
Both Manuka and Non-Manuka Honey Types Inhibit Antibiotic Resistant Wound-Infecting Bacteria


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
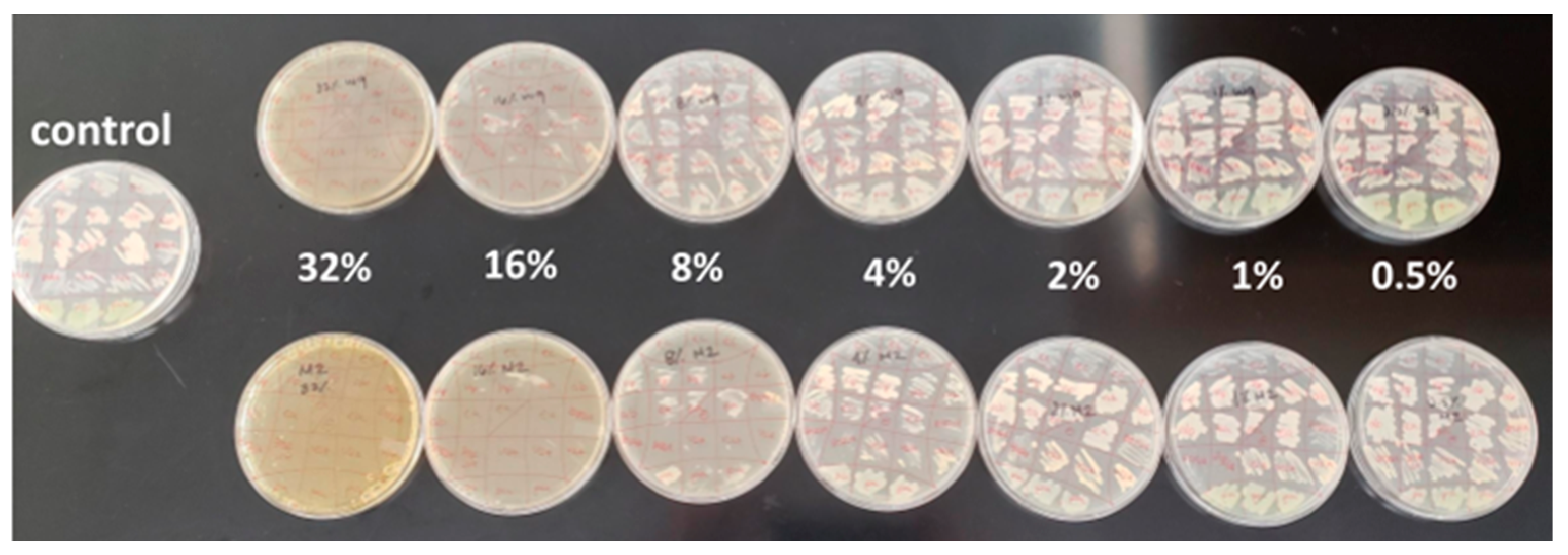
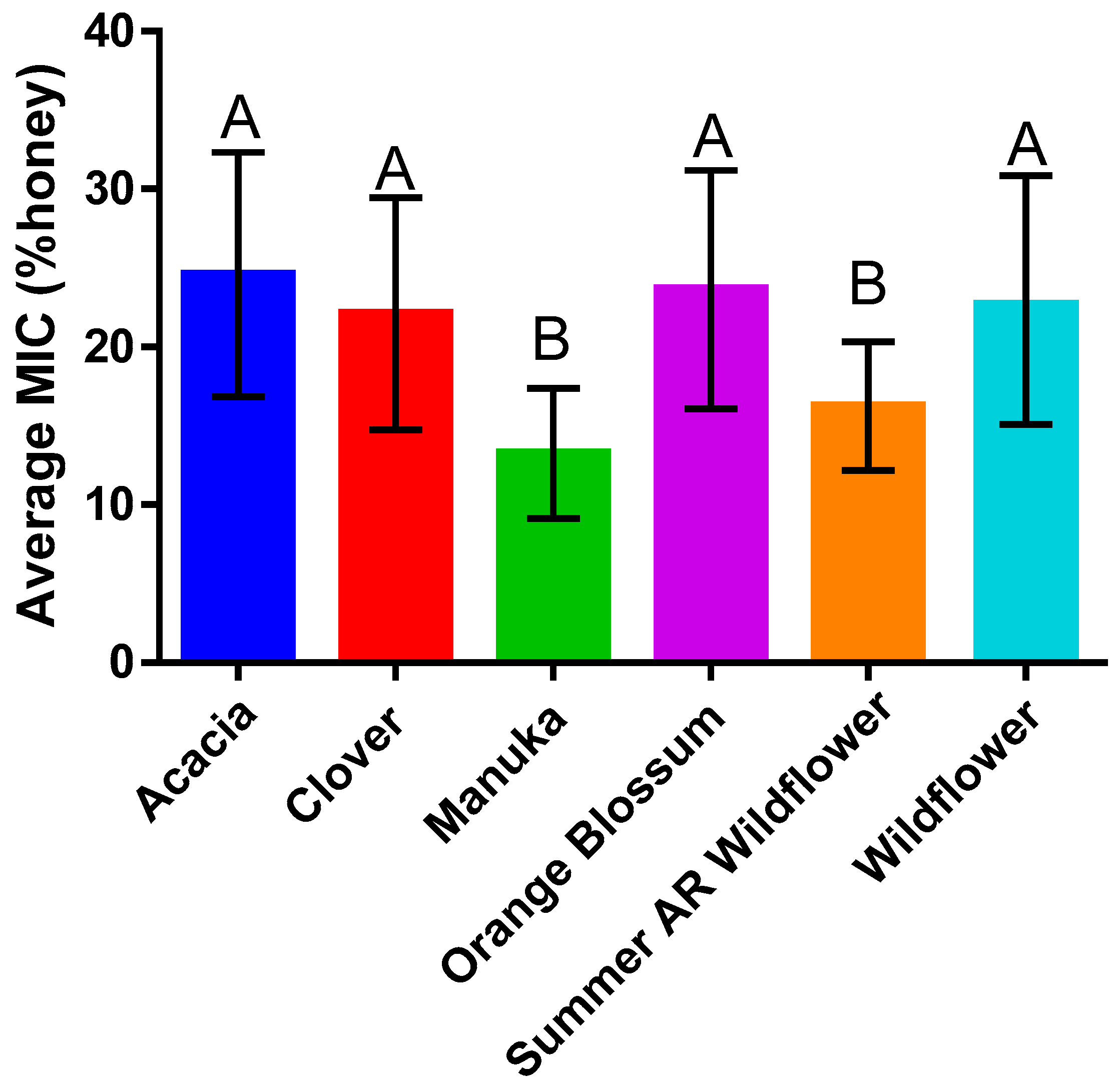
2.1. Antibacterial Efficacy Assays
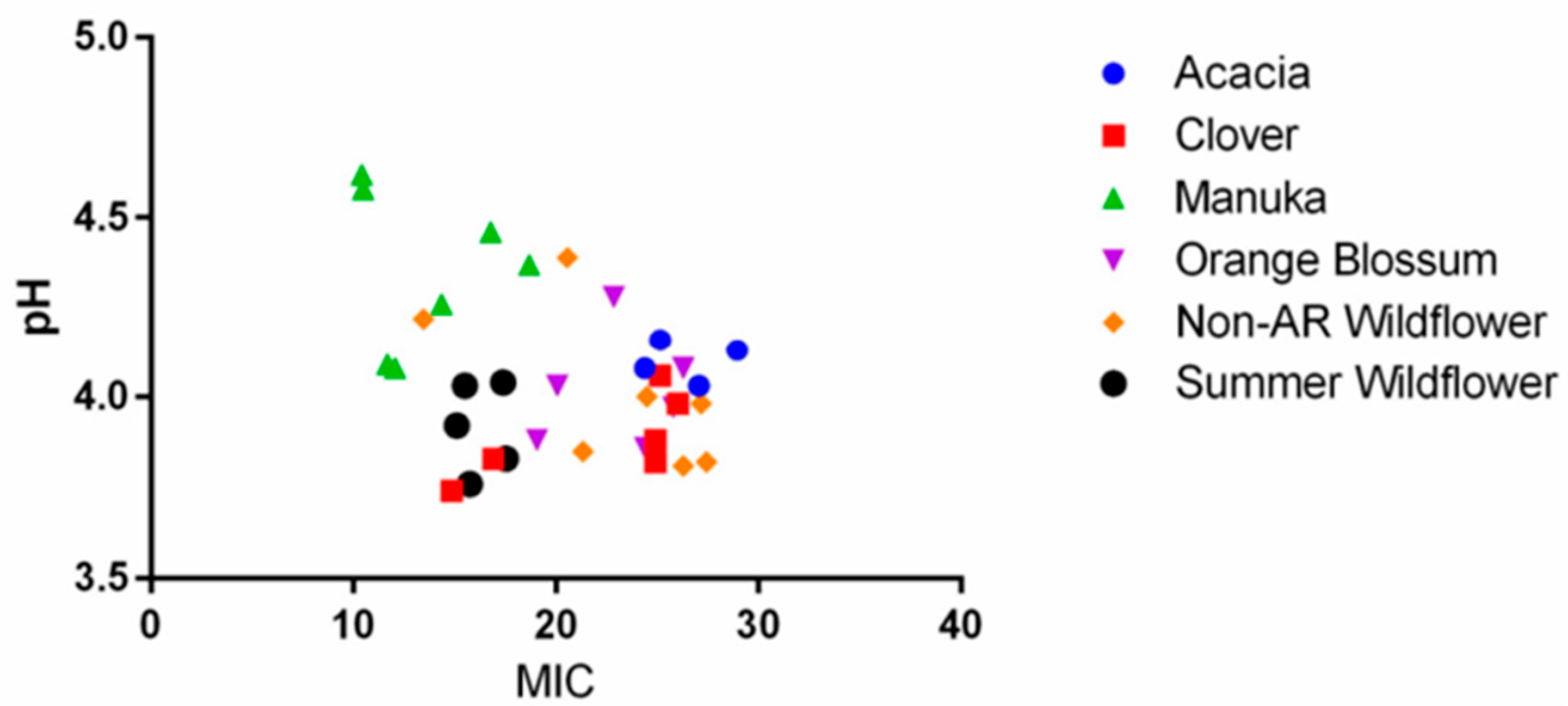
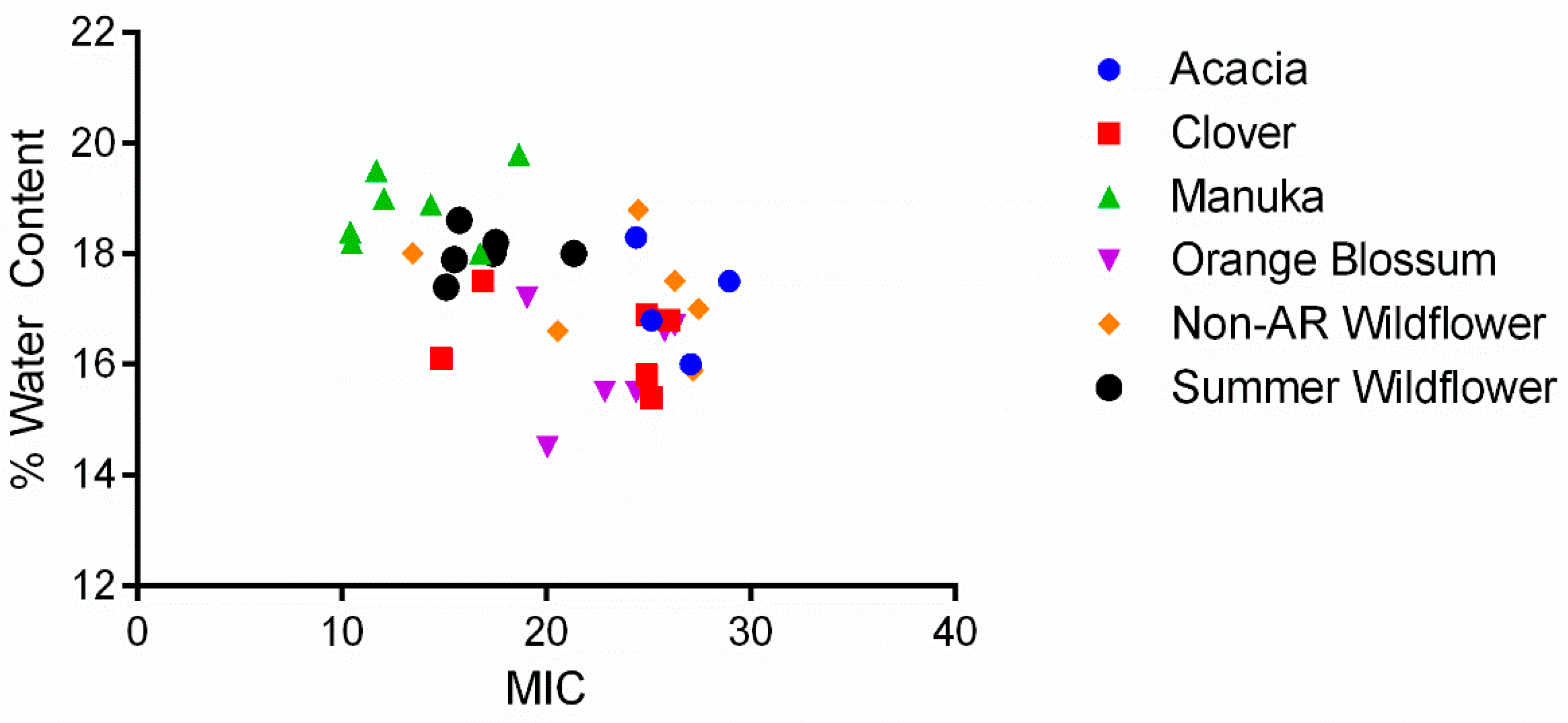
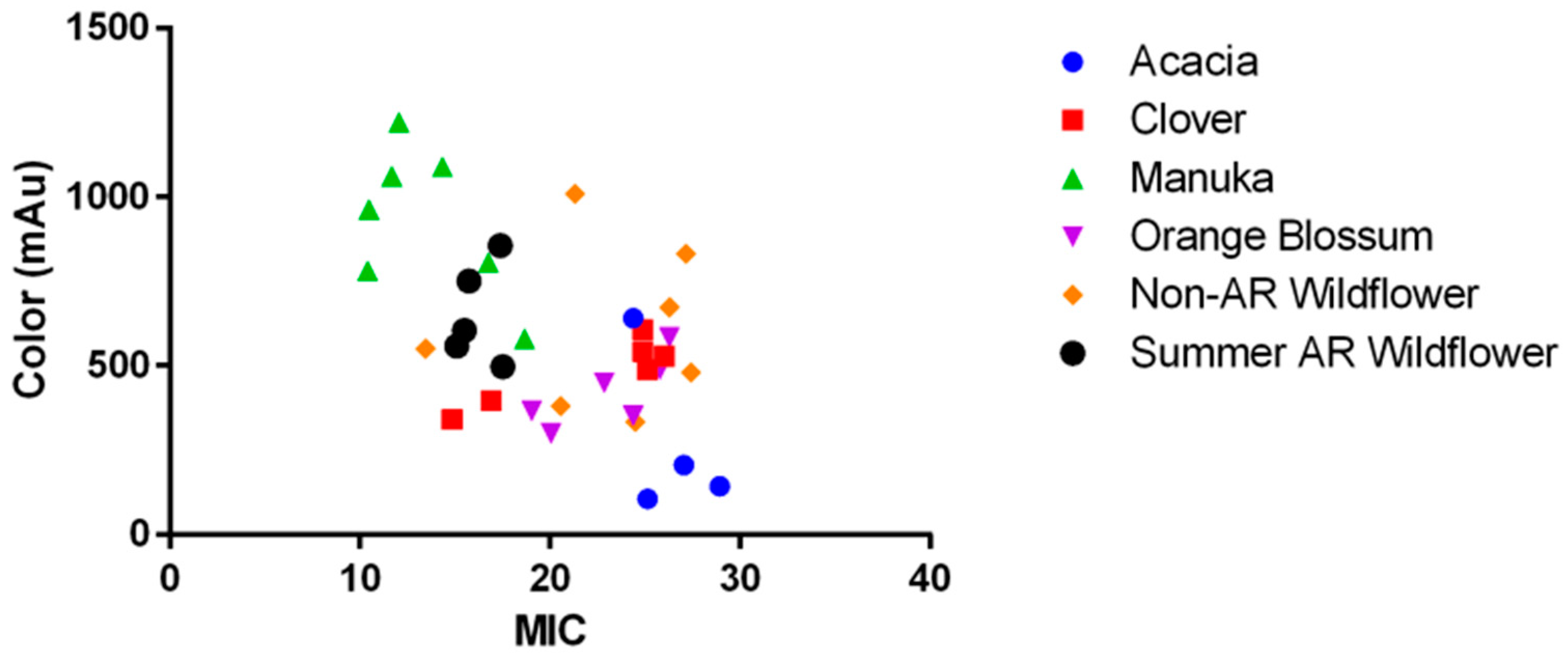
2.2. The Role of pH, Osmolarity, and Color in Honey Antibacterial Efficacy
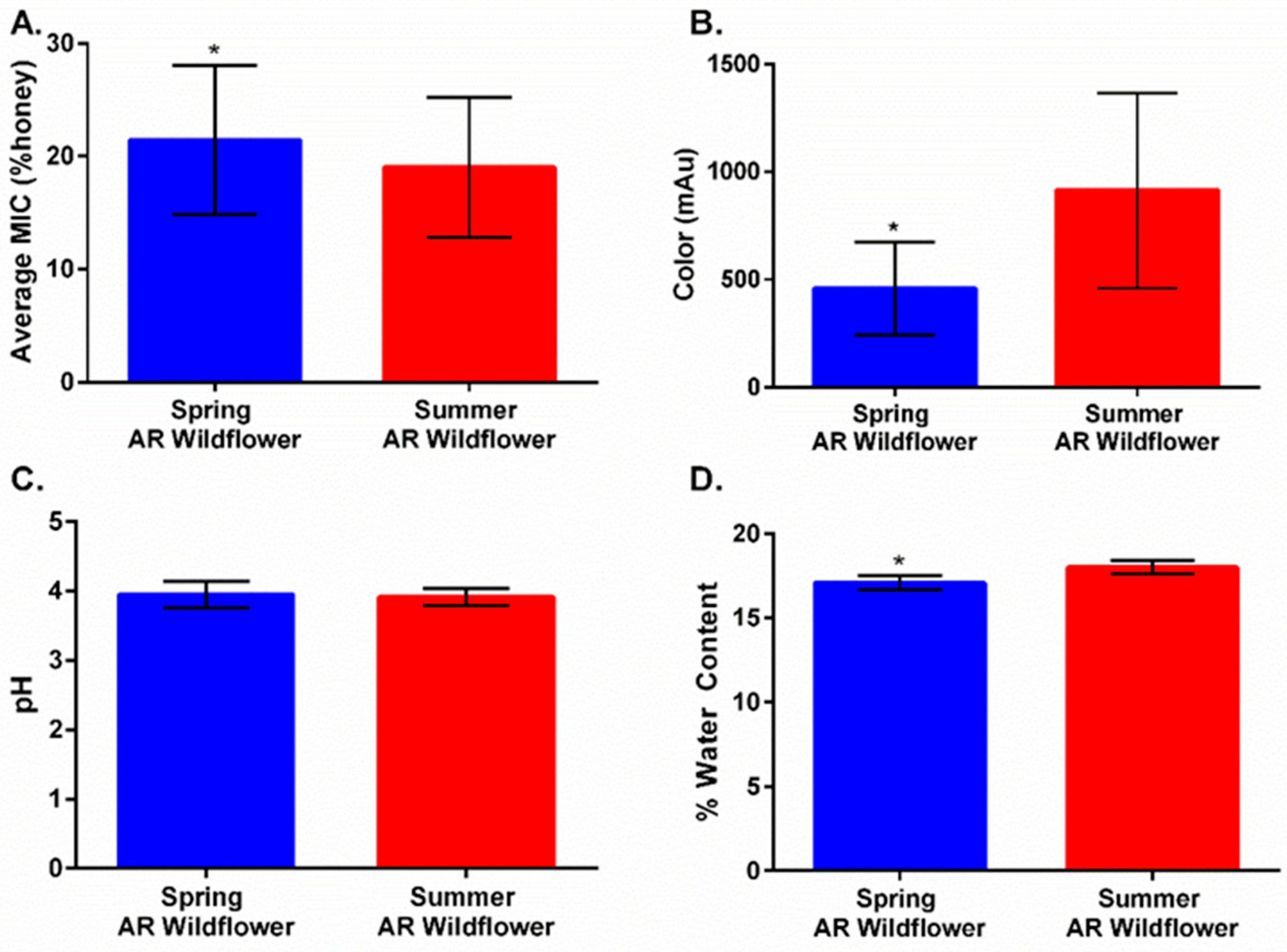
2.3. Harvest Timing Determines Antimicrobial Activity
2.4. Honey Type Efficacy Is Species-Dependent
3. Conclusions
4. Materials and Methods
4.1. Microorganisms
4.2. Honey Sample Acquisition and Processing
4.3. Honey pH
4.4. Osmolarity Determinations
4.5. Honey Color
4.6. Honey Type
4.7. Minimum Inhibitory Concentration (MIC) Assays
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate Point-Prevalence Survey of Health Care–Associated Infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.J.; Edwards, J.R.; Sievert, D.M. Antimicrobial-Resistant Pathogens Associated With Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Infect. Control Hosp. Epidemiol. 2016, 37, 1288–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molan, P. Honey: Antimicrobial Actions and Role in Disease Management. In New Stratagies Combating Bacterial Infection; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008; pp. 229–253. ISBN 978-3-527-62293-1. [Google Scholar]
- Hussain, M.B. Role of Honey in Topical and Systemic Bacterial Infections. J. Altern. Complement. Med. 2018, 24, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, J. The Production of Honey. Bee World 1925, 7, 95–98. [Google Scholar] [CrossRef]
- Crane, E. Bee Research Association In Honey: A Comprehensive Survey; Heinemann [for] the Bee Research Association: London, UK, 1975; pp. 157–206. ISBN 978-0-434-90270-5. [Google Scholar]
- Brudzynski, K.; Abubaker, K.; St-Martin, L.; Castle, A. Re-Examining the Role of Hydrogen Peroxide in Bacteriostatic and Bactericidal Activities of Honey. Front. Microbiol. 2011, 2, 213. [Google Scholar] [CrossRef] [Green Version]
- Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.-Y. Use of Hydrogen Peroxide as a Biocide: New Consideration of Its Mechanisms of Biocidal Action. J. Antimicrob. Chemother. 2012, 67, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Mandal, M.D.; Mandal, S. Honey: Its Medicinal Property and Antibacterial Activity. Asian Pac. J. Trop. Biomed. 2011, 1, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Molan, P.; Rhodes, T. Honey: A Biologic Wound Dressing. Wounds 2015, 27, 141–151. [Google Scholar]
- Brudzynski, K.; Lannigan, R. Mechanism of Honey Bacteriostatic Action Against MRSA and VRE Involves Hydroxyl Radicals Generated from Honey’s Hydrogen Peroxide. Front. Microbiol. 2012, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Brackman, G.; De Meyer, L.; Nelis, H.J.; Coenye, T. Biofilm Inhibitory and Eradicating Activity of Wound Care Products against Staphylococcus Aureus and Staphylococcus Epidermidis Biofilms in an in Vitro Chronic Wound Model. J. Appl. Microbiol. 2013, 114, 1833–1842. [Google Scholar] [CrossRef]
- Majtan, J.; Bohova, J.; Horniackova, M.; Klaudiny, J.; Majtan, V. Anti-Biofilm Effects of Honey against Wound Pathogens Proteus Mirabilis and Enterobacter Cloacae. Phytother. Res. 2014, 28, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Park, J.-H.; Kim, J.-A.; Neupane, G.P.; Cho, M.H.; Lee, C.-S.; Lee, J. Low Concentrations of Honey Reduce Biofilm Formation, Quorum Sensing, and Virulence in Escherichia Coli O157:H7. Biofouling 2011, 27, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; López-Gálvez, F.; Gil, M.I.; Tomás-Barberán, F.A.; Allende, A. Quorum Sensing Inhibitory and Antimicrobial Activities of Honeys and the Relationship with Individual Phenolics. Food Chem. 2009, 115, 1337–1344. [Google Scholar] [CrossRef]
- Wang, R.; Starkey, M.; Hazan, R.; Rahme, L.G. Honey’s Ability to Counter Bacterial Infections Arises from Both Bactericidal Compounds and QS Inhibition. Front. Microbiol. 2012, 3, 144. [Google Scholar] [CrossRef] [Green Version]
- Robson, V.; Dodd, S.; Thomas, S. Standardized Antibacterial Honey (MedihoneyTM) with Standard Therapy in Wound Care: Randomized Clinical Trial. J. Adv. Nurs. 2009, 65, 565–575. [Google Scholar] [CrossRef]
- Lusby, P.E.; Coombes, A.L.; Wilkinson, J.M. Bactericidal Activity of Different Honeys against Pathogenic Bacteria. Arch. Med. Res. 2005, 36, 464–467. [Google Scholar] [CrossRef]
- Molan, P. An Explanation of Why the MGO Level in Manuka Honey Does Not Show the Antibacterial Activity. N. Z. Beekeep. 2008, 16, 11–13. [Google Scholar]
- Bucekova, M.; Sojka, M.; Valachova, I.; Martinotti, S.; Ranzato, E.; Szep, Z.; Majtan, V.; Klaudiny, J.; Majtan, J. Bee-Derived Antibacterial Peptide, Defensin-1, Promotes Wound Reepithelialisation in Vitro and in Vivo. Sci. Rep. 2017, 7, 7340. [Google Scholar] [CrossRef] [Green Version]
- WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 25 July 2022).
- National Honey Board Honey: A Reference Guide to Nature’s Sweetener. Available online: https://honey.com/images/files/Detailed-Nutrition-Information.pdf (accessed on 26 July 2022).
- Roshan, N.; Rippers, T.; Locher, C.; Hammer, K.A. Antibacterial Activity and Chemical Characteristics of Several Western Australian Honeys Compared to Manuka Honey and Pasture Honey. Arch. Microbiol. 2017, 199, 347–355. [Google Scholar] [CrossRef]
- Anand, S.; Pang, E.; Livanos, G.; Mantri, N. Characterization of Physico-Chemical Properties and Antioxidant Capacities of Bioactive Honey Produced from Australian Grown Agastache Rugosa and Its Correlation with Colour and Poly-Phenol Content. Molecules 2018, 23, 108. [Google Scholar] [CrossRef] [Green Version]
- Brudzynski, K.; Adubaker, K.; Miotto, D. Unraveling a Mechanism of Honey Antibacterial Action: Polyphenol/H2O2-Induced Oxidative Effect on Bacterial Cell Growth and on DNA Degradation. Food Chem. 2012, 133, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Grainger, M.N.C.; Manley-Harris, M.; Lane, J.R.; Field, R.J. Kinetics of Conversion of Dihydroxyacetone to Methylglyoxal in New Zealand Mānuka Honey: Part I—Honey Systems. Food Chem. 2016, 202, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Cokcetin, N.N.; Lu, J.; Turnbull, L.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Rifampicin-Manuka Honey Combinations Are Superior to Other Antibiotic-Manuka Honey Combinations in Eradicating Staphylococcus Aureus Biofilms. Front. Microbiol. 2018, 8, 2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soffer, L. Arkansas Native Wildflowers. Available online: https://www.uaex.uada.edu/yard-garden/master-gardeners/Arkansas%20Native%20Wildflowers.pdf (accessed on 26 July 2022).
- USDA. Plants Database. Available online: https://plants.usda.gov/home (accessed on 11 July 2022).
- CDC. 2016 HAI Progress Report. Available online: https://www.cdc.gov/hai/data/archive/2016-HAI-progress-report.html (accessed on 13 July 2022).
- HAI|CDC. Acinetobacter in Healthcare Settings. Available online: https://www.cdc.gov/hai/organisms/acinetobacter.html (accessed on 13 July 2022).
- Klevens, R.M.; Edwards, J.R.; Richards, C.L.; Horan, T.C.; Gaynes, R.P.; Pollock, D.A.; Cardo, D.M. Estimating Health Care-Associated Infections and Deaths in U.S. Hospitals, 2002. Public Health Rep. 2007, 122, 160–166. [Google Scholar] [CrossRef]
- CDC. What CDC Is Doing: Antibiotic Resistance (AR) Solutions Initiative. Available online: https://www.cdc.gov/drugresistance/solutions-initiative/index.html (accessed on 13 July 2022).
- Jenkins, R.; Burton, N.; Cooper, R. Proteomic and Genomic Analysis of Methicillin-Resistant Staphylococcus Aureus (MRSA) Exposed to Manuka Honey in Vitro Demonstrated down-Regulation of Virulence Markers. J. Antimicrob. Chemother. 2014, 69, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Molan, P.C. The Evidence and the Rationale for the Use of Honey as a Wound Dressing. Wound Pract. Res. 2011, 19, 204–220. [Google Scholar]
- Aamer, A.A.; Abdul-Hafeez, M.M.; Sayed, S.M. Minimum Inhibitory and Bactericidal Concentrations (MIC and MBC) of Honey and Bee Propolis against Multi-Drug Resistant (MDR) Staphylococcus Sp. Isolated from Bovine Clinical Mastitis. Altern. Integr. Med. 2014, 3, 4. [Google Scholar] [CrossRef]
- Cooper, R.A.; Jenkins, L.; Henriques, A.F.M.; Duggan, R.S.; Burton, N.F. Absence of Bacterial Resistance to Medical-Grade Manuka Honey. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1237–1241. [Google Scholar] [CrossRef] [Green Version]
- Chris, D. New Klebsiella Strains “Worst-Case Scenario,” Experts Say. 2017. Available online: https://www.cidrap.umn.edu/news-perspective/2017/09/new-klebsiella-strains-worst-case-scenario-experts-say (accessed on 11 July 2022).
- Bogdanov, S. Harmonised Methods of the International Honey Commission. Available online: https://www.ihc-platform.net/ihcmethods2009.pdf (accessed on 25 July 2022).
- Beretta, G.; Granata, P.; Farrero, M.; Orioli, M.; Facino, R.M. Standardization of Antioxidant Properties of Honey by a Combination of Spectrophotometric/Fluorimetric Assays and Chemometrics. Anal. Chim. Acta 2005, 533, 185–191. [Google Scholar] [CrossRef]
- Hewett, S.R. Both Manuka and Non-Manuka Honey Types Inhibit Antibiotic Resistant Wound-Infecting Bacteria. Master’s Thesis, University of Central Arkansas, Conway, AR, USA, 2019. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hewett, S.R.; Crabtrey, S.D.; Dodson, E.E.; Rieth, C.A.; Tarkka, R.M.; Naylor, K. Both Manuka and Non-Manuka Honey Types Inhibit Antibiotic Resistant Wound-Infecting Bacteria. Antibiotics 2022, 11, 1132. https://doi.org/10.3390/antibiotics11081132
Hewett SR, Crabtrey SD, Dodson EE, Rieth CA, Tarkka RM, Naylor K. Both Manuka and Non-Manuka Honey Types Inhibit Antibiotic Resistant Wound-Infecting Bacteria. Antibiotics. 2022; 11(8):1132. https://doi.org/10.3390/antibiotics11081132
Chicago/Turabian StyleHewett, Samantha R., Stephany D. Crabtrey, Esther E. Dodson, C. Alexander Rieth, Richard M. Tarkka, and Kari Naylor. 2022. "Both Manuka and Non-Manuka Honey Types Inhibit Antibiotic Resistant Wound-Infecting Bacteria" Antibiotics 11, no. 8: 1132. https://doi.org/10.3390/antibiotics11081132