
Understanding microRNAs in the Context of Infection to Find New Treatments against Human Bacterial Pathogens

 ,
,  ,
, 
 ,
,  , and
, and
Abstract
:1. Introduction
2. Human miRNAs and Pathogen Infections
2.1. Role of miRNAs in the Regulation of Host Inflammatory Responses during Bacterial Infection
2.1.1. Mycobacterium tuberculosis
2.1.2. Francisella tularensis
2.1.3. Vibrio cholerae
2.1.4. Staphylococcus aureus
2.1.5. Helicobacter pylori
2.1.6. Chlamydia trachomatis
2.1.7. Broad-Spectrum miRNAs
2.2. Role of miRNAs in the Control of the Infected Host Cell
2.2.1. Mycobacterium tuberculosis
2.2.2. Adherent–Invasive Escherichia coli
2.2.3. Legionella pneumophila
2.2.4. Chlamydia trachomatis
2.2.5. Shigella flexneri and Salmonella enterica Serovar Typhimurium
2.2.6. Burkholderia pseudomallei
2.2.7. Listeria monocytogenes
3. Novel Antimicrobial Treatments Based on miRNA-Based Technology
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Kim, T.S.; Basu, J.; Jo, E.K. MicroRNA in innate immunity and autophagy during mycobacterial infection. Cell. Microbiol. 2017, 19, e12687. [Google Scholar] [CrossRef] [Green Version]
- Eulalio, A.; Schulte, L.N.; Voge, J. The mammalian microRNA response to bacterial infections. RNA Biol. 2012, 9, 742–750. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef] [Green Version]
- de Sousa, M.C.; Gjorgjieva, M.; Dolicka, D.; Sobolewski, C.; Foti, M. Deciphering miRNAs’ action through miRNA editing. Int. J. Mol. Sci. 2019, 20, 6249. [Google Scholar] [CrossRef] [Green Version]
- Orang, A.V.; Safaralizadeh, R.; Kazemzadeh-Bavili, M. Mechanisms of miRNA-mediated gene regulation from common downregulation to mRNA-specific upregulation. Int. J. Genom. 2014, 2014, 970607. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Rao, D.D.; Senzer, N.; Nemunaitis, J. RNA interference and cancer therapy. Pharm. Res. 2011, 28, 2983–2995. [Google Scholar] [CrossRef]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA regulation of RNA virus replication and pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Asl, E.R.; Amini, M.; Najafi, S.; Mansoori, B.; Mokhtarzadeh, A.; Mohammadi, A.; Lotfinejad, P.; Bagheri, M.; Shirjang, S.; Lotfi, Z.; et al. Interplay between MAPK/ERK signaling pathway and MicroRNAs: A crucial mechanism regulating cancer cell metabolism and tumor progression. Life Sci. 2021, 119499. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, C.; Cruz, A.R.; Rodrigues Lopes, I.; Maudet, C.; Sunkavalli, U.; Silva, R.J.; Sharan, M.; Lisowski, C.; Zaldívar-López, S.; Garrido, J.J.; et al. Functional screenings reveal different requirements for host microRNAs in Salmonella and Shigella infection. Nat. Microbiol. 2020, 5, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Londina, E.; Lohera, P.; Telonis, A.G.; Quann, K.; Clark, P.; Jinga, Y.; Hatzimichael, E.; Kirino, Y.; Honda, S.; Lally, M.; et al. Analysis of 13 cell types reveals evidence for the expression of numerous novel primate- and tissue-specific microRNAs. Proc. Natl. Acad. Sci. USA 2015, 112, E1106–E1115. [Google Scholar] [CrossRef] [Green Version]
- Cullen, B.R. Viruses and microRNAs: RISCy interactions with serious consequences. Genes Dev. 2011, 25, 1881–1894. [Google Scholar] [CrossRef] [Green Version]
- Dölken, L.; Haas, J. Small noncoding RNA: Novel targets for antiviral therapy. Future Microbiol. 2008, 3, 585–588. [Google Scholar] [CrossRef]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Kumar, A.; Ingle, H.; Kumar, H. The interplay between viral-derived miRNAs and host immunity during infection. Front. Immunol. 2020, 10, 3079. [Google Scholar] [CrossRef] [Green Version]
- Hum, C.; Loiselle, J.; Ahmed, N.; Shaw, T.A.; Toudic, C.; Pezacki, J.P. MicroRNA mimics or inhibitors as antiviral therapeutic approaches against COVID-19. Drugs 2021, 81, 517–531. [Google Scholar] [CrossRef]
- Yee, P.; Poh, C. Development of novel miRNA-based vaccines and antivirals against Enterovirus 71. Curr. Pharm. Des. 2016, 22. [Google Scholar] [CrossRef]
- Singh, P.K.; Singh, A.V.; Chauhan, D.S. Current understanding on micro RNAs and its regulation in response to Mycobacterial infections. J. Biomed. Sci. 2013, 20, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D.G. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 2016, 312, 436–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Garnica, O.; Dhandayuthapani, S. Modulation of host miRNAs by intracellular bacterial pathogens. Front. Cell. Infect. Microbiol. 2016, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Silwal, P.; Kim, Y.S.; Basu, J.; Jo, E.K. The roles of microRNAs in regulation of autophagy during bacterial infection. Semin. Cell Dev. Biol. 2020, 101, 51–58. [Google Scholar] [CrossRef]
- Zhou, X.; Li, X.; Wu, M. miRNAs reshape immunity and inflammatory responses in bacterial infection. Signal Transduct. Target. Ther. 2018, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Chen, H.; Hu, Y.; Li, Q.; Hu, Z.; Ma, T.; Mao, X. Burkholderia pseudomallei-derived miR-3473 enhances NF-κB via targeting TRAF3 and is associated with different inflammatory responses compared to Burkholderia thailandensis in murine macrophages. BMC Microbiol. 2016, 16, 283. [Google Scholar] [CrossRef] [Green Version]
- Britton, C.; Winter, A.D.; Gillan, V.; Devaney, E. MicroRNAs of parasitic helminths—Identification, characterization and potential as drug targets. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [Green Version]
- Larabi, A.; Dalmasso, G.; Delmas, J.; Barnich, N.; Nguyen, H.T.T. Exosomes transfer miRNAs from cell-to-cell to inhibit autophagy during infection with Crohn’s disease-associated Adherent–Invasive E. coli. Gut Microbes 2020, 11, 1677–1694. [Google Scholar] [CrossRef]
- Hu, Z.Q.; Rao, C.L.; Tang, M.L.; Zhang, Y.; Lu, X.X.; Chen, J.G.; Mao, C.; Deng, L.; Li, Q.; Mao, X.H. Rab32 GTpase, as a direct target of miR-30b/c, controls the intracellular survival of Burkholderia pseudomallei by regulating phagosome maturation. PLoS Pathog. 2019, 15, e1007879. [Google Scholar] [CrossRef]
- Chowdhury, S.R.; Reimer, A.; Sharan, M.; Kozjak-Pavlovic, V.; Eulalio, A.; Prusty, B.K.; Fraunholz, M.; Karunakaran, K.; Rudel, T. Chlamydia preserves the mitochondrial network necessary for replication via microRNA-dependent inhibition of fission. J. Cell Biol. 2017, 216, 1071–1089. [Google Scholar] [CrossRef] [PubMed]
- Igietseme, J.U.; Omosun, Y.; Stuchlik, O.; Reed, M.S.; Partin, J.; He, Q.; Joseph, K.; Ellerson, D.; Bollweg, B.; George, Z.; et al. Role of epithelial-mesenchyme transition in Chlamydia pathogenesis. PLoS ONE 2015, 10, e0145198. [Google Scholar] [CrossRef] [PubMed]
- Derrick, T.; Last, A.R.; Burr, S.E.; Roberts, C.H.; Nabicassa, M.; Cassama, E.; Bailey, R.L.; Mabey, D.C.W.; Burton, M.J.; Holland, M.J. Inverse relationship between microRNA-155 and -184 expression with increasing conjunctival inflammation during ocular Chlamydia trachomatis infection. BMC Infect. Dis. 2016, 16, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandyopadhyay, S.; Long, M.E.; Allen, L.A.H. Differential expression of microRNAs in Francisella tularensis-infected human macrophages: miR-155-dependent downregulation of MyD88 inhibits the inflammatory response. PLoS ONE 2014, 9, e109525. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Liu, S.F.; Dong, K.; Zhang, G.C.; Huang, J.; Wang, Z.H.; Wang, T.J. Exosome-transmitted miR-25 induced by H. pylori promotes vascular endothelial cell injury by targeting KLF2. Front. Cell. Infect. Microbiol. 2019, 9, 366. [Google Scholar] [CrossRef]
- Herkt, C.E.; Caffrey, B.E.; Surmann, K.; Blankenburg, S.; Salazar, G.; Jung, A.L.; Herbel, S.M.; Hoffmann, K.; Schulte, L.N.; Chen, W.; et al. A microRNA network controls Legionella pneumophila replication in human macrophages via LGALS8 and MX1. MBio 2020, 11, e03155-19. [Google Scholar] [CrossRef] [Green Version]
- Johnston, D.G.W.; Kearney, J.; Zasłona, Z.; Williams, M.A.; O’Neill, L.A.J.; Corr, S.C. MicroRNA-21 limits uptake of Listeria monocytogenes by macrophages to reduce the intracellular niche and control infection. Front. Cell. Infect. Microbiol. 2017, 7, 201. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, J.; Wang, L.; Zheng, Z.; Ran, H.; Liu, F.; Li, F.; Tang, X.; Zhang, J.; Ni, Q.; et al. MiR-26a targets EphA2 to resist intracellular Listeria monocytogenes in macrophages. Mol. Immunol. 2020, 128, 69–78. [Google Scholar] [CrossRef]
- Ma, F.; Xu, S.; Liu, X.; Zhang, Q.; Xu, X.; Liu, M.; Hua, M.; Li, N.; Yao, H.; Cao, X. The microRNA miR-29 controls innate and adaptive immune responses to intracellular bacterial infection by targeting interferon-γ. Nat. Immunol. 2011, 12, 861–869. [Google Scholar] [CrossRef]
- Guo, L.; Zhou, L.; Gao, Q.; Zhang, A.; Wei, J.; Hong, D.; Chu, Y.; Duan, X.; Zhang, Y.; Xu, G. MicroRNA-144-3p inhibits autophagy activation and enhances Bacillus Calmette-Guérin infection by targeting ATG4a in RAW264.7 macrophage cells. PLoS ONE 2017, 12, e0179772. [Google Scholar] [CrossRef]
- Kumar, R.; Sahu, S.K.; Kumar, M.; Jana, K.; Gupta, P.; Gupta, U.D.; Kundu, M.; Basu, J. MicroRNA 17-5p regulates autophagy in Mycobacterium tuberculosis-infected macrophages by targeting Mcl-1 and STAT3. Cell. Microbiol. 2016, 18, 679–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.; Chen, H.; Yang, Y.; Fu, Y.; Yi, Z. miR-18a promotes Mycobacterial survival in macrophages via inhibiting autophagy by downregulation of ATM. J. Cell. Mol. Med. 2020, 24, 2004–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Li, Z.; Cui, K.; Gao, Y.; Zhang, B.; Niu, J.; Wang, Y. MicroRNA-20a-3p regulates the host immune response to facilitate the Mycobacterium tuberculosis infection by targeting IKKβ/NF-κB pathway. Int. Immunopharmacol. 2021, 91, 107286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yi, Z.; Fu, Y. Downregulation of miR-20b-5p facilitates Mycobacterium tuberculosis survival in RAW 264.7 macrophages via attenuating the cell apoptosis by Mcl-1 upregulation. J. Cell. Biochem. 2019, 120, 5889–5896. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chen, J.; Wang, P.; Li, H.; Zhou, Y.; Liu, H.; Liu, Z.; Zheng, R.; Wang, L.; Yang, H.; et al. MicroRNA-27a controls the intracellular survival of Mycobacterium tuberculosis by regulating calcium-associated autophagy. Nat. Commun. 2018, 9, 4295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouimet, M.; Koster, S.; Sakowski, E.; Ramkhelawon, B.; Van Solingen, C.; Oldebeken, S.; Karunakaran, D.; Portal-celhay, C.; Sheedy, F.J.; Ray, T.D.; et al. Mycobacterium tuberculosis induces the miR-33 locus to reprogram autophagy and host lipid metabolism. Nat. Immunol. 2016, 17, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Kaul, V.; Mehra, A.; Chatterjee, S.; Tousif, S.; Dwivedi, V.P.; Suar, M.; Van Kaer, L.; Bishai, W.R.; Das, G. Mycobacterium tuberculosis controls MicroRNA-99b (miR-99b) expression in infected murine dendritic cells to modulate host immunity. J. Biol. Chem. 2013, 288, 5056–5061. [Google Scholar] [CrossRef] [Green Version]
- Zuo, X.; Wang, L.; Bao, Y.; Sun, J. The ESX-1 virulence factors downregulate miR-147-3p in Mycobacterium marinum-infected macrophages. Infect. Immun. 2020, 88, e00088-20. [Google Scholar] [CrossRef]
- Wu, H.; Bao, Y.; Wang, L.; Li, X.; Sun, J. Mycobacterium marinum downregulates miR-148a in macrophages in an EsxA-dependent manner. Int. Immunopharmacol. 2020, 73, 41–48. [Google Scholar] [CrossRef]
- Rothchild, A.C.; Sissons, J.R.; Shafiani, S.; Plaisier, C.; Min, D.; Mai, D.; Gilchrist, M.; Peschon, J.; Larson, R.P.; Bergthaler, A.; et al. MiR-155-regulated molecular network orchestrates cell fate in the innate and adaptive immune response to Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2016, 113, E6172–E6181. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yang, K.; Zhou, L.; Wu, M.; Wu, Y.; Zhu, M.; Lai, X.M.; Chen, T.; Feng, L.; Li, M.; et al. MicroRNA-155 promotes autophagy to eliminate intracellular mycobacteria by targeting Rheb. PLoS Pathog. 2013, 9, e1003697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.; Mao, G.; Xie, K.; Wu, D.; Wang, W. MiR-1178 regulates mycobacterial survival and inflammatory responses in Mycobacterium tuberculosis-infected macrophages partly via TLR4. J. Cell. Biochem. 2018, 119, 7449–7457. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Qu, Y.; Yang, S.; Zhao, Y.; Xu, G. Novel miR-1958 promotes Mycobacterium tuberculosis survival in RAW264.7 cells by inhibiting autophagy via Atg5. J. Microbiol. Biotechnol. 2019, 29, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Maudet, C.; Mano, M.; Sunkavalli, U.; Sharan, M.; Giacca, M.; Förstner, K.U.; Eulalio, A. Functional high-throughput screening identifies the miR-15 microRNA family as cellular restriction factors for Salmonella infection. Nat. Commun. 2014, 5, 4718. [Google Scholar] [CrossRef]
- Hoeke, L.; Sharbati, J.; Pawar, K.; Keller, A.; Einspanier, R.; Sharbati, S. Intestinal Salmonella Typhimurium infection leads to miR-29a induced Caveolin 2 regulation. PLoS ONE 2013, 8, e67300. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Mao, Y.; Kang, Y.; He, L.; Zhu, B.; Zhang, W.; Lu, Y.; Wu, Q.; Xu, D.; Shi, L. MicroRNA-127 promotes anti-microbial host defense through restricting A20-mediated de-ubiquitination of STAT3. iScience 2020, 23, 100763. [Google Scholar] [CrossRef] [Green Version]
- Bitar, A.; Aung, K.M.; Wai, S.N.; Hammarström, M.L. Vibrio cholerae derived outer membrane vesicles modulate the inflammatory response of human intestinal epithelial cells by inducing microRNA-146a. Sci. Rep. 2019, 9, 7212. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Li, Y.; Zeng, J.; Wu, X.; Liu, X.; Wang, Y. Mycobacterium bovis BCG triggered MyD88 induces miR-124 feedback negatively regulates immune response in alveolar epithelial cells. PLoS ONE 2014, 9, e92419. [Google Scholar] [CrossRef]
- Ma, C.; Li, Y.; Li, M.; Deng, G.; Wu, X.; Zeng, J.; Hao, X.; Wang, X.; Liu, J.; Cho, W.C.S.; et al. MicroRNA-124 negatively regulates TLR signaling in alveolar macrophages in response to mycobacterial infection. Mol. Immunol. 2014, 62, 150–158. [Google Scholar] [CrossRef]
- Zhou, X.; Li, X.; Ye, Y.; Zhao, K.; Zhuang, Y.; Li, Y.; Wei, Y.; Wu, M. MicroRNA-302b augments host defense to bacteria by regulating inflammatory responses via feedback to TLR/IRAK4 circuits. Nat. Commun. 2014, 5, 3619. [Google Scholar] [CrossRef]
- Wright, K.; de Silva, K.; Plain, K.M.; Purdie, A.C.; Blair, T.A.; Duggin, I.G.; Britton, W.J.; Oehlers, S.H. Mycobacterial infection-induced miR-206 inhibits protective neutrophil recruitment via the CXCL12/CXCR4 signalling axis. PLoS Pathog. 2021, 17, e1009186. [Google Scholar] [CrossRef] [PubMed]
- Cremer, T.J.; Ravneberg, D.H.; Clay, C.D.; Piper-Hunter, M.G.; Marsh, C.B.; Elton, T.S.; Gunn, J.S.; Amer, A.; Kanneganti, T.D.; Schlesinger, L.S.; et al. MiR-155 induction by F. novicida but not the virulent F. tularensis results in SHIP downregulation and enhanced pro-inflammatory cytokine response. PLoS ONE 2009, 4, e8508. [Google Scholar] [CrossRef] [PubMed]
- Podsiad, A.; Standiford, T.J.; Ballinger, M.N.; Eakin, R.; Park, P.; Kunkel, S.L.; Moore, B.B.; Bhan, U. MicroRNA-155 regulates host immune response to postviral bacterial pneumonia via IL-23/IL-17 pathway. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2016, 310, L465–L475. [Google Scholar] [CrossRef] [PubMed]
- Săsăran, M.O.; Meliț, L.E.; Dobru, E.D. MicroRNA modulation of host immune response and inflammation triggered by Helicobacter pylori. Int. J. Mol. Sci. 2021, 22, 1406. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Xiao, B.; Liu, Z.; Li, N.; Zhu, E.D.; Li, B.S.; Xie, Q.H.; Zhuang, Y.; Zou, Q.M.; Mao, X.H. Identification of MyD88 as a novel target of miR-155, involved in negative regulation of Helicobacter pylori-induced inflammation. FEBS Lett. 2010, 584, 1481–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceppi, M.; Pereira, A.M.; Dunand-Sauthier, I.; Barras, E.; Reith, W.; Santos, M.A.; Pierre, P. MicroRNA-155 modulates the interleukin-1 signaling pathway in activated human monocyte-derived dendritic cells. Proc. Natl. Acad. Sci. USA 2009, 106, 2735–2740. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Baek, H.; Park, J.S.; Kim, S.; Kyung, C.; Baik, S.J.; Lee, B.K.; Kim, J.H.; Ahn, C.W.; Kim, K.R.; et al. Current Helicobacter pylori infection is significantly associated with subclinical coronary atherosclerosis in healthy subjects: Across-sectional study. PLoS ONE 2018, 13, e0193646. [Google Scholar] [CrossRef]
- Li, B.S.; Zhao, Y.L.; Guo, G.; Li, W.; Zhu, E.D.; Luo, X.; Mao, X.H.; Zou, Q.M.; Yu, P.W.; Zuo, Q.F.; et al. Plasma microRNAs, miR-223, miR-21 and miR-218, as novel potential biomarkers for gastric cancer detection. PLoS ONE 2012, 7, e41629. [Google Scholar] [CrossRef] [Green Version]
- Eledge, M.R.; Yeruva, L. Host and pathogen interface: microRNAs are modulators of disease outcome. Microbes Infect. 2018, 20, 410–415. [Google Scholar] [CrossRef]
- Derrick, T.; Roberts, C.H.; Rajasekhar, M.; Burr, S.E.; Joof, H.; Makalo, P.; Bailey, R.L.; Mabey, D.C.W.; Burton, M.J.; Holland, M.J. Conjunctival MicroRNA Expression in Inflammatory Trachomatous Scarring. PLoS Negl. Trop. Dis. 2013, 7. [Google Scholar] [CrossRef]
- Yeruva, L.; Myers, G.S.A.; Spencer, N.; Creasy, H.H.; Adams, N.E.; Maurelli, A.T.; McChesney, G.R.; Cleves, M.A.; Ravel, J.; Bowlin, A.; et al. Early MicroRNA expression profile as a prognostic biomarker for the development of pelvic inflammatory disease in a mouse model of chlamydial genital infection. MBio 2014, 5, e01241-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, R.; Krishnamoorthy, P.; Kumar, H. MicroRNA-30e-5p regulates SOCS1 and SOCS3 during bacterial infection. Front. Cell. Infect. Microbiol. 2021, 10, 887. [Google Scholar] [CrossRef] [PubMed]
- Alice, A.F.; Kramer, G.; Bambina, S.; Baird, J.R.; Bahjat, K.S.; Gough, M.J.; Crittenden, M.R. Amplifying IFN-γ signaling in dendritic cells by CD11c-specific loss of SOCS1 increases innate immunity to infection while decreasing adaptive immunity. J. Immunol. 2018, 200, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Medina, M.; Aldeguer, X.; Lopez-Siles, M.; González-Huix, F.; López-Oliu, C.; Dahbi, G.; Bianco, J.E.; Blanco, J.; Garcia-Gil, L.J.; Darfeuille-Michaud, A. Molecular diversity of Escherichia coli in the human gut: New ecological evidence supporting the role of Adherent–Invasive E. coli (AIEC) in Crohn’s disease. Inflamm. Bowel Dis. 2009, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Spier, A.; Stavru, F.; Cossart, P. Interaction between intracellular bacterial pathogens and host cell mitochondria. Microbiol. Spectr. 2019, 7, BAI-0016-2019. [Google Scholar] [CrossRef] [PubMed]
- Sunkavalli, U.; Aguilar, C.; Silva, R.J.; Sharan, M.; Cruz, A.R.; Tawk, C.; Maudet, C.; Mano, M.; Eulalio, A. Analysis of host microRNA function uncovers a role for miR-29b-2-5p in Shigella capture by filopodia. PLoS Pathog. 2017, 13, e1006327. [Google Scholar] [CrossRef]
- Aguilar, C.; Costa, S.; Maudet, C.; Vivek-Ananth, R.P.; Zaldívar-López, S.; Garrido, J.J.; Samal, A.; Mano, M.; Eulalio, A. Reprogramming of microRNA expression via E2F1 downregulation promotes Salmonella infection both in infected and bystander cells. Nat. Commun. 2021, 12, 3392. [Google Scholar] [CrossRef]
- Du, C.T.; Gao, W.; Ma, K.; Yu, S.X.; Li, N.; Yan, S.Q.; Zhou, F.H.; Liu, Z.Z.; Chen, W.; Lei, L.C.; et al. MicroRNA-146a deficiency protects against Listeria monocytogenes infection by modulating the gut microbiota. Int. J. Mol. Sci. 2018, 19, 993. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Brandt, S.; Medeiros, A.; Wang, S.; Wu, H.; Dent, A.; Serezani, C.H. MicroRNA 21 Is a homeostatic regulator of macrophage polarization and prevents prostaglandin e2 -mediated M2 generation. PLoS ONE 2015, 10, e0115855. [Google Scholar] [CrossRef]
- Subbarayal, P.; Karunakaran, K.; Winkler, A.C.; Rother, M.; Gonzalez, E.; Meyer, T.F.; Rudel, T. EphrinA2 Receptor (EphA2) is an invasion and intracellular signaling receptor for Chlamydia trachomatis. PLoS Pathog. 2015, 11, e1004846. [Google Scholar] [CrossRef]
- Bravo-santano, N.; Stölting, H.; Cooper, F.; Bileckaja, N.; Majstorovic, A.; Ihle, N.; Mateos, L.M.; Calle, Y.; Behrends, V.; Letek, M. Host-directed kinase inhibitors act as novel therapies against intracellular Staphylococcus aureus. Sci. Rep. 2019, 9, 4876. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, M.; Dorhoi, A.; Kaufmann, S.H.E. Host-directed therapy of tuberculosis: What is in it for microRNA? Expert Opin. Ther. Targets 2014, 18, 491–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, D.K.W.; Chow, M.Y.T.; Casettari, L.; Gonzalez-Juarrero, M.; Lam, J.K.W. Potential and development of inhaled RNAi therapeutics for the treatment of pulmonary tuberculosis. Adv. Drug Deliv. Rev. 2016, 102, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, C.C.; Choy, K.W.; Du, Q.; Chen, J.; Wang, Q.; Li, L.; Chung, T.K.H.; Tang, T. Therapeutic potentials of gene silencing by RNA interference: Principles, challenges, and new strategies. Gene 2014, 538, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Pecot, C.V.; Calin, G.A.; Coleman, R.L.; Lopez-Berestein, G.; Sood, A.K. RNA interference in the clinic: Challenges and future directions. Nat. Rev. Cancer 2011, 11, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Lam, J.K.W.; Chow, M.Y.T.; Zhang, Y.; Leung, S.W.S. siRNA versus miRNA as therapeutics for gene silencing. Mol. Ther.-Nucleic Acids 2015, 4, e252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas-taraco, A.G.; Higgins, D.M.; Sánchez-campillo, J.; Lee, E.J.; Orme, I.M.; González-juarrero, M. Local pulmonary immunotherapy with siRNA targeting TGFβ1 enhances antimicrobial capacity in Mycobacterium tuberculosis infected mice. Tuberculosis 2011, 91, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Drury, R.E.; O’Connor, D.; Pollard, A.J. The clinical application of MicroRNAs in infectious disease. Front. Immunol. 2017, 8, 1182. [Google Scholar] [CrossRef] [PubMed]
- Jamalkhah, M.; Asaadi, Y.; Azangou-Khyavy, M.; Khanali, J.; Soleimani, M.; Kiani, J.; Arefian, E. MSC-derived exosomes carrying a cocktail of exogenous interfering RNAs an unprecedented therapy in era of COVID-19 outbreak. J. Transl. Med. 2021, 19, 164. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; Rebhan, M.A.E.; Crivelli, S.E.M.; Denzler, R.; Stoffel, M.; Hall, J. Miravirsen (SPC3649) can inhibit the biogenesis of miR-122. Nucleic Acids Res. 2014, 42, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Janssen, H.L.A.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullis, P.R.; Hope, M.J. Lipid nanoparticle systems for enabling gene therapies. Mol. Ther. 2017, 25, 1467–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, P.; Coban, O.; Snead, N.M.; Trebley, J.; Hoeprich, S.; Guo, S.; Shu, Y. Engineering RNA for targeted siRNA delivery and medical application. Adv. Drug Deliv. Rev. 2010, 62, 650–666. [Google Scholar] [CrossRef]
- Akuma, P.; Okagu, O.D.; Udenigwe, C.C. Naturally Occurring Exosome Vesicles as Potential Delivery Vehicle for Bioactive Compounds. Front. Sustain. Food Syst. 2019, 3, 23. [Google Scholar] [CrossRef]
- Mathiyalagan, P.; Sahoo, S. Exosomes-based gene therapy for MicroRNA delivery. Methods Mol. Biol. 2017, 1521, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Qiao, L.; Hu, S.; Huang, K.; Su, T.; Li, Z.; Vandergriff, A.; Cores, J. Theranostics tumor cell-derived exosomes home to their cells of origin and can be used as Trojan horses to deliver cancer drugs. Theranostics 2020, 10, 3474–3487. [Google Scholar] [CrossRef]
- Alvarez-erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Wang, J.; Dong, Y.; Li, Y.; Li, W.; Cheng, K.; Qian, Y.; Xu, G. Designer exosomes for active targeted chemo-photothermal synergistic tumor therapy. Adv. Func. Mat. 2018, 28, 1707360. [Google Scholar] [CrossRef]
- Matsuki, Y.; Yanagawa, T.; Sumiyoshi, H.; Yasuda, J. Modification of exosomes with carbonate apatite and a glycan polymer improves transduction ef fi ciency and target cell selectivity. Biochem. Biophys. Res. Commun. 2021, 583, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Schorey, J.S.; Harding, C.V.; Schorey, J.S.; Harding, C. V Extracellular vesicles and infectious diseases : New complexity Extracellular vesicles and infectious diseases : New complexity to an old story. J. Clin. Investig. 2016, 126, 1181–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, H.M.; Ali, A.; Fooladi, I.; Nourani, M.R.; Ghanezadeh, F. The role of exosomes in infectious diseases. Inflamm. Allergy-Drug Targets 2013, 12, 29–37. [Google Scholar] [CrossRef]
- Jones, L.B.; Bell, C.R.; Bibb, K.E.; Gu, L.; Coats, M.T.; Matthews, Q.L. Pathogens and their effect on exosome biogenesis and composition. Biomedicines 2018, 6, 79. [Google Scholar] [CrossRef] [Green Version]
- Neupane, K.R.; Mccorkle, J.R.; Kopper, T.J.; Lakes, J.E.; Aryal, S.P.; Abdullah, M.; Snell, A.A.; Gensel, J.C.; Kolesar, J.; Richards, C.I. Macrophage-engineered vesicles for therapeutic delivery and bidirectional reprogramming of immune cell polarization. ACS Omega 2021, 6, 3847–3857. [Google Scholar] [CrossRef]
- Alipoor, S.D.; Mortaz, E.; Tabarsi, P.; Farnia, P.; Mirsaeidi, M.; Garssen, J.; Movassaghi, M.; Adcock, I.M. Bovis Bacillus Calmette-Guerin (BCG) infection induces exosomal miRNA release by human macrophages. J. Transl. Med. 2017, 15, 105. [Google Scholar] [CrossRef] [Green Version]
- Mosquera-Heredia, M.I.; Morales, L.C.; Vidal, O.M.; Barceló, E.; Silvera-Redondo, C.; Vélez, J.I.; Garavito-Galofre, P. Exosomes: Potential disease biomarkers and new therapeutic targets. Biomedicines 2021, 9, 1061. [Google Scholar] [CrossRef]
- Barile, L.; Vassalli, G. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 2017, 174, 63–78. [Google Scholar] [CrossRef] [Green Version]


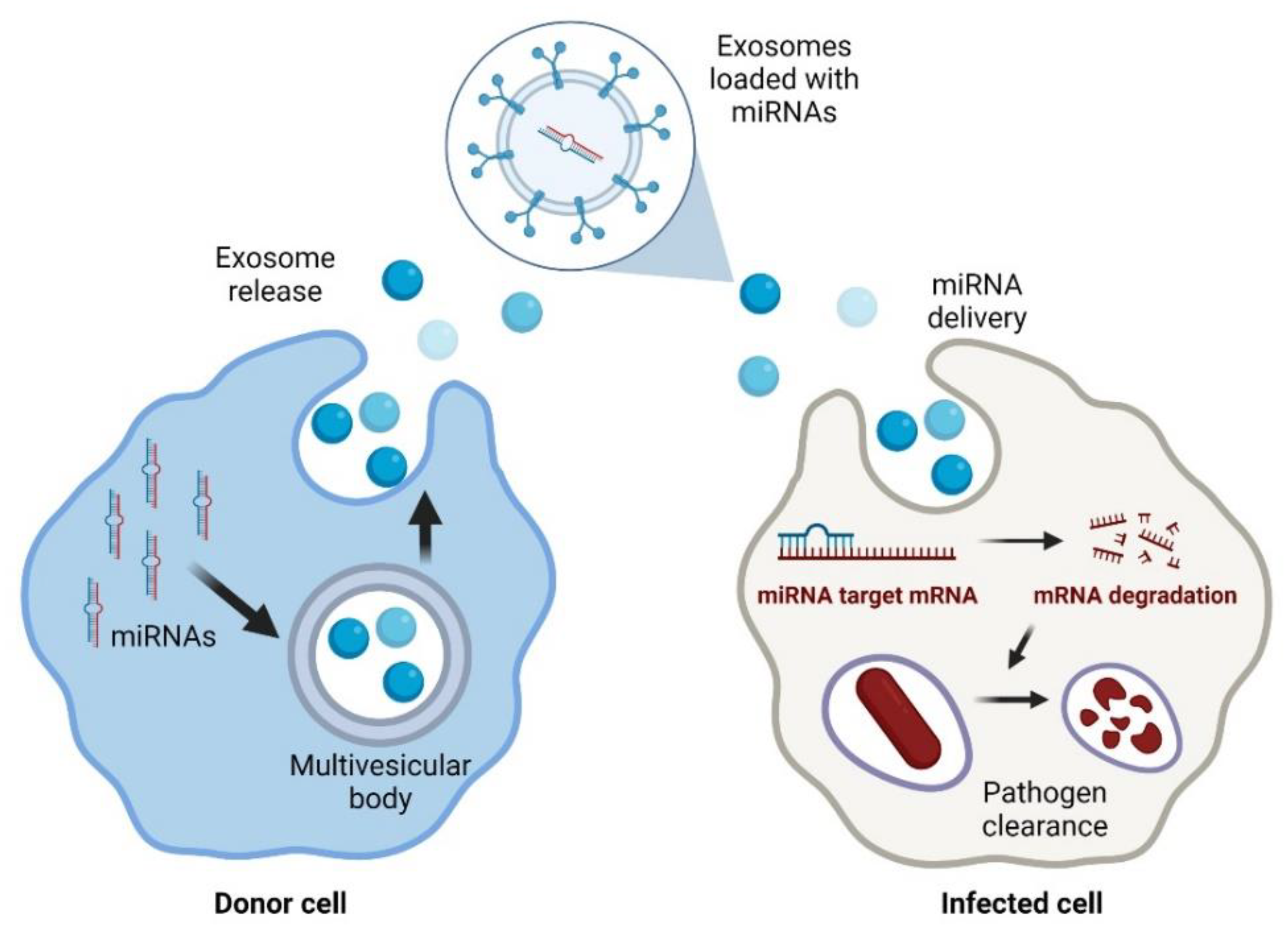{kind=link}
| Pathogen | miRNA | Targets | Mechanism of Action | References |
|---|---|---|---|---|
| Adherent–Invasive E. coli | ↑ miR-30c and miR-130a * | ↓ ATG5 and ATG16L1 * | Inhibits autophagy, facilitates bacterial intracellular survival | [29] |
| Burkholderia pseudomallei | ↑ miR-30b/c | ↓ Rab32 | Stops phagosome maturation, facilitates bacterial intracellular survival | [30] |
| ↑ miR-3473 | ↓ TRAF3 | Activates TNF-α release, cell apoptosis and inflammatory response, facilitates infection | [26] | |
| Chlamydia trachomatis | ↑ miR-30c-5p | ↓ Drp1 | Inhibition of mitochondrial fission to maintain ATP production, facilitates intracellular survival | [31] |
| ↑miR-9, miR-19 and miR-451 | ↑ NF-κB pathway | Inflammation control | [32] | |
| ↑ miR-155 and ↓ miR184 | ↓ Wnt pathway | Inflammation control | [33] | |
| Francisella tularensis | ↑ miR-155 | ↓ MyD88 and SHIP-1 | Downregulates the TLR adapter protein MyD88 and the inositol 5′-phosphatase SHIP-1 to inhibit the inflammatory response during infection | [34] |
| Helicobacter pylori | ↑ miR-25 ↑ miR-155 | ↓ KLF2 ↓ MyD88 | Kruppel-like factor 2 (KLF2) is a direct target of exosome-transmitted miR-25 in vascular endothelial cells, which may contribute to chronic heart disease Reduction of pro-inflammatory cytokine IL-8 | [35] |
| Legionella pneumophila | ↑ miR-125b, miR-221, and miR-579 | ↓ DDX58, TP53, LGALS8 and MX1 | Three miRNAs govern expression of the cytosolic RNA receptor DDX58, the tumor suppressor TP53, the antibacterial effector LGALS8, and the antiviral factor MX1 | [36] |
| Listeria monocytogenes | ↑ miR-21 | ↓ MARCKS and RhoB | The pro-phagocytic regulators myristoylated alanine-rich C-kinase substrate (MARCKS) and Ras homolog gene family, member B (RhoB) are downregulated to hinder pathogen internalization | [37] |
| ↑ miR-26a | ↓ EPHA2 | The downregulation of EPHA2 attenuates intracellular survival | [38] | |
| ↑ miR-29 | ↓ IFN-γ | Suppresses the immune response by downregulating the expression of interferon-γ | [39] | |
| Mycobacterium bovis (BCG) | ↑ miR-144-3p | ↓ ATG4a | Inhibition of autophagy, facilitates intracellular survival | [40] |
| Mycobacterium tuberculosis | ↓ miR-17-5p | ↑ Mcl-1 and ↑ STAT3 | Autophagy activation increasing the interaction of Mcl-1 and Beclin-1 | [41] |
| ↑ miR-18a | ↓ ATM | Inhibition of autophagy, facilitates intracellular survival | [42] | |
| ↑ miR-20a-3p | ↓ IKKβ | Suppression of immune response, facilitates intracellular survival | [43] | |
| ↓ miR-20b-5p | ↑ Mcl-1 | Inhibits apoptosis, facilitates intracellular survival | [44] | |
| ↑ miR-27 | ↓ CACNA2D3 | Autophagy inhibition by means of Calcium associated transporters | [45] | |
| ↑ miR-33 | ↓ ABCA1, CROT, CPT1, HADHB and PRKAA1 | Inhibiting cellular cholesterol transport and fatty acid oxidation | [46] | |
| ↑ miR-99b | ↓ Inflammatory cytokines | Inhibition of inflammation via MyD88 signaling | [47] | |
| ↓ miR-147 and miR-148a | ↑ Inflammatory cytokines | Inflammasome activation | [48,49] | |
| ↑ ↓ miR-155 # | ↑ SHIP1/Akt Pathway ↓ Rheb | Cytokine activation and control of autophagic flux | [50,51] | |
| ↑ miR-1178 | ↓TLR4-pathway | Blocks immune response | [52] | |
| ↑ miR-1958 | ↓ Atg5 | Reduction of autophagy | [53] | |
| Salmonella Typhimurium | ↑ miR-let-7i-3p | ↓ RGS2 | Inhibits bacterial replication by the modulation of endolysosomal trafficking and the vacuolar environment | [13] |
| ↓ miR-15 | ↓ E2F1 ↑ Cyclin D1 | Control of cell cycle progression, which facilitates host cell infection | [54] | |
| ↑ miR-29a | ↓ CAV2 | Caveolin 2 downregulation results in increased bacterial uptake | [55] | |
| Shigella flexneri | ↑ miR-29b-2-5p | ↓ UNC5C | Enhances filopodia production, facilitating bacterial capture and uptake | [13] |
| ↑ miR-3668, miR-4732-5p and miR-6073 | ↓ NWASP | Impairs bacterial actin-based motility, stops cell-to-cell spread, attenuates intracellular infection | [13] | |
| Staphylococcus aureus | ↑ miR-127 | ↑ STAT3 ubiquitination | Interleukin activation and bacterial clearance | [56] |
| Vibrio cholerae | ↑ miR-155 and miR-146a | ↓ NF-κB pathway | Reduction of inflammatory and immune responses in intestinal epithelial cells | [57] |
| Broad-spectrum miRNas | ↑ miR-29 ↑ miR-124 ↑ miR-302b | ↓ IFN-γ ↓ TLRs/NF-κB ↑ Cytokine genes | Inhibition of the immune response Inhibition of the immune response Activates the immune response | [39] [25,58,59] [60] |
| Lipopolysaccharide | ↑ miR-155 and miR-146a | ↓ TLR4 pathway | Negative-feedback loop of the TLR4-mediated cellular response in human monocytes exposed to lipopolysaccharide (LPS) | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mourenza, Á.; Lorente-Torres, B.; Durante, E.; Llano-Verdeja, J.; Aparicio, J.F.; Fernández-López, A.; Gil, J.A.; Mateos, L.M.; Letek, M. Understanding microRNAs in the Context of Infection to Find New Treatments against Human Bacterial Pathogens. Antibiotics 2022, 11, 356. https://doi.org/10.3390/antibiotics11030356
Mourenza Á, Lorente-Torres B, Durante E, Llano-Verdeja J, Aparicio JF, Fernández-López A, Gil JA, Mateos LM, Letek M. Understanding microRNAs in the Context of Infection to Find New Treatments against Human Bacterial Pathogens. Antibiotics. 2022; 11(3):356. https://doi.org/10.3390/antibiotics11030356
Chicago/Turabian StyleMourenza, Álvaro, Blanca Lorente-Torres, Elena Durante, Jesús Llano-Verdeja, Jesús F. Aparicio, Arsenio Fernández-López, José A. Gil, Luis M. Mateos, and Michal Letek. 2022. "Understanding microRNAs in the Context of Infection to Find New Treatments against Human Bacterial Pathogens" Antibiotics 11, no. 3: 356. https://doi.org/10.3390/antibiotics11030356