


Evaluation of Catechin Synergistic and Antibacterial Efficacy on Biofilm Formation and acrA Gene Expression of Uropathogenic E. coli Clinical Isolates

Abstract
:1. Introduction
2. Results
2.1. Evaluation of the Antibacterial Activity
2.2. Biofilm Inhibition
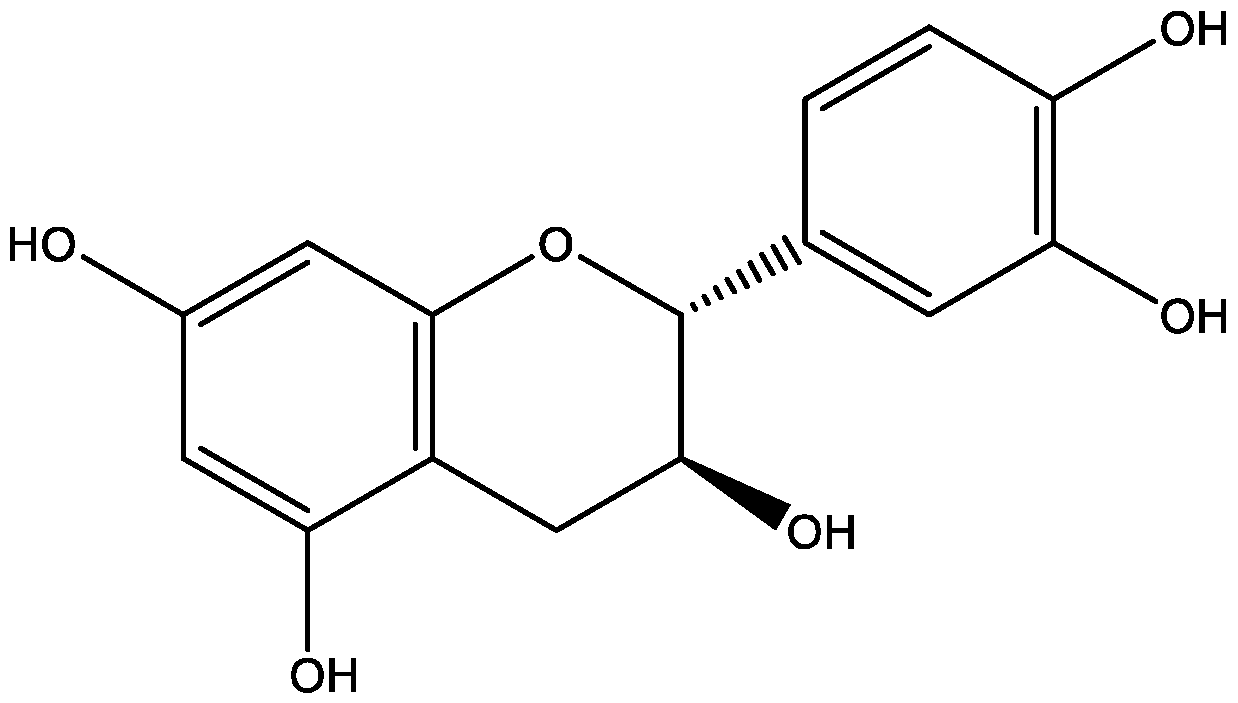
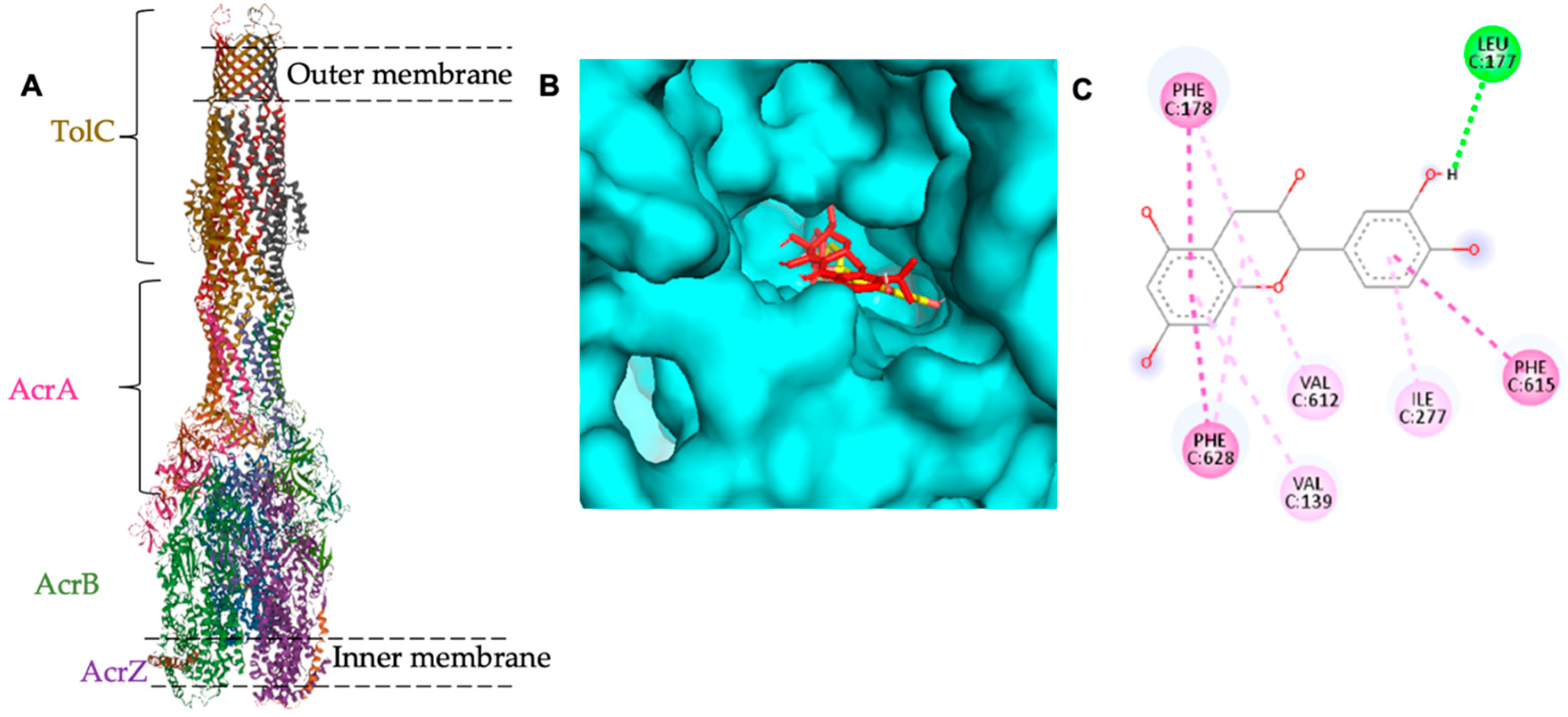
2.3. In Silico Study
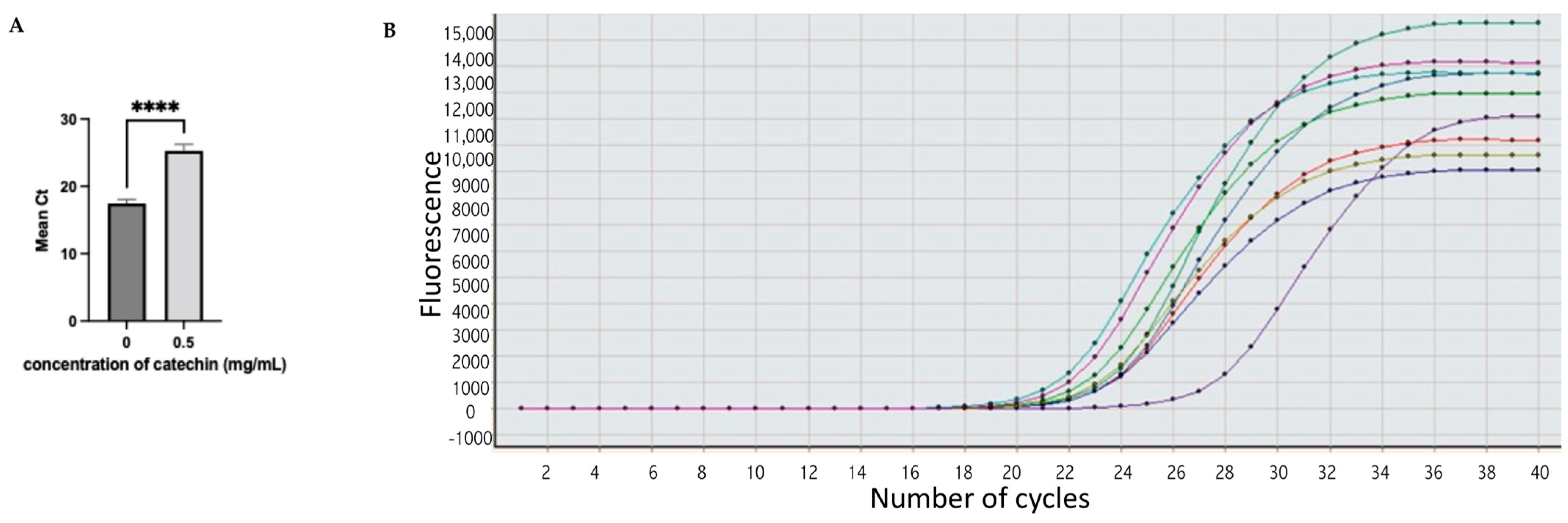
2.4. Gene Expression
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chemicals
4.3. Bacterial Strains
4.4. Evaluation of Antibacterial Activity
4.4.1. Disc Diffusion Assay
4.4.2. Minimum Inhibitory Concentration (MIC)
4.4.3. Minimum Bactericidal Concentration (MBC)
4.4.4. Synergistic Assay
4.5. Evaluation of Antibiofilm Activity
4.5.1. Biofilm Formation by E. coli
4.5.2. Biofilm Inhibition Assay
4.6. In Silico Study
Molecular Docking
4.7. Gene Expression Using Quantitative Real-Time RT-PCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Braz, V.S.; Melchior, K.; Moreira, C.G. Escherichia coli as a Multifaceted Pathogenic and Versatile Bacterium. Front. Cell. Infect. Microbiol. 2020, 10, 548492. [Google Scholar] [CrossRef]
- Desvaux, M.; Dalmasso, G.; Beyrouthy, R.; Barnich, N.; Delmas, J.; Bonnet, R. Pathogenicity Factors of Genomic Islands in Intestinal and Extraintestinal Escherichia coli. Front. Microbiol. 2020, 11, 2065. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Öztürk, R.; Murt, A. Epidemiology of urological infections: A global burden. World J. Urol. 2020, 38, 2669–2679. [Google Scholar] [CrossRef]
- Stamm, W.E.; Norrby, S.R. Urinary Tract Infections: Disease Panorama and Challenges. J. Infect. Dis. 2001, 183, S1–S4. [Google Scholar] [CrossRef]
- Medina, M.; Castillo-Pino, E. An introduction to the epidemiology and burden of urinary tract infections. Ther. Adv. Urol. 2019, 11, 1756287219832172. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Castillo, F.Y.; Moreno-Flores, A.C.; Avelar-González, F.J.; Márquez-Díaz, F.; Harel, J.; Guerrero-Barrera, A.L. An evaluation of multidrug-resistant Escherichia coli isolates in urinary tract infections from Aguascalientes, Mexico: Cross-sectional study. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Méndez, M. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6, 6–14. [Google Scholar] [CrossRef]
- Bohnert, J.A.; Karamian, B.; Nikaido, H. Optimized Nile Red efflux assay of AcrAB-TolC multidrug efflux system shows competition between substrates. Antimicrob. Agents Chemother. 2010, 54, 3770–3775. [Google Scholar] [CrossRef] [PubMed]
- Majumder, S.; Jung, D.; Ronholm, J.; George, S. Prevalence and mechanisms of antibiotic resistance in Escherichia coli isolated from mastitic dairy cattle in Canada. BMC Microbiol. 2021, 21, 222. [Google Scholar] [CrossRef]
- Chen, C.; Liao, X.; Jiang, H.; Zhu, H.; Yue, L.; Li, S.; Fang, B.; Liu, Y. Characteristics of Escherichia coli biofilm production, genetic typing, drug resistance pattern and gene expression under aminoglycoside pressures. Environ. Toxicol. Pharmacol. 2010, 30, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.A.; Khan, A.H.; Adnan, M.; Ullah, H. Traditional uses of medicinal plants used by Indigenous communities for veterinary practices at Bajaur Agency, Pakistan. J. Ethnobiol. Ethnomed. 2018, 14, 11. [Google Scholar] [CrossRef]
- Ahmad Khan, M.S.; Ahmad, I. Chapter 1—Herbal Medicine: Current Trends and Future Prospects. In New Look to Phytomedicine; Ahmad Khan, M.S., Ahmad, I., Chattopadhyay, D., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 3–13. Available online: https://www.sciencedirect.com/science/article/pii/B978012814619400001X (accessed on 13 March 2022).
- Leenhouts, P.W.; Kalkman, C.; Lam, H.J. Burseraceae. Flora Malesiana—Series 1. Spermatophyta 1955, 5, 209–296. [Google Scholar]
- Mogana, R.; Teng-Jin, K.; Wiart, C. In Vitro Antimicrobial, Antioxidant Activities and Phytochemical Analysis of Canarium patentinervium Miq. from Malaysia. Biotechnol. Res. Int. 2011, 2011, 768673. [Google Scholar] [CrossRef] [PubMed]
- Mogana, R.; Adhikari, A.; Tzar, M.N.; Ramliza, R.; Wiart, C. Antibacterial activities of the extracts, fractions and isolated compounds from Canarium patentinervium Miq. against bacterial clinical isolates. BMC Complement. Med. Ther. 2020, 20, 55. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Sartoratto, A.; Machado, A.L.M.; Delarmelina, C.; Figueira, G.M.; Duarte, M.C.T.; Rehder, V.L.G. Composition and antimicrobial activity of essential oils from aromatic plants used in Brazil. Braz. J. Microbiol. 2004, 35, 275–280. [Google Scholar] [CrossRef]
- Thomas, B.T.; Adeleke, A.J.; Raheem-Ademola, R.R.; Kolawole, R.; Musa, O.S. Efficiency of some disinfectants on bacterial wound pathogens. Life Sci. J. 2012, 9, 752–755. [Google Scholar]
- Mathur, T.; Singhal, S.; Khan, S.; Upadhyay, D.J.; Fatma, T.; Rattan, A. Detection of biofilm formation among the clinical isolates of Staphylococci: An evaluation of three different screening methods. Indian J. Med. Microbiol. 2006, 24, 25–29. [Google Scholar] [CrossRef]
- Rajapaksha, P.; Ojo, I.; Yang, L.; Pandeya, A.; Abeywansha, T.; Wei, Y. Insight into the AcrAB-TolC Complex Assembly Process Learned from Competition Studies. Antibiotics 2021, 10, 830. [Google Scholar] [CrossRef]
- Eicher, T.; Seeger, M.A.; Anselmi, C.; Zhou, W.; Brandstätter, L.; Verrey, F.; Diederichs, K.; Faraldo-Gómez, J.D.; Pos, K.M. Coupling of remote alternating-access transport mechanisms for protons and substrates in the multidrug efflux pump AcrB. eLife 2014, 3, e03145. [Google Scholar] [CrossRef]
- Daury, L.; Orange, F.; Taveau, J.-C.; Verchère, A.; Monlezun, L.; Gounou, C.; Marreddy, R.; Picard, M.; Broutin, I.; Pos, K.M.; et al. Tripartite assembly of RND multidrug efflux pumps. Nat. Commun. 2016, 7, 10731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobylka, J.; Kuth, M.S.; Müller, R.T.; Geertsma, E.R.; Pos, K.M. AcrB: A mean, keen, drug efflux machine. Ann. N. Y. Acad. Sci. 2020, 1459, 38–68. [Google Scholar] [CrossRef]
- Zgurskaya, H.I.; Nikaido, H. AcrA is a highly asymmetric protein capable of spanning the periplasm. J. Mol. Biol. 1999, 285, 409–420. Available online: https://www.sciencedirect.com/science/article/pii/S0022283698923130 (accessed on 10 December 2021). [CrossRef]
- Hobbs, E.C.; Yin, X.; Paul, B.J.; Astarita, J.L.; Storz, G. Conserved small protein associates with the multidrug efflux pump AcrB and differentially affects antibiotic resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 16696–16701. [Google Scholar] [CrossRef]
- Ponchel, F.; Toomes, C.; Bransfield, K.; Leong, F.T.; Douglas, S.H.; Field, S.L.; Bell, S.M.; Combaret, V.; Puisieux, A.; Mighell, A.J.; et al. Real-time PCR based on SYBR-Green I fluorescence: An alternative to the TaqMan assay for a relative quantification of gene rearrangements, gene amplifications and micro gene deletions. BMC Biotechnol. 2003, 3, 18. [Google Scholar] [CrossRef]
- Woese, C.R. Bacterial evolution. Microbiol. Rev. 1987, 51, 221–271. [Google Scholar] [CrossRef]
- Kim, Y.H.; Yang, I.; Bae, Y.-S.; Park, S.-R. Performance evaluation of thermal cyclers for PCR in a rapid cycling condition. BioTechniques 2008, 44, 495–505. [Google Scholar] [CrossRef]
- de Oliveira, H.C.; Pinto Garcia Junior, A.A.; Gromboni, J.G.G.; Farias Filho, R.V.; do Nascimento, C.S.; Arias Wenceslau, A. Influence of heat stress, sex and genetic groups on reference genes stability in muscle tissue of chicken. PLoS ONE 2017, 12, e0176402. [Google Scholar]
- Waldeisen, J.R.; Wang, T.; Mitra, D.; Lee, L.P. A Real-Time PCR Antibiogram for Drug-Resistant Sepsis. PLoS ONE 2011, 6, e28528. [Google Scholar] [CrossRef]
- Murray, B.O.; Flores, C.; Williams, C.; Flusberg, D.A.; Marr, E.E.; Kwiatkowska, K.M.; Charest, J.L.; Isenberg, B.C.; Rohn, J.L. Recurrent Urinary Tract Infection: A Mystery in Search of Better Model Systems. Front. Cell. Infect. Microbiol. 2021, 11. Available online: https://www.frontiersin.org/article/10.3389/fcimb.2021.691210 (accessed on 2 January 2022). [CrossRef]
- Mittal, S.; Sharma, M.; Chaudhary, U. Biofilm and multidrug resistance in uropathogenic Escherichia coli. Pathog. Glob. Health 2015, 109, 26–29. [Google Scholar] [CrossRef]
- Foxman, B. The epidemiology of urinary tract infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef]
- de la Fuente-Núñez, C.; Korolik, V.; Bains, M.; Nguyen, U.; Breidenstein, E.B.M.; Horsman, S.; Lewenza, S.; Burrows, L.; Hancock, R.E.W. Inhibition of bacterial biofilm formation and swarming motility by a small synthetic cationic peptide. Antimicrob. Agents Chemother. 2012, 56, 2696–2704. [Google Scholar] [CrossRef]
- Famuyide, I.M.; Aro, A.O.; Fasina, F.O.; Eloff, J.N.; McGaw, L.J. Antibacterial and antibiofilm activity of acetone leaf extracts of nine under-investigated south African Eugenia and Syzygium (Myrtaceae) species and their selectivity indices. BMC Complement. Altern. Med. 2019, 19, 141. [Google Scholar] [CrossRef]
- de la Fuente-Núñez, C.; Cardoso, M.H.; de Souza Cândido, E.; Franco, O.L.; Hancock, R.E.W. Synthetic antibiofilm peptides. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 1061–1069. Available online: https://www.sciencedirect.com/science/article/pii/S0005273615004137 (accessed on 13 December 2021). [CrossRef]
- Gajdács, M.; Kárpáti, K.; Nagy, Á.L.; Gugolya, M.; Stájer, A.; Burián, K. Association between biofilm-production and antibiotic resistance in Escherichia coli isolates: A laboratory-based case study and a literature review. Acta Microbiol. Immunol. Hung. 2021, 68, 217–226. [Google Scholar] [CrossRef]
- Bryers, J.D. Medical biofilm. Biotechnol. Bioeng. 2008, 100, 1–18. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Adnan, M.; Patel, M.; Deshpande, S.; Alreshidi, M.; Siddiqui, A.J.; Reddy, M.N.; Emira, N.; De Feo, V. Effect of Adiantum philippense Extract on Biofilm Formation, Adhesion with Its Antibacterial Activities Against Foodborne Pathogens, and Characterization of Bioactive Metabolites: An in vitro-in silico Approach. Front. Microbiol. 2020, 11, 823. [Google Scholar] [CrossRef]
- Burseraceae, P.W.; Leyden, H.J.L. Leenhouts Dioecious, Rarely Monoecious Trees the Outer Side by Distinct, Closed Twigs, Petioles with Those in Twigs Mostly Amphivasal Mainly Sclerenchymatic XYLEM, Those in the Petioles and Petiolules Collateral and Consisting of Abundant Imparipinnat. Available online: https://repository.naturalis.nl/pub/532548 (accessed on 10 May 2022).
- Mogana, R.; Bradshaw, T.D.; Jin, K.T.; Wiart, C. In Vitro antitumor Potential of Canarium patentinervium Miq. Acad J. Cancer Res. 2011, 4, 1–4. [Google Scholar]
- Mogana, R.; Teng-Jin, K.; Wiart, C. Anti-Inflammatory, Anticholinesterase, and Antioxidant Potential of Scopoletin Isolated from Canarium patentinervium Miq. (Burseraceae Kunth). Evid.-Based Complementary Altern. Med. 2013, 2013, 734824. [Google Scholar] [CrossRef]
- Ma, Y.; Ding, S.; Fei, Y.; Liu, G.; Jang, H.; Fang, J. Antimicrobial activity of anthocyanins and catechins against foodborne pathogens Escherichia coli and Salmonella. Food Control 2019, 106, 106712. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, J.H.; Lee, C.K.; Oh, C.H.; Song, H.J. The Antimicrobial Activity of (−)-Epigallocatehin-3-Gallate and Green Tea Extracts against Pseudomonas aeruginosa and Escherichia coli Isolated from Skin Wounds. Ann. Dermatol. 2014, 26, 564–569. Available online: https://pubmed.ncbi.nlm.nih.gov/25324647 (accessed on 26 September 2014).
- Chunmei, D.; Jiabo, W.; Weijun, K.; Cheng, P.; Xiaohe, X. Investigation of anti-microbial activity of catechin on Escherichia coli growth by microcalorimetry. Environ. Toxicol. Pharmacol. 2010, 30, 284–288. [Google Scholar] [CrossRef]
- Díaz-Gómez, R.; Toledo-Araya, H.; López-Solís, R.; Obreque-Slier, E. Combined effect of gallic acid and catechin against Escherichia coli. LWT 2014, 59, 896–900. [Google Scholar] [CrossRef]
- Zhao, Y.; Qu, Y.; Tang, J.; Chen, J.; Liu, J. Tea Catechin Inhibits Biofilm Formation of Methicillin-Resistant S. aureus. J. Food Qual. 2021, 2021, 8873091. [Google Scholar] [CrossRef]
- Hengge, R. Targeting Bacterial Biofilms by the Green Tea Polyphenol EGCG. Molecules 2019, 24, 2403. [Google Scholar] [CrossRef]
- Hamilton-Miller, J.T. Anti-cariogenic properties of tea (Camellia sinensis). J. Med. Microbiol. 2001, 50, 299–302. [Google Scholar] [CrossRef]
- Lee, K.-M.; Kim, W.-S.; Lim, J.; Nam, S.; Youn, M.; Nam, S.-W.; Kim, Y.; Kim, S.-H.; Park, W.; Park, S. Antipathogenic Properties of Green Tea Polyphenol Epigallocatechin Gallate at Concentrations below the MIC against Enterohemorrhagic Escherichia coli O157:H7. J. Food Prot. 2009, 72, 325–331. [Google Scholar] [CrossRef]
- Wang, Z.; Fan, G.; Hryc, C.F.; Blaza, J.N.; Serysheva, I.I.; Schmid, M.F.; Chiu, W.; Luisi, B.F.; Du, D. An allosteric transport mechanism for the AcrAB-TolC multidrug efflux pump. eLife 2017, 6, e24905. [Google Scholar] [CrossRef]
- Mogana, R.; Adhikari, A.; Debnath, S.; Hazra, S.; Hazra, B.; Teng-Jin, K.; Wiart, C. The Antiacetylcholinesterase and Antileishmanial Activities of Canarium patentinervium Miq. BioMed Res. Int. 2014, 2014, 903529. [Google Scholar] [CrossRef]
- Kiehlbauch, J.A.; Hannett, G.E.; Salfinger, M.; Archinal, W.; Monserrat, C.; Carlyn, C. Use of the National Committee for Clinical Laboratory Standards Guidelines for Disk Diffusion Susceptibility Testing in New York State Laboratories. J. Clin. Microbiol. 2000, 38, 3341–3348. [Google Scholar] [CrossRef]
- CLSI. 2021 Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI M100-ED29; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; Volume 40, pp. 50–51. [Google Scholar]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2015, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Eloff, J.N. A Sensitive and Quick Microplate Method to Determine the Minimal Inhibitory Concentration of Plant Extracts for Bacteria. Planta Med. 1998, 64, 711–713. [Google Scholar] [CrossRef]
- Parvekar, P.; Palaskar, J.; Metgud, S.; Maria, R.; Dutta, S. The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of silver nanoparticles against Staphylococcus aureus. Biomater. Investig. Dent. 2020, 7, 105–109. [Google Scholar] [CrossRef]
- Ozturk, S.; Ercisli, S. Chemical composition and in vitro antibacterial activity of Seseli libanotis. World J. Microbiol. Biotechnol. 2006, 22, 261–265. [Google Scholar] [CrossRef]
- Levison, M.E. Pharmacodynamics of antimicrobial drugs. Infect. Dis. Clin. 2004, 18, 451–465. Available online: https://www.sciencedirect.com/science/article/pii/S0891552004000753 (accessed on 15 May 2022).
- Cantore, P.L.; Iacobellis, N.S.; De Marco, A.; Capasso, F.; Senatore, F. Antibacterial Activity of Coriandrum sativum L. and Foeniculum vulgare Miller Var. vulgare (Miller) Essential Oils. J. Agric. Food Chem. 2004, 52, 7862–7866. [Google Scholar] [CrossRef]
- Saquib, S.A.; AlQahtani, N.A.; Ahmad, I.; Kader, M.A.; Al Shahrani, S.S.; Asiri, E.A. Evaluation and Comparison of Antibacterial Efficacy of Herbal Extracts in Combination with Antibiotics on Periodontal pathobionts: An in vitro Microbiological Study. Antibiotics 2019, 8, 89. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef] [PubMed]
- Sandasi, M.; Leonard, C.; Viljoen, A. The effect of five common essential oil components on Listeria monocytogenes biofilms. Food Control 2008, 19, 1070–1075. [Google Scholar] [CrossRef]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter Plate Assay for Assessment of Listeria monocytogenes Biofilm Formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Danish, S.; Shakil, S.; Haneef, D. A simple click by click protocol to perform docking: AutoDock 4.2 made easy for non-bioinformaticians. EXCLI J. 2013, 12, 831–857. [Google Scholar]
- Yuan, S.; Chan, H.S.; Hu, Z. Using PyMOL as a platform for computational drug design. WIREs Comput. Mol. Sci. 2017, 7, e1298. [Google Scholar] [CrossRef]
- Kosari, F.; Taheri, M.; Moradi, A.; Alni, R.H.; Alikhani, M.Y. Evaluation of cinnamon extract effects on clbB gene expression and biofilm formation in Escherichia coli strains isolated from colon cancer patients. BMC Cancer 2020, 20, 267. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Antimicrobials | Zone of Inhibition (ZI) (mm) | ||||
|---|---|---|---|---|---|---|
| ZI of Antimicrobials | ZI Breakpoints According to CLSI * | ZI of Catechin | ZI of Catechin in Combination with Antimicrobials | Outcome | ||
| Escherichia coli ATCC 25922 # | Rifampin | 7.42 ± 0.0 | R | 10.47 ± 0.0 | 16 ± 0.0 | Antagonism |
| Tetracycline | - | R | 10.4 ± 0.0 | Additive | ||
| Erythromycin | - | R | 10.0 ± 0.0 | Antagonism | ||
| Clindamycin | - | R | 10.45 ± 0.0 | Additive | ||
| Azithromycin | 10.95 ± 0.0 | R | 22.5 | Synergy | ||
| Vancomycin | - | R | 10.4 ± 0.0 | Additive | ||
| Gentamicin | 31.32 ± 0.0 | S | 35 ± 0.0 | Antagonism | ||
| Escherichia coli ATCC 8739 # | Rifampin | 17.24 ± 0.0 | I | 9 ± 0.0 | 26 ± 0.0 | Additive |
| Tetracycline | - | R | 9.1 ± 0.0 | Additive | ||
| Erythromycin | - | R | 8.8 ± 0.0 | Antagonism | ||
| Clindamycin | - | R | 9.0 ± 0.0 | Additive | ||
| Azithromycin | 15.15 ± 0.0 | S | 25 ± 0.0 | Synergy | ||
| Vancomycin | - | R | 9.5 ± 0.0 | Synergy | ||
| Gentamicin | 13.14 ± 0.0 | I | 22 ± 0.0 | Additive | ||
| Escherichia coli ATCC 43895 # | Rifampin | - | R | 11.8 ± 0.0 | 11.5 ± 0.0 | Antagonism |
| Tetracycline | - | R | 12.0 ± 0.0 | Synergy | ||
| Erythromycin | - | R | 11.5 ± 0.0 | Antagonism | ||
| Clindamycin | - | R | 11.45 ± 0.0 | Antagonism | ||
| Azithromycin | 6.03 ± 0.0 | R | 17 ± 0.0 | Synergy | ||
| Vancomycin | - | R | 11.8 ± 0.0 | Additive | ||
| Gentamycin | 13.6 ± 0.0 | I | 22 | Antagonism | ||
| Escherichia coli (isolate 1) | Rifampin | 2.45 ± 4.2 | R | 12.27 ± 1.3 | 13.89 ± 3.6 | Antagonism |
| Tetracycline | 13.85 ± 2.5 | I | 28.58 ± 1.0 | Synergy | ||
| Erythromycin | 11.97 ± 7.9 | I | 25.04 ± 0.6 | Synergy | ||
| Clindamycin | 7.71 ± 8.7 | R | 22.19 ± 1.3 | Synergy | ||
| Azithromycin | 11.58 ± 3.2 | R | 24.8 ± 0.5 | Synergy | ||
| Vancomycin | 13.72 ± 7.2 | I | 26.43 ± 0.5 | Synergy | ||
| Gentamicin | 10.42 ± 5.6 | R | 23.58 ± 0.4 | Synergy | ||
| Escherichia coli (isolate 2) | Rifampin | - | R | 13.92 ± 3.6 | 14.0 ± 0.0 | Additive |
| Tetracycline | 16.83 ± 1.0 | S | 31.5 ± 10.0 | Synergy | ||
| Erythromycin | 18.97 ± 3.5 | I | 33.1 ± 0.0 | Synergy | ||
| Clindamycin | 17.75 ± 0.4 | I | 31.5 ± 4.5 | Additive | ||
| Azithromycin | 12.60 ± 2.84 | I | 27.2 ± 0.5 | Synergy | ||
| Vancomycin | 17.87 ± 2.2 | S | 31.6 ± 0.9 | Additive | ||
| Gentamicin | 16.95 ± 0.3 | S | 30.7 ± 0.8 | Additive | ||
| Escherichia coli (isolate 3) | Rifampin | - | R | 12.9 ± 2.4 | 9.0 ± 5.2 | Antagonism |
| Tetracycline | 12.35 ± 0.4 | I | 26.12 ± 0.2 | Synergy | ||
| Erythromycin | 16.19 ± 1.0 | I | 31.5 ± 2.1 | Synergy | ||
| Clindamycin | 15.37 ± 3.8 | I | 28.2 ± 3.1 | Additive | ||
| Azithromycin | 12.43 ± 2.2 | I | 26.3 ± 0.3 | Synergy | ||
| Vancomycin | 18.87 ± 0.0 | S | 32.0 ± 0.7 | Synergy | ||
| Gentamicin | 16.24 ± 1.3 | S | 29.0 ± 0.1 | Additive | ||
| Escherichia coli (isolate 4) | Rifampin | 10.87 ± 0.0 | R | 13.09 ± 0.0 | 20.21 ± 1.6 | Antagonism |
| Tetracycline | 15.26 ± 0.5 | S | 30.0 ± 1.6 | Synergy | ||
| Erythromycin | 20.61 ± 4.0 | I | 34.0 ± 0.0 | Synergy | ||
| Clindamycin | 16.42 ± 1.1 | I | 29.5 ± 1.2 | Additive | ||
| Azithromycin | 12.42 ± 0.7 | I | 26.1 ± 0.5 | Synergy | ||
| Vancomycin | 20.03 ± 5.2 | S | 25.5 ± 0.1 | Additive | ||
| Gentamicin | - | R | 13.1 ± 2.4 | Additive | ||
| Escherichia coli (isolate 5) | Rifampin | - | R | 10.68 ± 5.6 | 10.71 ± 1.1 | Additive |
| Tetracycline | 16.82 ± 2.3 | S | 30.23 ± 0.0 | Synergy | ||
| Erythromycin | 16.5 ± 0.0 | I | 28.0 ± 4.4 | Synergy | ||
| Clindamycin | - | R | 12.1 ± 3.3 | Synergy | ||
| Azithromycin | 12.12 ± 1.5 | I | 24.0 ± 0.8 | Synergy | ||
| Vancomycin | 17.71 ± 3.4 | S | 29.0 ± 0.3 | Synergy | ||
| Gentamicin | - | R | 11.0 ± 0.2 | Synergy | ||
| Escherichia coli (isolate 6) | Rifampin | - | R | 9.93 ± 3.6 | 9.0 ± 4.5 | Additive |
| Tetracycline | 12.26 ± 1.0 | I | 25.0 ± 0.0 | Synergy | ||
| Erythromycin | 14.1 ± 0.6 | I | 25.0 ± 0.0 | Synergy | ||
| Clindamycin | - | R | 10.5 ± 0.0 | Synergy | ||
| Azithromycin | 12.31 ± 2.6 | I | 23.2 ± 0.4 | Synergy | ||
| Vancomycin | 16.82 ± 0.5 | I | 27.0 ± 0.5 | Synergy | ||
| Gentamicin | 11.45 ± 0.0 | R | 21.8 ± 0.3 | Synergy | ||
| Escherichia coli (isolate 7) | Rifampin | - | R | 12.75 ± 5.6 | 14.74 ± 8.3 | Synergy |
| Tetracycline | 12.93 ± 0.2 | I | 27.3 ± 0.9 | Synergy | ||
| Erythromycin | 14.59 ± 1.4 | I | 28.1 ± 3.4 | Synergy | ||
| Clindamycin | - | R | 13.22 ± 0.2 | Synergy | ||
| Azithromycin | 12.94 ± 0.6 | I | 26.6 ± 0.9 | Synergy | ||
| Vancomycin | 18.21 ± 1.5 | S | 30.89 ± 0.6 | Additive | ||
| Gentamicin | 11.81 ± 0.0 | R | 25.6 ± 0.4 | Synergy | ||
| Escherichia coli (isolate 8) | Rifampin | - | R | 10.08 ± 4.3 | 14.69 ± 0.8 | Synergy |
| Tetracycline | 12.12 ± 11.0 | I | 24.6 ± 5.3 | Synergy | ||
| Erythromycin | 17.73 ± 8.76 | I | 28.4 ± 0.0 | Synergy | ||
| Clindamycin | - | R | 11.54 ± 1.3 | Synergy | ||
| Azithromycin | 12.04 ± 5.6 | I | 23.1 ± 0.2 | Synergy | ||
| Vancomycin | - | R | 10.0 ± 3.4 | Additive | ||
| Gentamicin | 11.02 ± 7.0 | R | 22.0 ± 1.1 | Synergy | ||
| Escherichia coli (isolate 9) | Rifampin | 9.46 ± 1.1 | R | 13.58 ± 2.3 | 23.1 ± 2.5 | Additive |
| Tetracycline | 13.46 ± 2.3 | I | 27.89 ± 6.8 | Synergy | ||
| Erythromycin | 19.09 ± 0.6 | I | 33.1 ± 0.5 | Synergy | ||
| Clindamycin | - | R | 14.0 ± 0.0 | Synergy | ||
| Azithromycin | 12.82 ± 0.0 | I | 27.0 ± 0.4 | Synergy | ||
| Vancomycin | - | R | 13.5 ± 0.6 | Additive | ||
| Gentamicin | 11.1 ± 0.0 | R | 26.4 ± 0.8 | Synergy | ||
| Escherichia coli (isolate 10) | Rifampin | - | R | 12.9 ± 1.7 | 12.6 ± 1.3 | Additive |
| Tetracycline | 13.4 ± 0.5 | I | 27.0 ± 1.1 | Synergy | ||
| Erythromycin | 14.34 ± 4.1 | I | 28.5 ± 8.85 | Synergy | ||
| Clindamycin | 15.38 ± 3.2 | I | 30.0 ± 5.5 | Synergy | ||
| Azithromycin | 12.55 ± 8.5 | I | 26.4 ± 0.5 | Synergy | ||
| Vancomycin | - | R | 12.82 ± 0.9 | Additive | ||
| Gentamicin | 13.52 ± 6.1 | I | 27.2 ± 0.4 | Synergy | ||
| Escherichia coli (isolate 11) | Rifampin | - | R | 10.38 ± 0.6 | 10.4 ± 0.5 | Additive |
| Tetracycline | 12.28 ± 0.0 | I | 24.5 ± 0.7 | Synergy | ||
| Erythromycin | - | R | 10.5 ± 1.4 | Additive | ||
| Clindamycin | 16.69 ± 1.1 | I | 28.1 ± 1.1 | Synergy | ||
| Azithromycin | 12.21 ± 0.9 | I | 22.5 ± 0.5 | Additive | ||
| Vancomycin | 16.59 ± 3.1 | I | 29.0 ± 0.7 | Synergy | ||
| Gentamicin | 13.76 ± 2.5 | I | 25.4 ± 1.1 | Synergy | ||
| Escherichia coli (isolate 12) | Rifampin | 8.76 ± 5.3 | R | 13.17 ± 1.5 | 22.5 ± 6.4 | Synergy |
| Tetracycline | 18.34 ± 7.1 | I | 31.9 ± 0.5 | Synergy | ||
| Erythromycin | - | R | 13.4 ± 11.0 | Additive | ||
| Clindamycin | 15.71 ± 2.6 | I | 30.5 ± 0.0 | Synergy | ||
| Azithromycin | 12.77 ± 1.5 | I | 26.4 ± 0.2 | Synergy | ||
| Vancomycin | 15.24 ± 1.1 | I | 28.5 ± 0.4 | Additive | ||
| Gentamicin | 13.73 ± 0.6 | I | 27.5 ± 0.9 | Synergy | ||
| Escherichia coli (isolate 13) | Rifampin | - | R | 13.18 ± 8.7 | 13.0 ± 0.0 | Additive |
| Tetracycline | 9.82 ± 4.3 | R | 25.4 ± 1.4 | Synergy | ||
| Erythromycin | - | R | 14.21 ± 0.0 | Synergy | ||
| Clindamycin | 18.27 ± 5.2 | I | 32.0 ± 0.5 | Synergy | ||
| Azithromycin | 12.67 ± 9.7 | I | 27.1 ± 0.0 | Synergy | ||
| Vancomycin | 16.37 ± 10.0 | I | 29.45 ± 0.5 | Additive | ||
| Gentamicin | 13.14 ± 8.7 | I | 27.2 ± 0.3 | Synergy | ||
| Escherichia coli (isolate 14) | Rifampin | 7.67 ± 2.6 | R | 12.4 ± 1.3 | 20.0 ± 3.8 | Additive |
| Tetracycline | 10.22 ± 11.0 | R | 23.4 ± 1.5 | Synergy | ||
| Erythromycin | - | R | 12.42 ± 0.5 | Additive | ||
| Clindamycin | - | R | 13.1 ± 0.6 | Synergy | ||
| Azithromycin | 11.28 ± 5.4 | I | 23.5 ± 2.7 | Additive | ||
| Vancomycin | 16.24 ± 8.6 | I | 29.0 ± 0.4 | Synergy | ||
| Gentamicin | 11.81 ± 5.9 | I | 25.1 ± 0.2 | Synergy | ||
| Escherichia coli (isolate 15) | Rifampin | - | R | 12.62 ± 6.6 | 12.5 ± 7.8 | Additive |
| Tetracycline | 15.89 ± 7.4 | S | 29.1 ± 0.0 | Synergy | ||
| Erythromycin | 9.42 ± 11.0 | R | 24.01 ± 11.6 | Synergy | ||
| Clindamycin | - | R | 14.0 ± 4.3 | Synergy | ||
| Azithromycin | - | R | 12.5 ± 0.9 | Additive | ||
| Vancomycin | 15.37 ± 8.3 | I | 28.0 ± 0.9 | Additive | ||
| Gentamicin | - | R | 13.2 ± 0.6 | Synergy | ||
| Bacteria | Effect | Rifampin | Tetracycline | Erythromycin | Clindamycin | Azithromycin | Vancomycin | Gentamycin | Catechin |
|---|---|---|---|---|---|---|---|---|---|
| Escherichia coli ATCC 25922 # | MBC | 8 | 32 | 32 | 8 | 8 | 64 | 4 | 2 |
| MIC | 4 | 16 | 8 | 4 | 4 | 16 | 2 | 1 | |
| MBC/MIC | 2(+) | 2(+) | 4(+) | 2(+) | 2(+) | 4(+) | 2(+) | 2(+) | |
| Escherichia coli ATCC 8739 # | MBC | 4 | 32 | 32 | 16 | 1 | 32 | 4 | 4 |
| MIC | 2 | 16 | 16 | 8 | 0.5 | 16 | 2 | 2 | |
| MBC/MIC | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | |
| Escherichia coli ATCC 43895 # | MBC | 8 | 32 | 16 | 8 | 16 | 16 | 4 | 2 |
| MIC | 4 | 16 | 8 | 4 | 8 | 8 | 2 | 1 | |
| MBC/MIC | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | 2(+) | |
| Escherichia coli (isolate 1) | MBC | 32 | 32 | 32 | 32 | 2 | 32 | 8 | 4 |
| MIC | 16 | 8 | 4 | 4 | 0.5 | 8 | 4 | 1 | |
| MBC/MIC | 2(+) | 4(+) | 8(−) | 8(−) | 4(+) | 4(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 2) | MBC | 16 | 32 | 16 | 32 | 2 | 32 | 8 | 4 |
| MIC | 8 | 4 | 4 | 4 | 1 | 8 | 4 | 1 | |
| MBC/MIC | 2(+) | 8(−) | 4(+) | 8(−) | 2(+) | 4(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 3) | MBC | 16 | 32 | 32 | 32 | 1 | 16 | 8 | 4 |
| MIC | 8 | 8 | 4 | 4 | 0.5 | 2 | 4 | 1 | |
| MBC/MIC | 2(+) | 4(+) | 8(−) | 8(−) | 2(+) | 8(−) | 2(+) | 4(+) | |
| Escherichia coli (isolate 4) | MBC | 32 | 64 | 32 | 32 | 1 | 32 | 8 | 4 |
| MIC | 16 | 4 | 8 | 8 | 0.5 | 8 | 4 | 1 | |
| MBC/MIC | 2(+) | 16(−) | 4(+) | 4(+) | 2(+) | 4(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 5) | MBC | 16 | 64 | 32 | 32 | 1 | 32 | 8 | 4 |
| MIC | 8 | 4 | 8 | 2 | 0.5 | 8 | 4 | 1 | |
| MBC/MIC | 2(+) | 16(−) | 4(+) | 16(−) | 2(+) | 4(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 6) | MBC | 16 | 32 | 16 | 32 | 2 | 64 | 4 | 4 |
| MIC | 8 | 4 | 4 | 8 | 1 | 16 | 1 | 1 | |
| MBC/MIC | 2(+) | 8(−) | 4(+) | 4(+) | 2(+) | 4(+) | 4(+) | 4(+) | |
| Escherichia coli (isolate 7) | MBC | 32 | 32 | 64 | 32 | 2 | 64 | 16 | 4 |
| MIC | 8 | 8 | 8 | 2 | 1 | 8 | 8 | 1 | |
| MBC/MIC | 4(+) | 4(+) | 8(−) | 16(−) | 2(+) | 8(−) | 2(+) | 4(+) | |
| Escherichia coli (isolate 8) | MBC | 64 | 32 | 32 | 32 | 4 | 32 | 32 | 4 |
| MIC | 8 | 8 | 4 | 4 | 1 | 16 | 16 | 1 | |
| MBC/MIC | 8(−) | 4(+) | 8(−) | 8(−) | 4(+) | 2(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 9) | MBC | 64 | 32 | 32 | 32 | 2 | 32 | 4 | 2 |
| MIC | 16 | 8 | 8 | 4 | 1 | 8 | 2 | 0.5 | |
| MBC/MIC | 4(+) | 4(+) | 4(−) | 8(−) | 2(+) | 4(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 10) | MBC | 64 | 32 | 32 | 32 | 4 | 64 | 4 | 2 |
| MIC | 32 | 8 | 4 | 4 | 1 | 16 | 2 | 0.5 | |
| MBC/MIC | 2(+) | 4(+) | 8(−) | 8(−) | 4(+) | 4(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 11) | MBC | 64 | 32 | 16 | 32 | 2 | 16 | 8 | 4 |
| MIC | 32 | 8 | 4 | 4 | 0.5 | 4 | 2 | 1 | |
| MBC/MIC | 2(+) | 4(+) | 4(+) | 8(−) | 4(+) | 4(+) | 4(+) | 4(+) | |
| Escherichia coli (isolate 12) | MBC | 64 | 32 | 64 | 32 | 4 | 32 | 8 | 4 |
| MIC | 32 | 8 | 8 | 4 | 1 | 8 | 2 | 1 | |
| MBC/MIC | 2(+) | 4(+) | 8(−) | 8(−) | 4(+) | 4(+) | 4(+) | 4(+) | |
| Escherichia coli (isolate 13) | MBC | 64 | 32 | 16 | 16 | 2 | 32 | 8 | 4 |
| MIC | 32 | 4 | 8 | 4 | 0.5 | 8 | 4 | 1 | |
| MBC/MIC | 2(+) | 8(−) | 2(+) | 4(+) | 4(+) | 4(+) | 2(+) | 4(+) | |
| Escherichia coli (isolate 14) | MBC | 16 | 64 | 32 | 32 | 2 | 32 | 8 | 4 |
| MIC | 8 | 32 | 4 | 4 | 1 | 4 | 4 | 1 | |
| MBC/MIC | 2(+) | 2(+) | 8(−) | 8(−) | 2(+) | 8(−) | 2(+) | 4(+) | |
| Escherichia coli (isolate 15) | MBC | 32 | 64 | 32 | 32 | 1 | 32 | 8 | 4 |
| MIC | 16 | 32 | 4 | 4 | 0.5 | 16 | 4 | 1 | |
| MBC/MIC | 2(+) | 2(+) | 8(−) | 8(−) | 2(+) | 2(+) | 2(+) | 4(+) |
| Compound | Molecular Docking Binding Affinity ΔG (Kcal/mol) | Residue Involved in the Binding Site | Bonds Involved in the Binding Site |
|---|---|---|---|
| Minocycline (control) | −8.8 | PHE-178 (chain C), ASN-274 (chain C) | Hydrogen bond |
| VAL-612 (chain C), ALA-279 (chain C) | Pi-Pi stacked | ||
| ILE-277 (chain C), PHE-615 (chain C) | Pi-Sigma | ||
| Catechin | −8.2 | GLY-179 (chain C) | Hydrogen bond |
| LEU-177 (chain C) | Carbon Hydrogen bond | ||
| PHE-178 (chain C), VAL-612 (chain C), ILE-277 (chain C) | Pi-Pi stacked |
| Isolate Number | Mean of Ct of 16S rRNA (Untreated) | Mean of Ct of 16S rRNA (Treated) | Mean of Ct of acrA Gene (Untreated) | Mean of Ct of acrA Gene (Catechin Treated) | ΔCt for acrA Gene | Result |
|---|---|---|---|---|---|---|
| 1 | 17 | 19.34 | 17.8 | 24.8 | 7 | Down |
| 2 | 16.9 | 18.45 | 17.2 | 24.7 | 7.5 | Down |
| 3 | 17.2 | 19.5 | - | - | - | - |
| 4 | 17.2 | 18.45 | - | - | - | |
| 5 | 17.3 | 21.5 | 16.8 | 24.7 | 7.9 | Down |
| 6 | 17.6 | 19.34 | 18.3 | 25 | 6.9 | Down |
| 7 | 18.2 | 19 | - | - | - | - |
| 8 | 17.7 | 21 | - | - | - | - |
| 9 | 18 | 20.41 | 17.8 | 24.8 | 7 | Down |
| 10 | 18.1 | 19.45 | 16.6 | 26.7 | 10.1 | Down |
| 11 | 17.6 | 18 | - | - | - | |
| 12 | 17.7 | 18.2 | 18 | 27.2 | 9.2 | Down |
| 13 | 16.9 | 17.7 | 17.7 | 24.8 | 7.1 | Down |
| 14 | 16.7 | 17.9 | 16.9 | 24.7 | 7.8 | Down |
| 15 | 17.3 | 19.3 | - | - | - | - |
| Bacteria | Source | Resistance Profile |
|---|---|---|
| Escherichia coli ATCC 25922 | Central Health Lab/Iraq | (R) TET, ERY, VAN, RIF, AZM, PIP (S) GEN, MIN, MEM |
| Escherichia coli ATCC 8739 | Central Health Lab/Iraq | (R) TET, ERY, CLI, VAN (S) GEN, AZM, RIF |
| Escherichia coli ATCC 43895 | Central Health Lab/Iraq | (R) RIF, TET, ERY, AZM, PIP, TIC, TOB (S) CFM, FEP, MIN |
| Escherichia coli | Urine from UTI samples | (R) TIC, PIP, GEN, TOB, CIP, SXT (S) TIM, TZP, CAZ, FEP, ATM, IPM, MEM, AMK, MIN |
| Genes | Type | Sequences (5′–3′) | Temperature (C) |
|---|---|---|---|
| 16 sRNA (reference gene) | Forward Reverse | AGAGTTTGATCMTGGCTCAG CTGCTGCSYCCCGTAG | 50 52 |
| acrAgene (target gene) | Forward Reverse | TTGAAATTCAGGAT CTTAGCCCTAACAGGATGTG | 53 57.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jubair, N.; R., M.; Fatima, A.; Mahdi, Y.K.; Abdullah, N.H. Evaluation of Catechin Synergistic and Antibacterial Efficacy on Biofilm Formation and acrA Gene Expression of Uropathogenic E. coli Clinical Isolates. Antibiotics 2022, 11, 1223. https://doi.org/10.3390/antibiotics11091223
Jubair N, R. M, Fatima A, Mahdi YK, Abdullah NH. Evaluation of Catechin Synergistic and Antibacterial Efficacy on Biofilm Formation and acrA Gene Expression of Uropathogenic E. coli Clinical Isolates. Antibiotics. 2022; 11(9):1223. https://doi.org/10.3390/antibiotics11091223
Chicago/Turabian StyleJubair, Najwan, Mogana R., Ayesha Fatima, Yasir K. Mahdi, and Nor Hayati Abdullah. 2022. "Evaluation of Catechin Synergistic and Antibacterial Efficacy on Biofilm Formation and acrA Gene Expression of Uropathogenic E. coli Clinical Isolates" Antibiotics 11, no. 9: 1223. https://doi.org/10.3390/antibiotics11091223