
Antibacterial Activity against Clinical Isolates and In Vivo Efficacy of Coralmycins

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. In Vitro Antibacterial Activities
2.2. Time-Kill Study
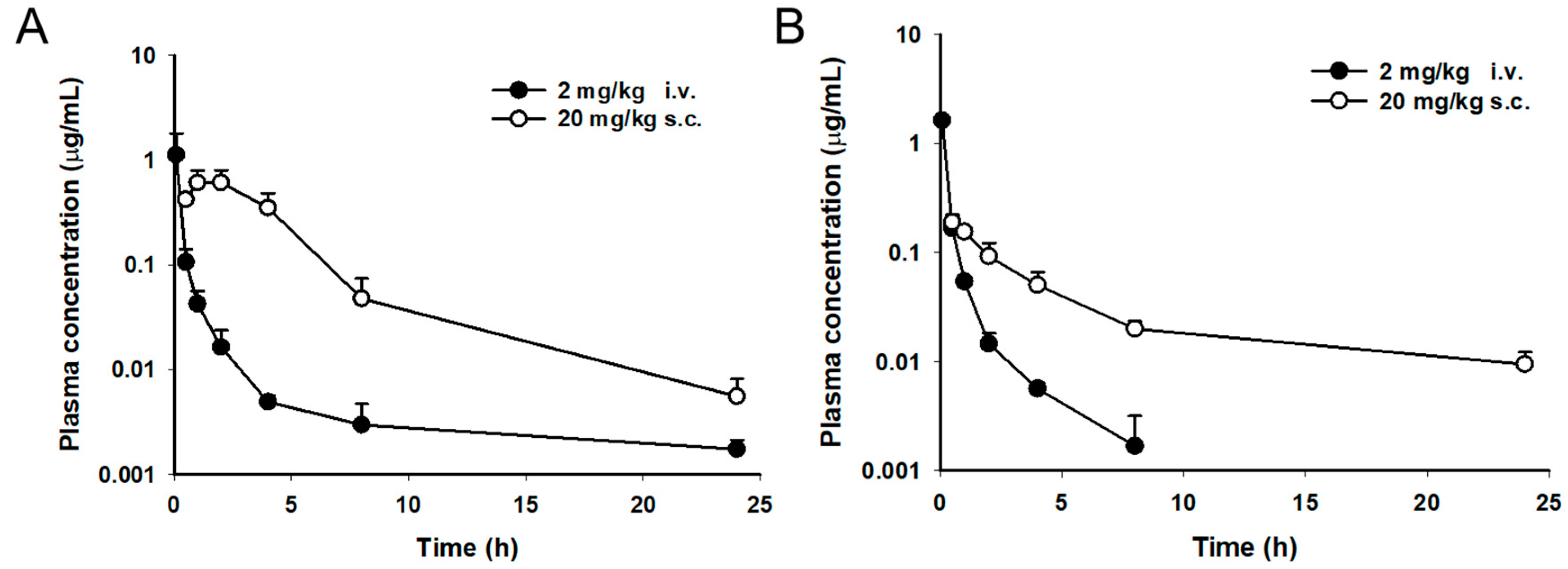
2.3. Pharmacokinetic Study
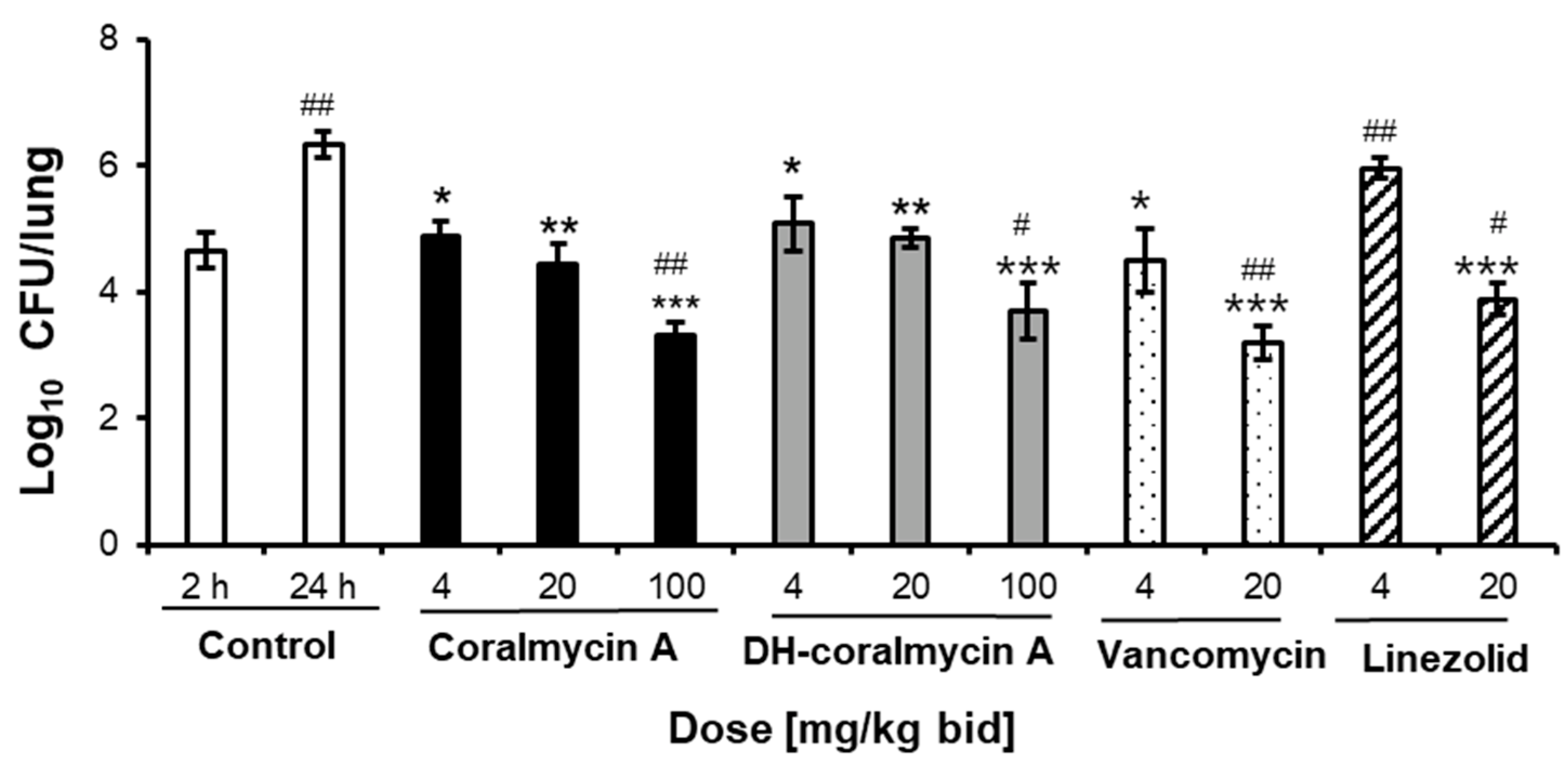
2.4. In Vivo Efficacy
3. Discussion
4. Materials and Methods
4.1. Antimicrobial Agents and Reagents
4.2. Bacterial Strains
4.3. Susceptibility Testing
4.4. Time-Kill Assay
4.5. Animals
4.6. Pharmacokinetic Study
4.7. In Vitro Plasma Protein-Binding Studies
4.8. In Vivo Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nature 2017, 543, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiodras, S.; Gold, H.S.; Sakoulas, G.; Eliopoulos, G.M.; Wennersten, C.; Venkataraman, L.; Moellering, R.C.; Ferraro, M.J. Linezolid resistance in a clinical isolate of Staphylococcus aureus. Lancet 2001, 358, 207–208. [Google Scholar] [CrossRef]
- Wilson, P.; Andrews, J.A.; Charlesworth, R.; Walesby, R.; Singer, M.; Farrell, D.J.; Robbins, M. Linezolid resistance in clinical isolates of Staphylococcus aureus. J. Antimicrob. Chemother. 2003, 51, 186–188. [Google Scholar] [CrossRef]
- Azhar, A.; Rasool, S.; Haque, A.; Shan, S.; Saeed, M.; Ehsan, B.; Haque, A. Detection of high levels of resistance to linezolid and vancomycin in Staphylococcus aureus. J. Med. Microbiol. 2017, 66, 1328–1331. [Google Scholar] [CrossRef]
- Mittal, G.; Bhandari, V.; Gaind, R.; Rani, V.; Chopra, S.; Dawar, R.; Sardana, R.; Verma, P.K. Linezolid resistant coagulase negative staphylococci (LRCoNS) with novel mutations causing blood stream infections (BSI) in India. BMC Infect. Dis. 2019, 19, 717. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Gulati, N.; Sharma, S.; Gupta, R.; Soni, A.; Chander, J. Linezolid Resistance in Staphylococcus haemolyticus—Case Series and Review of Literature. Infect. Disord. Drug Targets 2020, 20, 713–717. [Google Scholar] [CrossRef]
- Lee, C.H.; Wang, M.C.; Huang, I.W.; Chen, F.J.; Lauderdale, T.L. Development of daptomycin nonsusceptibility with heterogeneous vancomycin-intermediate resistance and oxacillin susceptibility in methicillin-resistant Staphylococcus aureus during high-dose daptomycin treatment. Antimicrob. Agents Chemother. 2010, 54, 4038–4040. [Google Scholar] [CrossRef] [Green Version]
- Humphries, R.M.; Pollett, S.; Sakoulas, G. A current perspective on daptomycin for the clinical microbiologist. Clin. Microbiol. Rev. 2013, 26, 759–780. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Kim, H.J.; Kim, G.W.; Cho, K.; Takahashi, S.; Koshino, H.; Kim, W.G. Isolation of Coralmycins A and B, Potent Anti-Gram Negative Compounds from the Myxobacteria Corallococcus coralloides M23. J. Nat. Prod. 2016, 79, 2223–2228. [Google Scholar] [CrossRef]
- Baumann, S.; Herrmann, J.; Raju, R.; Steinmetz, H.; Mohr, K.I.; Huttel, S.; Harmrolfs, K.; Stadler, M.; Muller, R. Cystobactamids: Myxobacterial topoisomerase inhibitors exhibiting potent antibacterial activity. Angew. Chem. Int. Ed. Engl. 2014, 53, 14605–14609. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Minh, N.V.; Choi, H.Y.; Kim, W.G. Coralmycin Derivatives with Potent Anti-Gram Negative Activity Produced by the Myxobacteria Corallococcus coralloides M23. Molecules 2019, 24, 1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttel, S.; Testolin, G.; Herrmann, J.; Planke, T.; Gille, F.; Moreno, M.; Stadler, M.; Bronstrup, M.; Kirschning, A.; Muller, R. Discovery and Total Synthesis of Natural Cystobactamid Derivatives with Superior Activity against Gram-Negative Pathogens. Angew. Chem. Int. Ed. Engl. 2017, 56, 12760–12764. [Google Scholar] [CrossRef]
- Cheng, B.; Muller, R.; Trauner, D. Total Syntheses of Cystobactamids and Structural Confirmation of Cystobactamid 919-2. Angew. Chem. Int. Ed. Engl. 2017, 56, 12755–12759. [Google Scholar] [CrossRef] [PubMed]
- Testolin, G.; Cirnski, K.; Rox, K.; Prochnow, H.; Fetz, V.; Grandclaudon, C.; Mollner, T.; Baiyoumy, A.; Ritter, A.; Leitner, C.; et al. Synthetic studies of cystobactamids as antibiotics and bacterial imaging carriers lead to compounds with high in vivo efficacy. Chem. Sci. 2020, 11, 1316–1334. [Google Scholar] [CrossRef] [Green Version]
- Cociancich, S.; Pesic, A.; Petras, D.; Uhlmann, S.; Kretz, J.; Schubert, V.; Vieweg, L.; Duplan, S.; Marguerettaz, M.; Noell, J.; et al. The gyrase inhibitor albicidin consists of p-aminobenzoic acids and cyanoalanine. Nat. Chem. Biol. 2015, 11, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Kaatz, G.W.; Seo, S.M. In vitro activities of oxazolidinone compounds U100592 and U100766 against Staphylococcus aureus and Staphylococcus epidermidis. Antimicrob. Agents Chemother. 1996, 40, 799–801. [Google Scholar] [CrossRef] [Green Version]
- Zurenko, G.E.; Yagi, B.H.; Schaadt, R.D.; Allison, J.W.; Kilburn, J.O.; Glickman, S.E.; Hutchinson, D.K.; Barbachyn, M.R.; Brickner, S.J. In vitro activities of U-100592 and U-100766, novel oxazolidinone antibacterial agents. Antimicrob. Agents Chemother. 1996, 40, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.A.; Di, L.; Kerns, E.H. The effect of plasma protein binding on in vivo efficacy: Misconceptions in drug discovery. Nat. Rev. Drug Discov. 2010, 9, 929–939. [Google Scholar] [CrossRef]
- Waters, N.J.; Jones, R.; Williams, G.; Sohal, B. Validation of a rapid equilibrium dialysis approach for the measurement of plasma protein binding. J. Pharm. Sci. 2008, 97, 4586–4595. [Google Scholar] [CrossRef]
- Stratton, C.W. Dead bugs don’t mutate: Susceptibility issues in the emergence of bacterial resistance. Emerg. Infect. Dis. 2003, 9, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Lourenco, O.; Queiroz, J.A.; Domingues, F.C. Bacteriostatic versus bactericidal activity of ciprofloxacin in Escherichia coli assessed by flow cytometry using a novel far-red dye. J. Antibiot. 2011, 64, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Spangler, S.K.; Jacobs, M.R.; Appelbaum, P.C. Time-kill study of the activity of trovafloxacin compared with ciprofloxacin, sparfloxacin, metronidazole, cefoxitin, piperacillin and piperacillin/tazobactam against six anaerobes. J. Antimicrob. Chemother. 1997, 39 (Suppl. B), 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaddady, P.K.; Lee, R.E.; Meibohm, B. In vitro pharmacokinetic/pharmacodynamic models in anti-infective drug development: Focus on TB. Future Med. Chem. 2010, 2, 1355–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahme, L.G.; Stevens, E.J.; Wolfort, S.F.; Shao, J.; Tompkins, R.G.; Ausubel, F.M. Common virulence factors for bacterial pathogenicity in plants and animals. Science 1995, 268, 1899–1902. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.; Barbour, A.; Sahre, M.; Rand, K.H.; Derendorf, H. PK/PD: New insights for antibacterial and antiviral applications. Curr. Opin. Pharmacol. 2008, 8, 549–556. [Google Scholar] [CrossRef]
- Labreche, M.J.; Graber, C.J.; Nguyen, H.M. Recent Updates on the Role of Pharmacokinetics-pharmacodynamics in Antimicrobial Susceptibility Testing as Applied to Clinical Practice. Clin. Infect. Dis. 2015, 61, 1446–1452. [Google Scholar]
- Craig, W.A. Does the dose matter? Clin. Infect. Dis. 2001, 33 (Suppl. 3), S233–S237. [Google Scholar] [CrossRef]
- Dasgupta, A. Usefulness of monitoring free (unbound) concentrations of therapeutic drugs in patient management. Clin. Chim. Acta 2007, 377, 1–13. [Google Scholar] [CrossRef]
- LeBel, M. Ciprofloxacin: Chemistry, mechanism of action, resistance, antimicrobial spectrum, pharmacokinetics, clinical trials, and adverse reactions. Pharmacotherapy 1988, 8, 3–33. [Google Scholar] [CrossRef]
- Rybak, M.J. The pharmacokinetic and pharmacodynamic properties of vancomycin. Clin. Infect. Dis. 2006, 42 (Suppl. 1), S35–S39. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, Y.; Holford, N.H.G.; Kasai, H.; Ogami, C.; Heo, Y.A.; Higashi, Y.; Mizoguchi, A.; To, H.; Yamamoto, Y. Population pharmacokinetics and pharmacodynamics of linezolid-induced thrombocytopenia in hospitalized patients. Br. J. Clin. Pharmacol. 2017, 83, 1758–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulldemolins, M.; Roberts, J.A.; Rello, J.; Paterson, D.L.; Lipman, J. The effects of hypoalbuminaemia on optimizing antibacterial dosing in critically ill patients. Clin. Pharmacokinet. 2011, 50, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Lepak, A.J.; Zhao, M.; Andes, D.R. Comparative Pharmacodynamics of Telavancin and Vancomycin in the Neutropenic Murine Thigh and Lung Infection Models against Staphylococcus aureus. Antimicrob. Agents Chemother. 2017, 61, e00281-17. [Google Scholar] [CrossRef] [Green Version]
- Andes, D.; van Ogtrop, M.L.; Peng, J.; Craig, W.A. In vivo pharmacodynamics of a new oxazolidinone (linezolid). Antimicrob. Agents Chemother. 2002, 46, 3484–3489. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. Twenty-Third Informational Supplement M100-S23; CLSI: Wayne, PA, USA, 2013. [Google Scholar]
- National Committee for Clinical Laboratory Standards. Methods for Determining Bactericidal Activity of Antimicrobial Agents: Approved Guideline M26-A; NCCLS: Wayne, PA, USA, 1999. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism (No. of Strains) Antimicrobial Agent | MIC (µg/mL) | Organism (No. of Strains) Antimicrobial Agent | MIC (µg/mL) | ||
|---|---|---|---|---|---|
| MIC50 | MIC90 | MIC50 | MIC90 | ||
| MSSA (26) | S. pneumoniae (27) | ||||
| Coralmycin A | 0.5 | 1 | Coralmycin A | 0.25 | 1 |
| DH-coralmycin A | 1 | 2 | DH-coralmycin A | 0.5 | 4 |
| Oxacillin | 0.5 | 1 | Meropenem | 0.5 | 4 |
| Meropenem | 0.125 | 0.25 | Levofloxacin | 0.5 | 16 |
| Levofloxacin | 0.25 | 0.5 | Gentamicin | 32 | >32 |
| Gentamicin | 1 | >32 | Vancomycin | 0.25 | 0.5 |
| Vancomycin | 1 | 2 | Cefoxitin | >32 | >32 |
| Cefoxitin | 4 | 4 | Ceftazidime | >32 | >32 |
| Ceftazidime | 8 | 16 | Daptomycin | 2 | 8 |
| Daptomycin | 2 | 4 | Linezolid | 2 | 4 |
| Linezolid | 2 | 2 | |||
| Vancomycin-sensitive E. faecalis (32) | |||||
| MRSA (42) | Coralmycin A | 1 | 2 | ||
| Coralmycin A | 1 | 1 | DH-coralmycin A | 2 | 4 |
| DH-coralmycin A | 4 | 8 | Meropenem | 4 | 16 |
| Oxacillin | >32 | >32 | Levofloxacin | 2 | 32 |
| Levofloxacin | 16 | >32 | Gentamicin | >32 | >32 |
| Gentamicin | 32 | >32 | Vancomycin | 2 | 2 |
| Vancomycin | 1 | 2 | Cefoxitin | >32 | >32 |
| Ceftazidime | >32 | >32 | Ceftazidime | >32 | >32 |
| Daptomycin | 2 | 4 | Daptomycin | 4 | 8 |
| Linezolid | 2 | 4 | Linezolid | 2 | 4 |
| CNS (31) | Vancomycin-sensitive E. faecium (32) | ||||
| Coralmycin A | 0.25 | 1 | Coralmycin A | 0.25 | 0.5 |
| DH-coralmycin A | 1 | 16 | DH-coralmycin A | 1 | 4 |
| Levofloxacin | 8 | >32 | Meropenem | >32 | >32 |
| Gentamicin | >32 | >32 | Levofloxacin | 16 | 32 |
| Vancomycin | 2 | 4 | Gentamicin | >32 | >32 |
| Ceftazidime | 32 | >32 | Vancomycin | 1 | 2 |
| Daptomycin | 2 | 4 | Cefoxitin | >32 | >32 |
| Linezolid | 2 | 4 | Ceftazidime | >32 | >32 |
| Daptomycin | 4 | 8 | |||
| VRE (46) | Linezolid | 2 | 4 | ||
| Coralmycin A | 0.5 | 1 | |||
| DH-coralmycin A | 4 | 16 | |||
| Meropenem | >32 | >32 | |||
| Levofloxacin | >32 | >32 | |||
| Gentamicin | >32 | >32 | |||
| Vancomycin | >32 | >32 | |||
| Cefoxitin | >32 | >32 | |||
| Ceftazidime | >32 | >32 | |||
| Daptomycin | 4 | 8 | |||
| Linezolid | 2 | 4 | |||
| Parameter | Coralmycin A | DH-Coralmycin A | ||
|---|---|---|---|---|
| 2 mg/kg i.v. | 20 mg/kg s.c. | 2 mg/kg i.v. | 20 mg/kg s.c. | |
| Tmax (h) | 0.08 ± 0.00 | 1.67 ± 0.58 | 0.08 ± 0.00 | 0.67 ± 0.29 |
| Cmax (μg/mL) | 1.12 ± 0.68 | 0.70 ± 0.12 | 1.63 ± 0.16 | 0.19 ± 0.03 |
| T1/2 (h) | 1.32 ± 0.71 | 3.31 ± 0.15 | 2.16 ± 1.08 | 7.01 ± 1.22 |
| AUClast (μg·h/mL) | 0.52 ± 0.26 | 3.16 ± 1.05 | 0.66 ± 0.06 | 0.78 ± 0.11 |
| AUCinf (μg·h/mL) | 0.52 ± 0.26 | 3.19 ± 1.06 | 0.67 ± 0.06 | 0.87 ± 0.12 |
| CL (L/h/kg) | 4.43 ± 1.73 | NA | 2.99 ± 0.27 | NA |
| Vdss (L/kg) | 8.53 ± 5.68 | NA | 1.46 ± 0.40 | NA |
| F (%) | NA | 61.3 | NA | 11.7 |
| 0.2 and 1 μM | 5 μM | 5 μM | ||
|---|---|---|---|---|
| Coralmycin A or DH-Coralmycin A | Coralmycin A | DH-Coralmycin A | Quinidine b | |
| Mouse | ND a | 98.7 ± 0.48 | 92.5 ± 2.24 | 77.1 ± 1.44 |
| Rat | ND | 99.8 ± 0.02 | 98.5 ± 0.90 | 73.8 ± 4.53 |
| Human | ND | 99.7 ± 0.09 | 97.7 ± 0.64 | 83.5 ± 1.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.-Y.; Kim, B.-M.; Kim, Y.-R.; Yang, T.; Ahn, S.; Yong, D.; Kwak, J.-H.; Kim, W.-G. Antibacterial Activity against Clinical Isolates and In Vivo Efficacy of Coralmycins. Antibiotics 2022, 11, 902. https://doi.org/10.3390/antibiotics11070902
Choi H-Y, Kim B-M, Kim Y-R, Yang T, Ahn S, Yong D, Kwak J-H, Kim W-G. Antibacterial Activity against Clinical Isolates and In Vivo Efficacy of Coralmycins. Antibiotics. 2022; 11(7):902. https://doi.org/10.3390/antibiotics11070902
Chicago/Turabian StyleChoi, Ha-Young, Bo-Min Kim, Young-Rok Kim, Taehui Yang, Sunjoo Ahn, Dongeun Yong, Jin-Hwan Kwak, and Won-Gon Kim. 2022. "Antibacterial Activity against Clinical Isolates and In Vivo Efficacy of Coralmycins" Antibiotics 11, no. 7: 902. https://doi.org/10.3390/antibiotics11070902