | 62 | B2 | blaSHV, blaTEM | aac-(6′)-lb-cr | FIB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cozma, A.P.; Rimbu, C.M.; Zendri, F.; Maciuca, I.E.; Timofte, D. Clonal Dissemination of Extended-Spectrum Cephalosporin-Resistant Enterobacterales between Dogs and Humans in Households and Animal Shelters of Romania. Antibiotics 2022, 11, 1242. https://doi.org/10.3390/antibiotics11091242
Cozma AP, Rimbu CM, Zendri F, Maciuca IE, Timofte D. Clonal Dissemination of Extended-Spectrum Cephalosporin-Resistant Enterobacterales between Dogs and Humans in Households and Animal Shelters of Romania. Antibiotics. 2022; 11(9):1242. https://doi.org/10.3390/antibiotics11091242
Chicago/Turabian StyleCozma, Andreea Paula, Cristina Mihaela Rimbu, Flavia Zendri, Iuliana Elena Maciuca, and Dorina Timofte. 2022. "Clonal Dissemination of Extended-Spectrum Cephalosporin-Resistant Enterobacterales between Dogs and Humans in Households and Animal Shelters of Romania" Antibiotics 11, no. 9: 1242. https://doi.org/10.3390/antibiotics11091242