

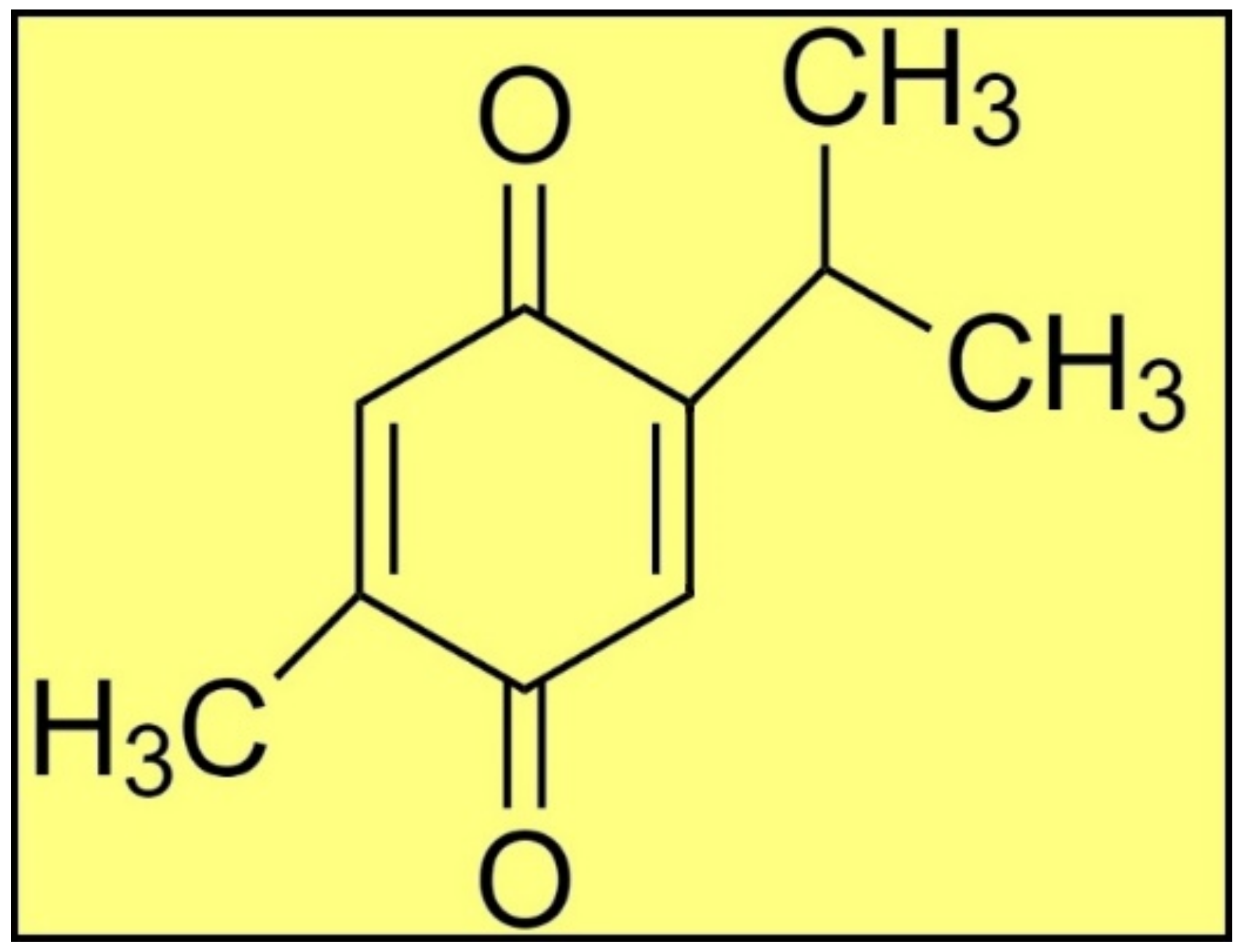
Antiprotozoal Activity of Thymoquinone (2-Isopropyl-5-methyl-1,4-benzoquinone) for the Treatment of Leishmania major-Induced Leishmaniasis: In Silico and In Vitro Studies

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. In Vitro Antileishmanial Activity of TQ
2.1.1. Antipromastigote Activity
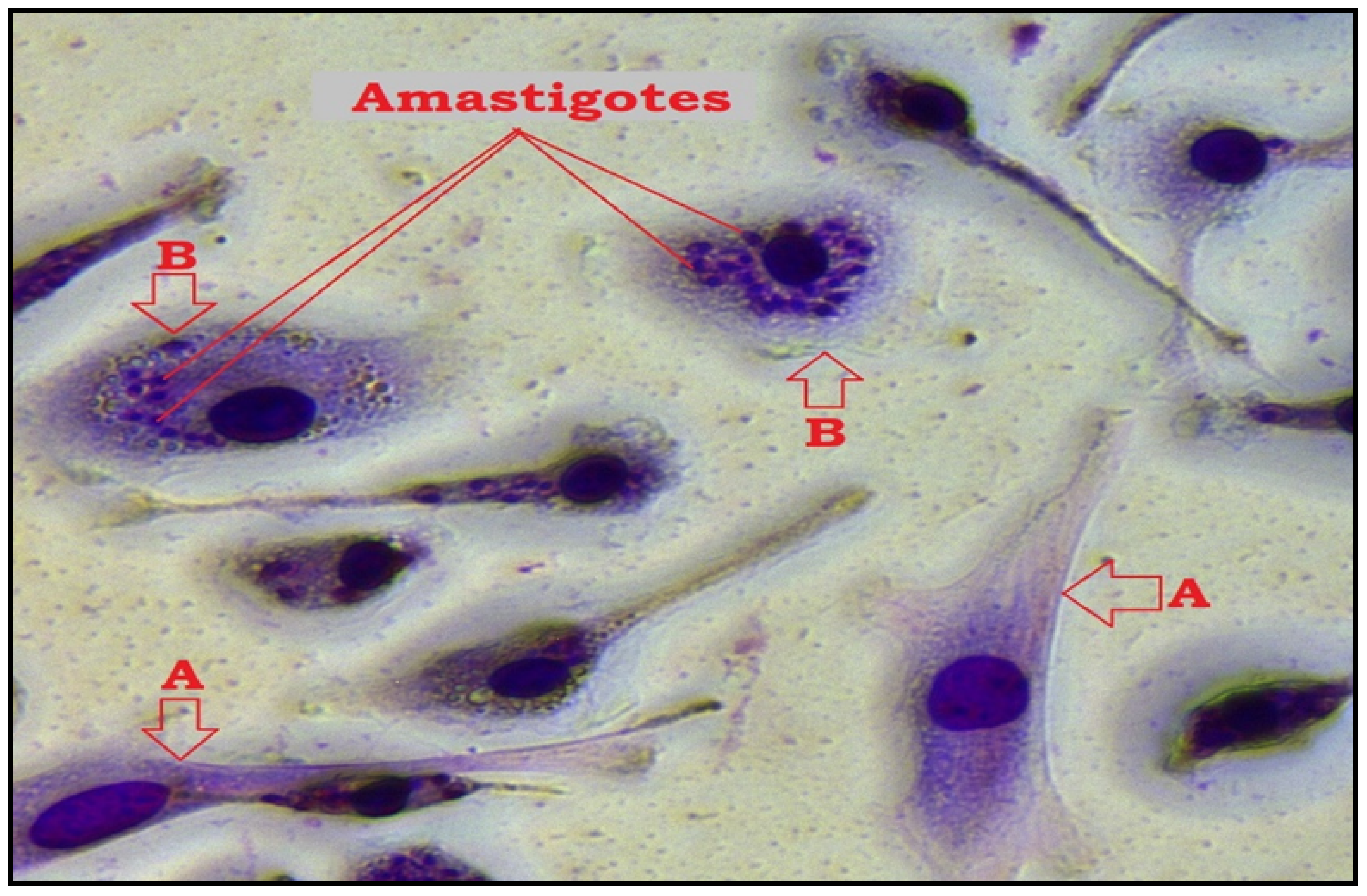
2.1.2. Antiamastigote Activity
2.2. In Silico Antileishmanial Activity of TQ
2.2.1. Molecular Docking Studies
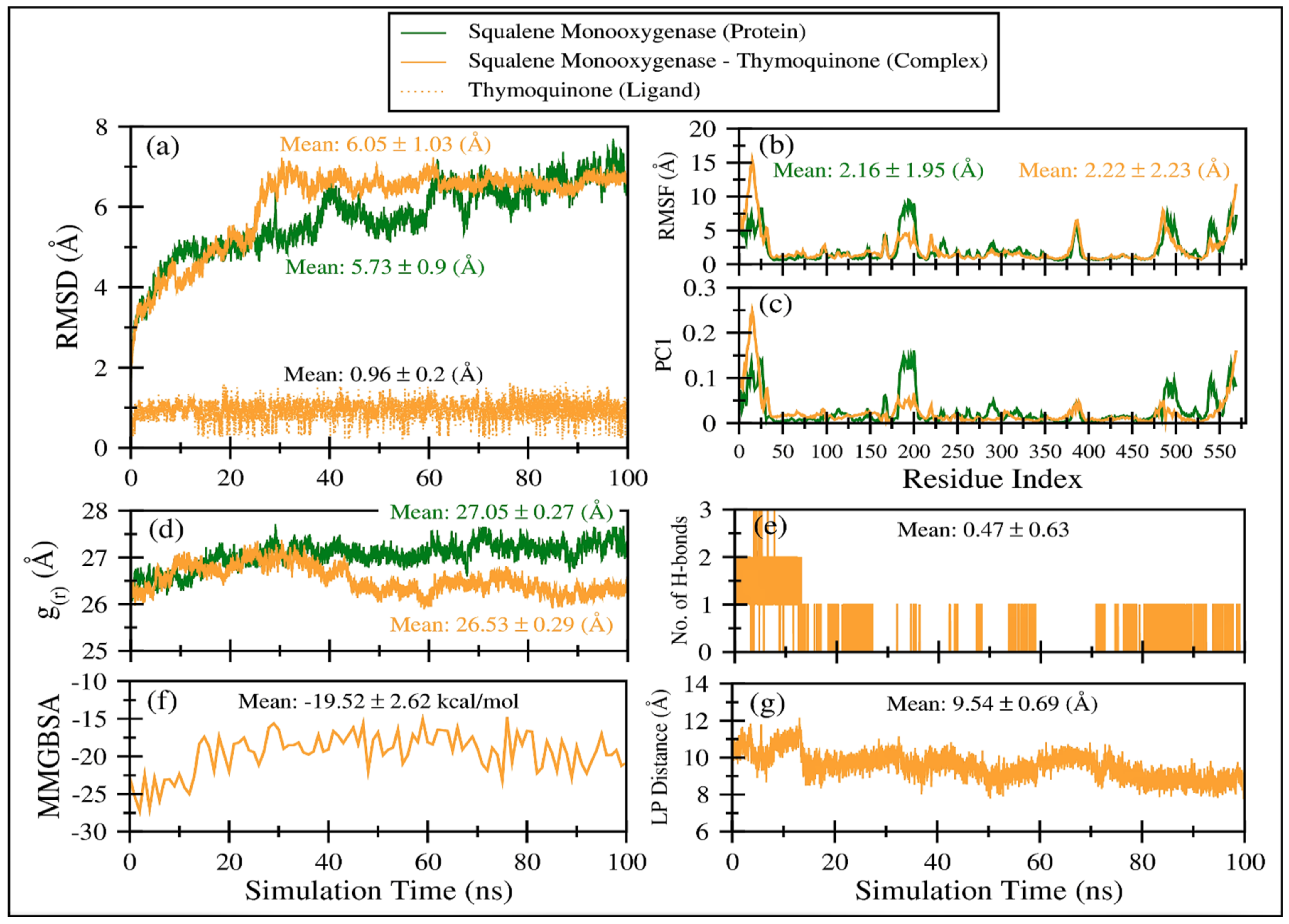
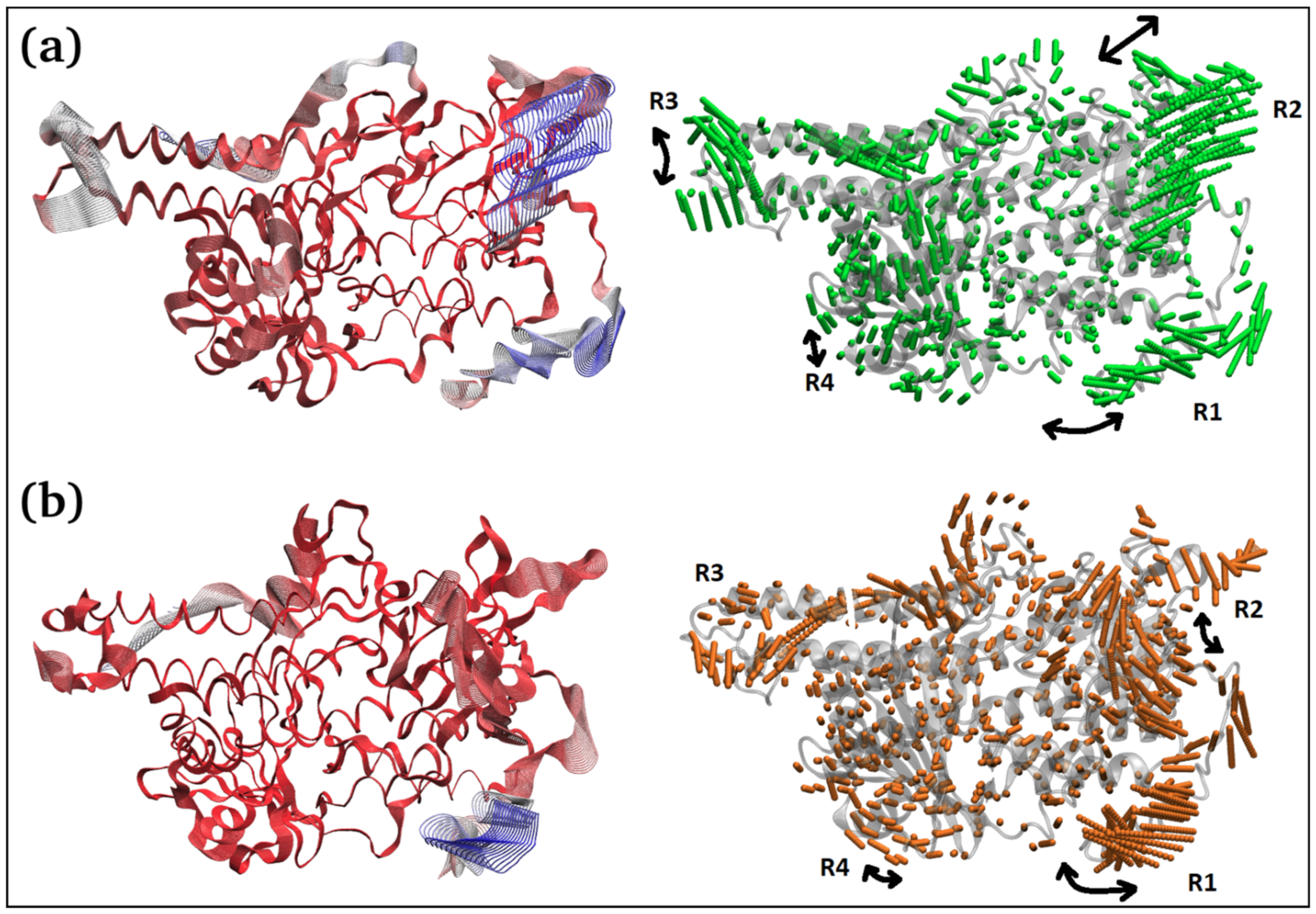
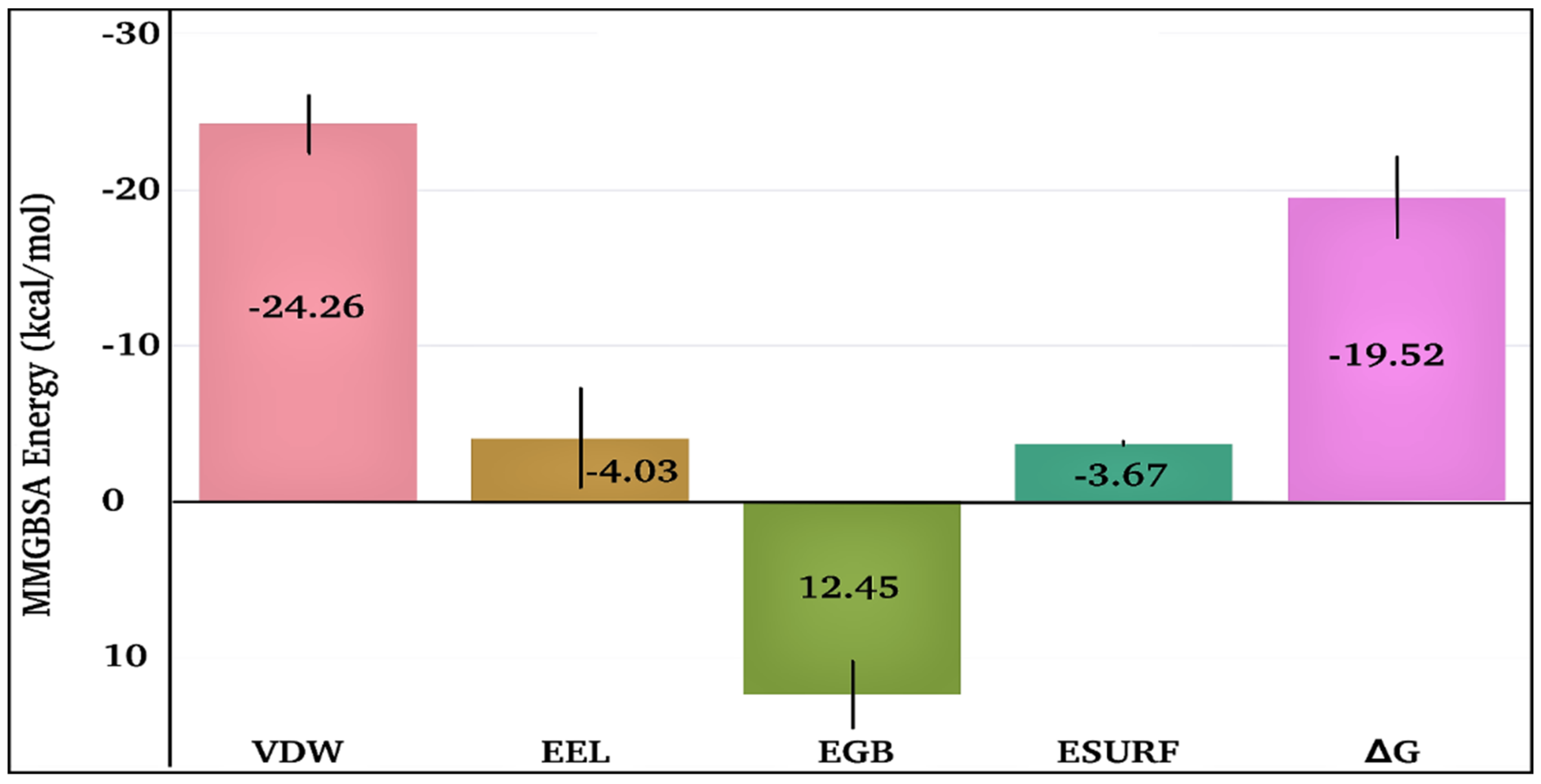
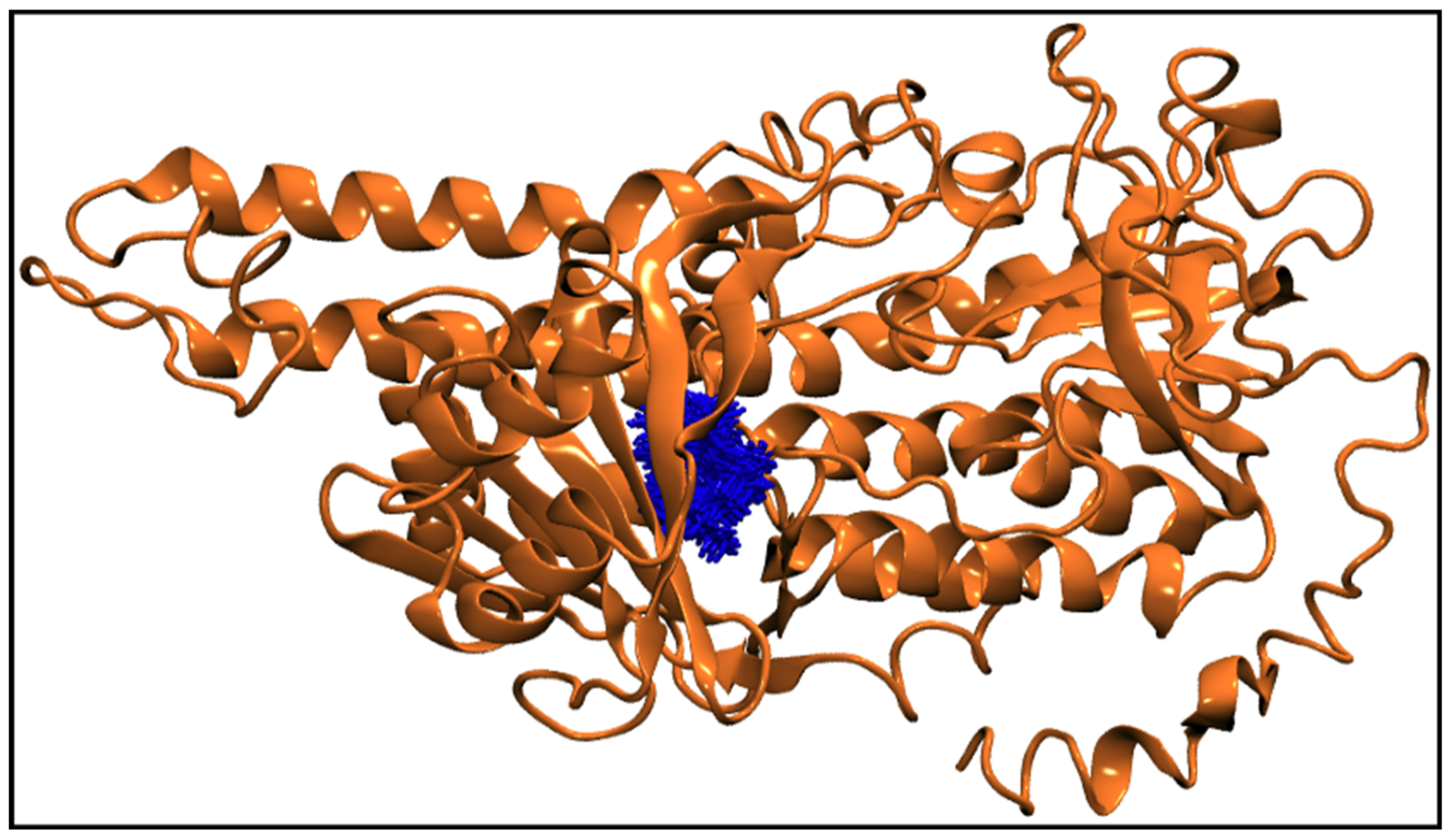
2.2.2. MD Simulations
2.3. Statistical Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Test Organism and Cultural Conditions
4.3. In Vitro Antileishmanial Activity of TQ
4.3.1. Antipromastigotes Activity
4.3.2. Antiamastigote Activity
4.3.3. In Vitro Cytotoxicity Activity
4.4. In Silico Antileishmanial Activity of TQ
4.4.1. Molecular Docking Studies
4.4.2. Molecular Dynamics (MD) Simulations
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chawla, B.; Madhubala, R. Drug targets in Leishmania. J. Parasit. Dis. 2010, 34, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Al Nasr, I.; Jentzsch, J.; Winter, I.; Schobert, R.; Ersfeld, K.; Koko, W.S.; Mujawah, A.A.H.; Khan, T.A.; Biersack, B. Antiparasitic activities of new lawsone Mannich bases. Arch. Pharm. 2019, 352, 1900128. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.K.; Prajapati, V.K.; Verma, G.K.; Chakraborty, D.; Sundar, S.; Rai, M.; Dubey, V.K.; Singh, M.S. Molecular docking and in vitro antileishmanial evaluation of chromene-2-thione analogues. ACS Med. Chem. Lett. 2012, 3, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.A.; Al Nasr, I.S.; Mujawah, A.H.; Koko, W.S. Assessment of Euphorbia retusa and Pulicaria undulata activity against Leishmania major and Toxoplasma gondii. Trop. Biomed. 2021, 38, 135–141. [Google Scholar] [CrossRef]
- Qureshi, K.A.; Al Nasr, I.; Koko, W.S.; Khan, T.A.; Fatmi, M.Q.; Imtiaz, M.; Khan, R.A.; Mohammed, H.A.; Jaremko, M.; Emwas, A.H.; et al. In vitro and in silico approaches for the antileishmanial activity evaluations of actinomycins isolated from novel Streptomyces smyrnaeus strain UKAQ_23. Antibiotics 2021, 10, 887. [Google Scholar] [CrossRef]
- Lodi, G.; Sannino, M.; Caterino, P.; Cannarozzo, G.; Bennardo, L.; Nisticò, S.P. Fractional CO2 laser-assisted topical rifamycin drug delivery in the treatment of pediatric cutaneous leishmaniasis. Pediatr. Dermatol. 2021, 38, 717–720. [Google Scholar] [CrossRef]
- Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.A.; Adlem, E.; Aert, R.; et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005, 309, 436–442. [Google Scholar] [CrossRef]
- Mohammed, H.A.A.; Ali, H.M.; Qureshi, K.A.; Alsharidah, M.; Kandil, Y.I.; Said, R.; Mohammed, S.A.A.; Al-Omar, M.S.; Al Rugaie, O.; Abdellatif, A.A.H.; et al. Comparative phytochemical profile and biological activity of four major medicinal halophytes from Qassim flora. Plants 2021, 10, 2208. [Google Scholar] [CrossRef]
- Mohammed, H.A.A.; Al-Omar, M.S.; Khan, R.A.; Mohammed, S.A.A.; Qureshi, K.A.; Abbas, M.M.; Al Rugaie, O.; Abd-Elmoniem, E.; Ahmad, A.M.; Kandil, Y.I. Chemical profile, antioxidant, antimicrobial, and anticancer activities of the water-ethanol extract of Pulicaria undulata growing in the oasis of central Saudi Arabian desert. Plants 2021, 10, 1811. [Google Scholar] [CrossRef]
- Amin, E.; Abdel-bakky, M.S.; Mohammed, H.A.; Chigrupati, S.; Qureshi, K.A.; Hassan, M.H.A. Phytochemical analysis and evaluation of the antioxidant and antimicrobial activities of five halophytes from Qassim flora. Polish J. Environ. Stud. 2022, 31, 1–8. [Google Scholar] [CrossRef]
- Abdallah, E.; Musa, K.; Qureshi, K.; Sadeek, A. Antimicrobial activity and antioxidant potential of the methanolic leaf extracts of three cultivars of date palm trees (Phoenix dactylifera) from Saudi Arabia. Med. Sci.|Int. Med. J. 2017, 6, 1. [Google Scholar] [CrossRef]
- Abdallah, E.M.; Qureshi, K.A.; Ali, A.M.H.; Elhassan, G.O. Evaluation of some biological properties of Saussurea costus crude root extract. Biosci. Biotechnol. Res. Commun. 2017, 10, 601–611. [Google Scholar] [CrossRef]
- Abdullah, E.M.; Qureshi, K.A.; Musa, K.H. Antimicrobial, antioxidant and phytochemical screening of lupin seeds (Lupinus termis Forrsk.) from Sudan. CIBTech J. Microbiol. 2017, 6, 1–8. [Google Scholar]
- Woo, C.C.; Kumar, A.P.; Sethi, G.; Tan, K.H.B. Thymoquinone: Potential cure for inflammatory disorders and cancer. Biochem. Pharmacol. 2012, 83, 443–451. [Google Scholar] [CrossRef]
- Goyal, S.N.; Prajapati, C.P.; Gore, P.R.; Patil, C.R.; Mahajan, U.B.; Sharma, C.; Talla, S.P.; Ojha, S.K. Therapeutic potential and pharmaceutical development of thymoquinone: A multitargeted molecule of natural origin. Front. Pharmacol. 2017, 8, 656. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, K.A.; Imtiaz, M.; Parvez, A.; Rai, P.K.; Jaremko, M.; Emwas, A.H.; Bholay, A.D.; Fatmi, M.Q. In vitro and in silico approaches for the evaluation of antimicrobial activity, time-kill kinetics, and anti-biofilm potential of thymoquinone (2-Methyl-5-propan-2-ylcyclohexa-2, 5-diene-1,4-dione) against selected human pathogens. Antibiotics 2022, 11, 79. [Google Scholar] [CrossRef]
- Forouzanfar, F.; Fazly Bazzaz, B.S.; Hosseinzadeh, H. Black cumin (Nigella sativa) and its constituent (thymoquinone): A review on antimicrobial effects. Iran. J. Basic Med. Sci. 2014, 17, 929–938. [Google Scholar] [CrossRef]
- Halawani, E. Antibacterial activity of thymoquinone and thymohydroquinone of Nigella sativa L. and their interaction with some antibiotics. Adv. Biol. Res. 2009, 3, 148–152. [Google Scholar]
- Darakhshan, S.; Bidmeshki Pour, A.; Hosseinzadeh Colagar, A.; Sisakhtnezhad, S. Thymoquinone and its therapeutic potentials. Pharmacol. Res. 2015, 95–96, 138–158. [Google Scholar] [CrossRef]
- Abamor, E.S.; Tosyali, O.A.; Bagirova, M.; Allahverdiyev, A. Nigella sativa oil entrapped polycaprolactone nanoparticles for leishmaniasis treatment. IET Nanobiotechnol. 2018, 12, 1018–1026. [Google Scholar] [CrossRef]
- Abamor, E.S.; Allahverdiyev, A.M. A nanotechnology based new approach for chemotherapy of cutaneous leishmaniasis: TIO2@AG nanoparticles—Nigella sativa oil combinations. Exp. Parasitol. 2016, 166, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Islamuddin, M.; Ali, A.; Afzal, O.; Ali, A.; Ali, I.; Altamimi, A.S.A.; Alamri, M.A.; Kato, K.; Parveen, S. Thymoquinone induced leishmanicidal effect via programmed cell death in Leishmania donovani. ACS Omega 2022, 7, 10718–10728. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudvand, H.; Tavakoli, R.; Sharififar, F.; Minaie, K.; Ezatpour, B.; Jahanbakhsh, S.; Sharifi, I. Leishmanicidal and cytotoxic activities of Nigella sativa and its active principle, thymoquinone. Pharm. Biol. 2015, 53, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Al-Turkmani, M.O.; Mokrani, L.; Soukkarieh, C. Antileishmanial apoptotic activity of Nigella sativa L. essential oil and thymoquinone triggers on Leishmania tropica. Indian J. Exp. Biol. 2020, 58, 7. [Google Scholar]
- Koko, W.S.; Al Nasr, I.S.; Khan, T.A.; Schobert, R.; Biersack, B. An Update on natural antileishmanial treatment options from plants, fungi and algae. Chem. Biodivers. 2022, 19, e202100542. [Google Scholar] [CrossRef]
- Morris, G.M.; Ruth, H.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Software news and updates AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef]
- Thymoquinone|C10H12O2—PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Thymoquinone (accessed on 2 March 2022).
- Gasteiger, J.; Marsili, M. Iterative partial equalization of orbital electronegativity-a rapid access to atomic charges. Tetrahedron 1980, 36, 3219–3228. [Google Scholar] [CrossRef]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational protein-ligand docking and virtual drug screening with the AutoDock suite. Nat. Protoc. 2016, 11, 905–919. [Google Scholar] [CrossRef]
- Martínez-Rosell, G.; Giorgino, T.; De Fabritiis, G. PlayMolecule ProteinPrepare: A web application for protein preparation for molecular dynamics simulations. J. Chem. Inf. Model. 2017, 57, 1511–1516. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI input generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindah, E. Gromacs: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; De Groot, B.L.; Grubmüller, H.; MacKerell, A.D. CHARMM36m: An improved force field for folded and intrinsically disordered proteins. Nat. Methods 2016, 14, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Vanommeslaeghe, K.; MacKerell, A.D. Automation of the CHARMM general force field (CGenFF) I: Bond perception and atom typing. J. Chem. Inf. Model. 2012, 52, 3144–3154. [Google Scholar] [CrossRef] [PubMed]
- Vanommeslaeghe, K.; Raman, E.P.; MacKerell, A.D. Automation of the CHARMM general force field (CGenFF) II: Assignment of bonded parameters and partial atomic charges. J. Chem. Inf. Model. 2012, 52, 3155–3168. [Google Scholar] [CrossRef]
- Genheden, S.; Kuhn, O.; Mikulskis, P.; Hoffmann, D.; Ryde, U. The normal-mode entropy in the MM/GBSA method: Effect of system truncation, buffer region, and dielectric constant. J. Chem. Inf. Model. 2012, 52, 2079–2088. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Antipromastigote Activity | ||
|---|---|---|---|
| EC50 (μM) | CC50 (μM) | SI | |
| TQ | 2.62 ± 0.12 | 29.54 ± 0.07 | 11.27 |
| CTL-D | 0.84 ± 0.04 | 8.01 ± 0.29 | 9.54 |
| Compounds | Antiamastigote Activity | ||
|---|---|---|---|
| EC50 (μM) | CC50 (μM) | SI | |
| TQ | 17.52 ± 0.15 | 29.54 ± 0.07 | 1.69 |
| CTL-D | 0.50 ± 0.06 | 8.01 ± 0.29 | 16.02 |
| # | Enzymes | Pathway | Binding Energy (kcal/mol) |
|---|---|---|---|
| 1. | Squalene monooxygenase | Sterol biogenetic pathway | −7.1 |
| 2. | Fructose-bisphosphate aldolase | Glycolytic pathway | −6.8 |
| 3. | Mannosyltransferase (GPI-14) | Glycosylphosphatidylinositol-anchor biosynthesis | −6.8 |
| 4. | Trypanothione reductase | Trypanothione pathway | −6.6 |
| 5. | Trypanothione synthetase-amidase | Trypanothione pathway | −6.4 |
| 6. | Phosphoglycerate kinase | Glycolytic pathway | −6.3 |
| 7. | Deoxyhypusine hydroxylase | Hypusine biosynthetic | −6.3 |
| 8. | Adenine phosphoribosyltransferase | Purine salvage pathway | −6.2 |
| 9. | Xanthine phosphoribosyltransferase | Purine salvage pathway | −6.2 |
| 10. | Squalene synthase | Sterol biogenetic pathway | −6.1 |
| 11. | Farnesyl pyrophosphate synthase | Sterol biogenetic pathway | −5.9 |
| 12. | Pyruvate kinase | Glycolytic pathway | −5.9 |
| 13. | Tryparedoxin peroxidase | Trypanothione pathway | −5.8 |
| 14. | Phosphoglycerate mutase (2,3-diphosphoglycerate-independent) | Glycolytic pathway | −5.8 |
| 15. | Glyceraldehyde-3-phosphate dehydrogenase | Glycolytic pathway | −5.6 |
| 16. | Triosephosphate isomerase | Glycolytic pathway | −5.1 |
| Independent Samples Test | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Levene’s Test for Equality of Variances | t-Test for Equality of Means | |||||||||
| F | Sig. | t | df | Sig. (2-Tailed) | Mean Difference | Std. Error Difference | 95% Confidence Interval of the Difference | |||
| Lower | Upper | |||||||||
| EC50 | Equal variances assumed | 0.203 | 0.676 | −131.830 | 4 | 0.000 | −2.44667 | 0.01856 | −2.49820 | −2.39514 |
| Equal variances not assumed | −131.830 | 3.806 | 0.000 | −2.44667 | 0.01856 | −2.49925 | −2.39409 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qureshi, K.A.; Imtiaz, M.; Al Nasr, I.; Koko, W.S.; Khan, T.A.; Jaremko, M.; Mahmood, S.; Fatmi, M.Q. Antiprotozoal Activity of Thymoquinone (2-Isopropyl-5-methyl-1,4-benzoquinone) for the Treatment of Leishmania major-Induced Leishmaniasis: In Silico and In Vitro Studies. Antibiotics 2022, 11, 1206. https://doi.org/10.3390/antibiotics11091206
Qureshi KA, Imtiaz M, Al Nasr I, Koko WS, Khan TA, Jaremko M, Mahmood S, Fatmi MQ. Antiprotozoal Activity of Thymoquinone (2-Isopropyl-5-methyl-1,4-benzoquinone) for the Treatment of Leishmania major-Induced Leishmaniasis: In Silico and In Vitro Studies. Antibiotics. 2022; 11(9):1206. https://doi.org/10.3390/antibiotics11091206
Chicago/Turabian StyleQureshi, Kamal A., Mahrukh Imtiaz, Ibrahim Al Nasr, Waleed S. Koko, Tariq A. Khan, Mariusz Jaremko, Syed Mahmood, and M. Qaiser Fatmi. 2022. "Antiprotozoal Activity of Thymoquinone (2-Isopropyl-5-methyl-1,4-benzoquinone) for the Treatment of Leishmania major-Induced Leishmaniasis: In Silico and In Vitro Studies" Antibiotics 11, no. 9: 1206. https://doi.org/10.3390/antibiotics11091206