
PurN Is Involved in Antibiotic Tolerance and Virulence in Staphylococcus aureus

Abstract
:1. Introduction
2. Results
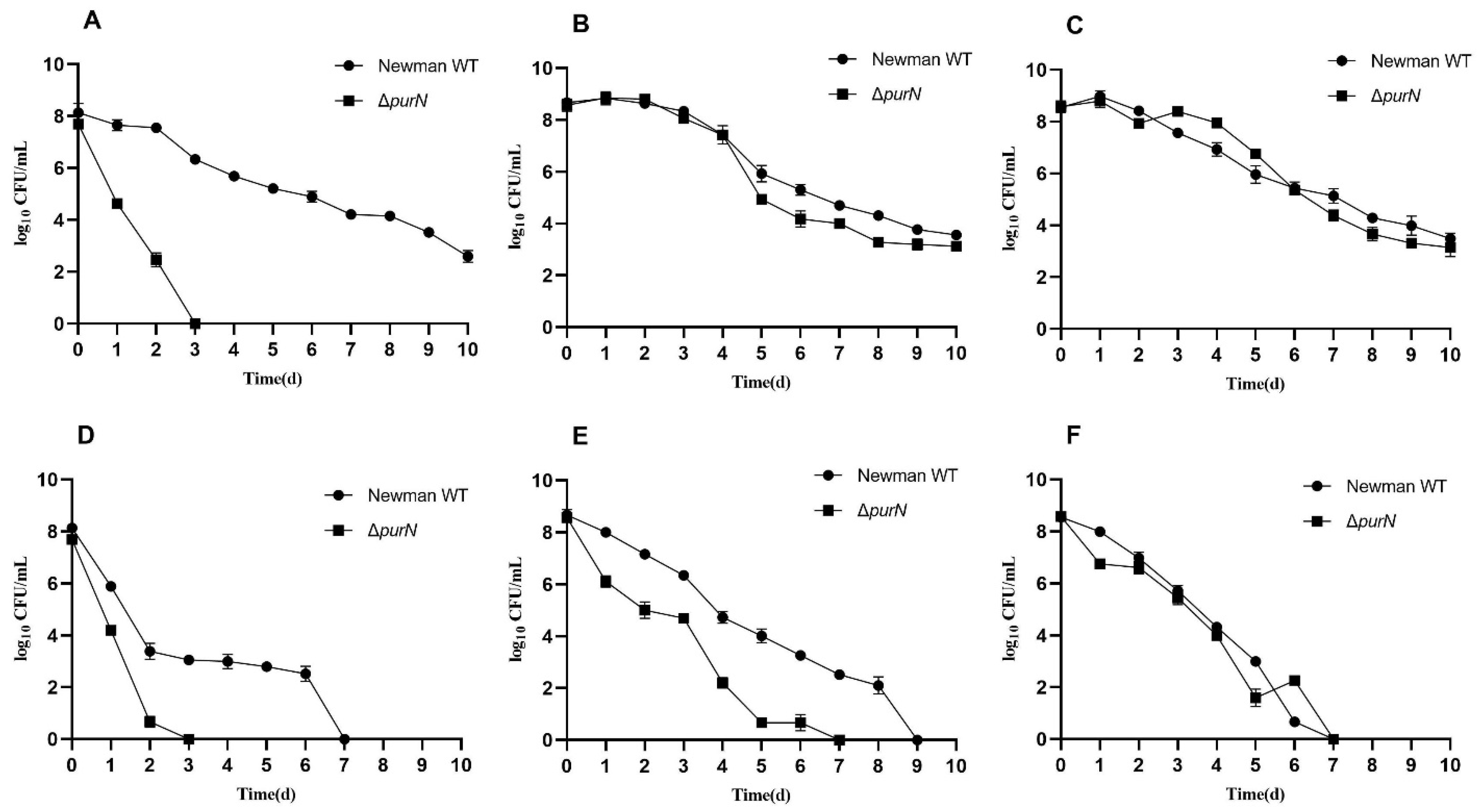
2.1. ΔpurN Had Significantly Decreased Antibiotic Tolerance
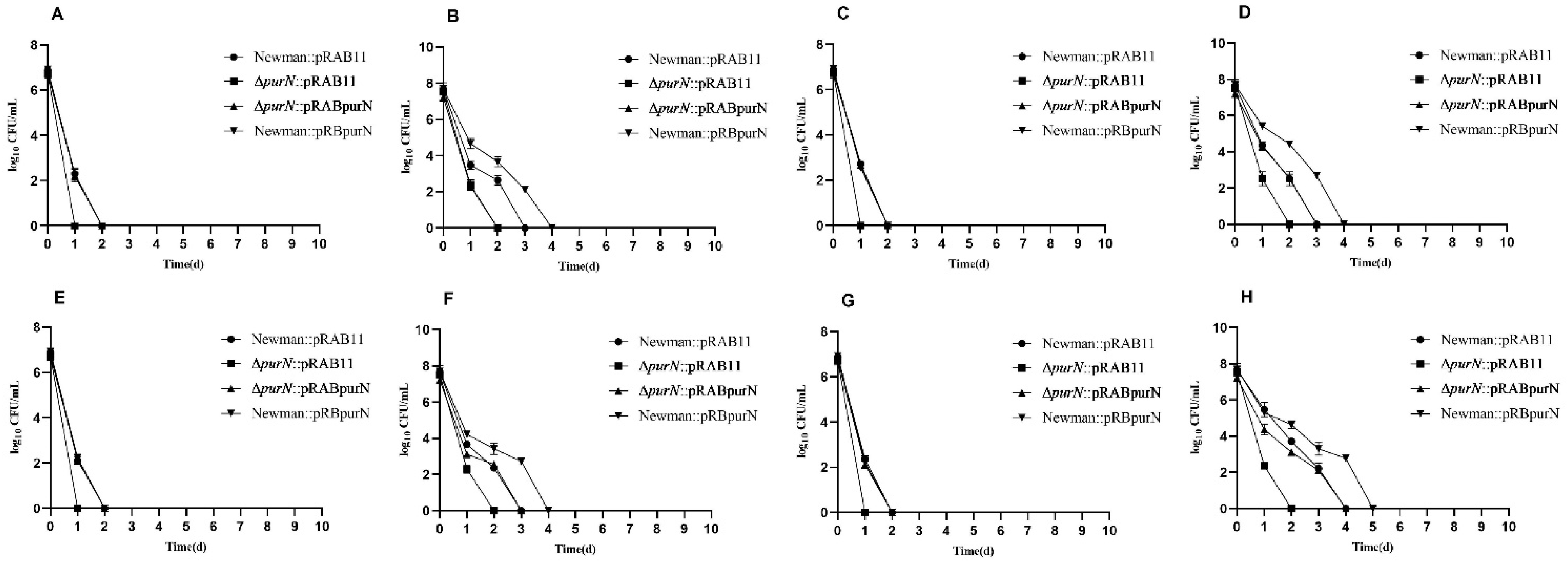
2.2. Complementation of the purN Restored Tolerance to Various Antibiotics
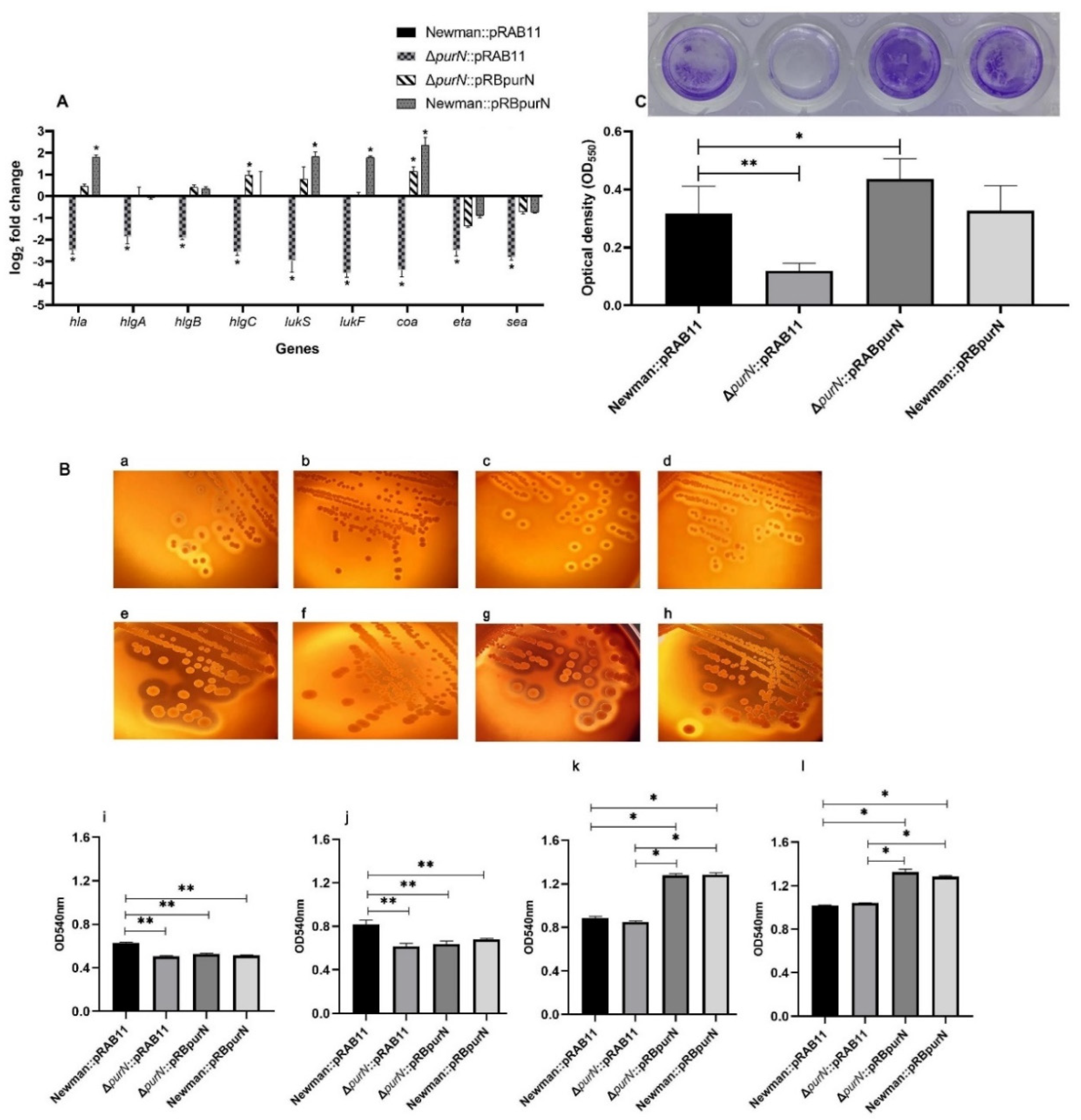
2.3. Knockout of purN Affected the Expression of Virulence Factors in S. aureus
2.4. The Ability of the ∆purN to Lyse Sheep Erythrocytes Was Significantly Reduced
2.5. Knockout of purN Affected Biofilm Formation in S. aureus
2.6. The LD50 Values of ΔpurN in Mice Were Significantly Higher Than That of Wild-Type S. aureus
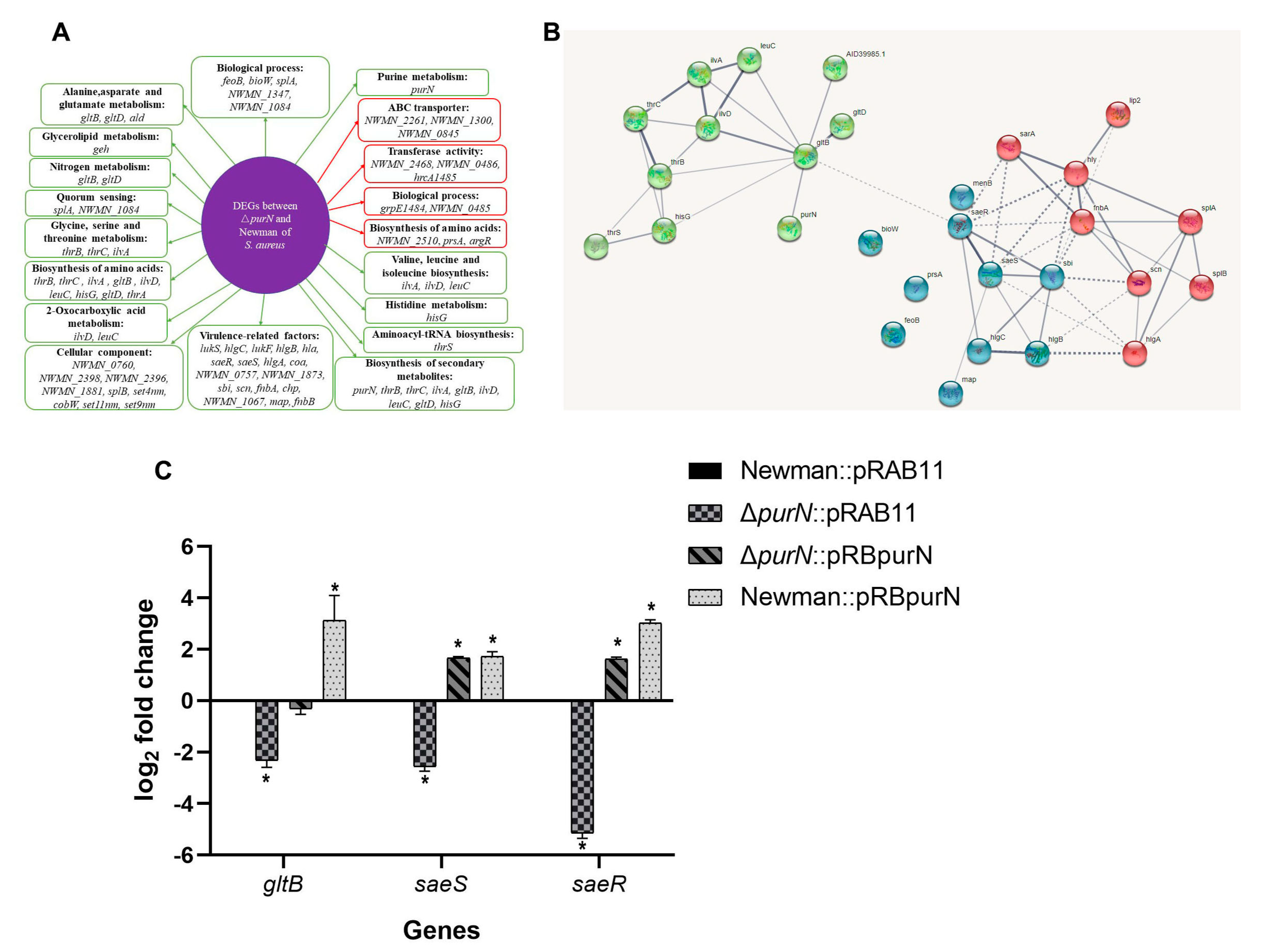
2.7. Comparative Transcriptome Analysis of the ΔpurN and the Wild-Type
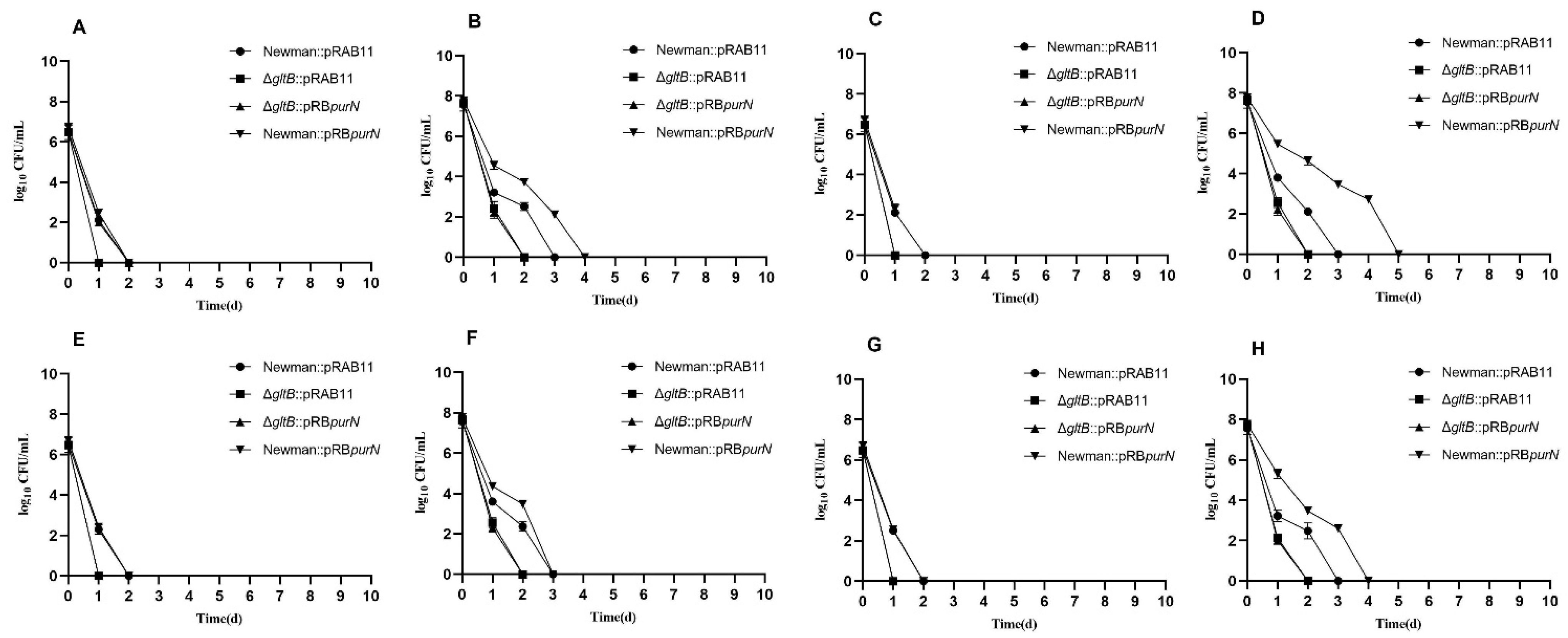
2.8. purN Affects the Persister Formation in S. aureus via gltB
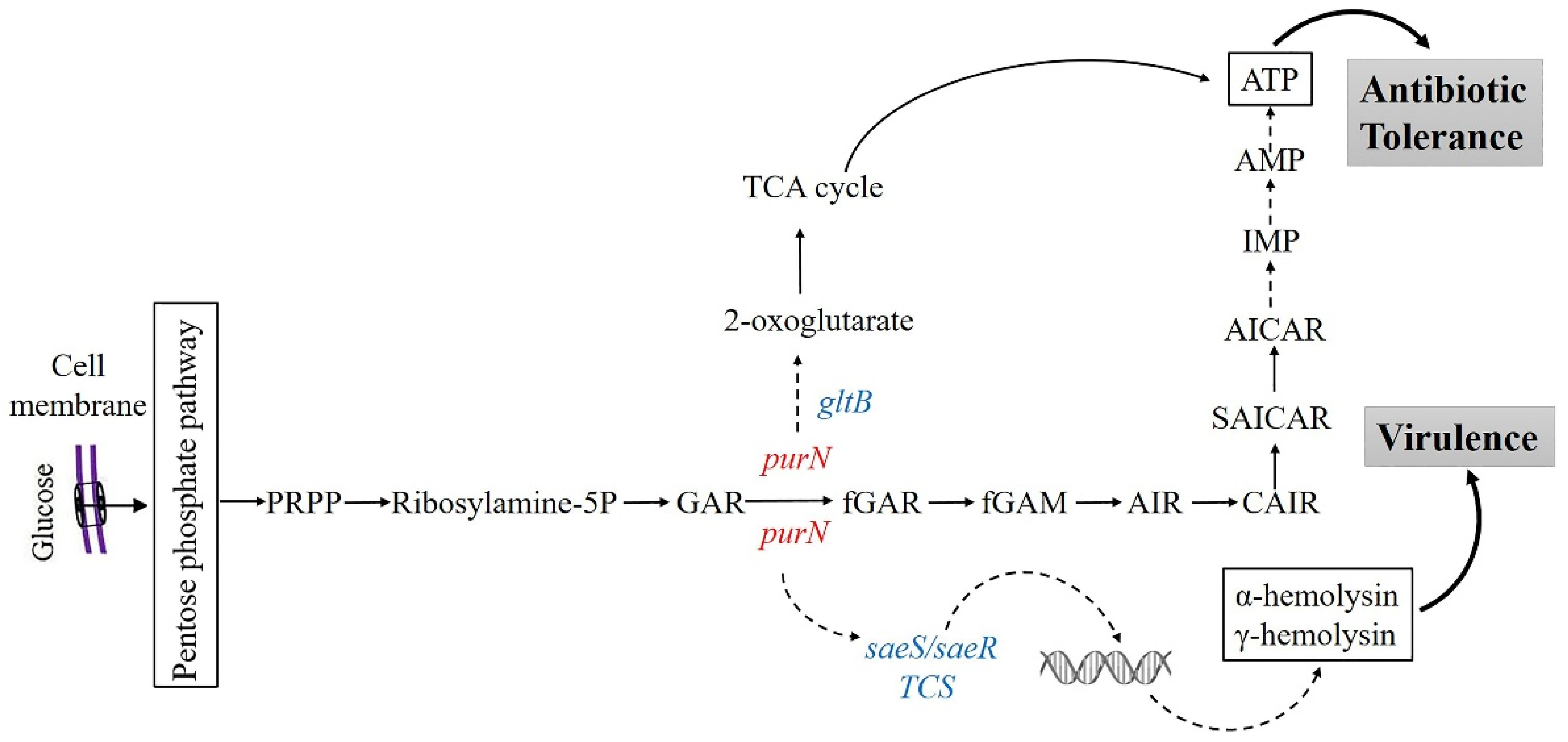
3. Discussion
4. Materials and Methods
4.1. Culture Media, Antibiotics, and Animals
4.2. Bacterial Strains and Culture Conditions
4.3. Susceptibility of Mutants to Antibiotics
4.4. Construction of Gene Knockout and Overexpression Strains
4.5. RT-qPCR Detected Genes Expression
4.6. Hemolysis Assay
4.7. Establishment of an In Vitro S. aureus Biofilm Model
4.8. Median Lethal Dose Determination
4.9. Transcriptome Analysis
4.10. Protein-Protein Interaction Network
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dayan, G.H.; Mohamed, N.; Scully, I.L.; Cooper, D.; Begier, E.; Eiden, J.; Jansen, K.U.; Gurtman, A.; Anderson, A.S. Staphylococcus aureus: The current state of disease, pathophysiology and strategies for prevention. Expert. Rev. Vaccines 2016, 15, 1373–1392. [Google Scholar] [CrossRef] [PubMed]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus hemolysins, bi-component leukocidins, and cytolytic peptides: A redundant arsenal of membrane-damaging virulence factors? Front. Cell. Infect. Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, C.P.; Williams, P.; Chan, W.C. Attenuating Staphylococcus aureus virulence gene regulation: A medicinal chemistry perspective. J. Med. Chem. 2013, 56, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Bronner, S.; Monteil, H.; Prevost, G. Regulation of virulence determinants in Staphylococcus aureus: Complexity and applications. FEMS Microbiol. Rev. 2004, 28, 183–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, A.L.; Bayer, A.S.; Zhang, G.; Gresham, H.; Xiong, Y.Q. Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 2004, 40, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.R.; Rosenthal, K.S.; Pfaller, M.A. Medical Microbiology, 8th ed.; ELSEVIER Inc.: Philadelphia, PA, USA, 2016; pp. 173–175. [Google Scholar]
- Conlon, B.P. Staphylococcus aureus chronic and relapsing infections: Evidence of a role for persister cells: An investigation of persister cells, their formation and their role in S. aureus disease. Bioessays 2014, 36, 991–996. [Google Scholar] [CrossRef]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354, aaf4268. [Google Scholar] [CrossRef]
- Defraine, V.; Fauvart, M.; Michiels, J. Fighting bacterial persistence: Current and emerging anti-persister strategies and therapeutics. Drug Resist. Updat. 2018, 38, 12–26. [Google Scholar] [CrossRef]
- Zhang, Y. Persisters, persistent infections and the Yin-Yang model. Emerg. Microbes. Infect. 2014, 3, e3. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Li, T.; Yee, R.; Yuan, Y.; Bai, C.; Cai, M.; Shi, W.; Embers, M.; Brayton, C.; Saeki, H.; et al. Stationary phase persister/biofilm microcolony of Borrelia burgdorferi causes more severe disease in a mouse model of Lyme arthritis: Implications for understanding persistence, Post-treatment Lyme Disease Syndrome (PTLDS), and treatment failure. Discov. Med. 2019, 27, 125–138. [Google Scholar] [PubMed]
- Shi, W.; Zhang, X.; Jiang, X.; Yuan, H.; Lee, J.S.; Barry, C.E., 3rd; Wang, H.; Zhang, W.; Zhang, Y. Pyrazinamide inhibits trans-translation in Mycobacterium tuberculosis. Science. 2011, 333, 1630–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Wang, X.Y.; Cui, P.; Zhang, Y.M.; Zhang, W.H.; Zhang, Y. The Agr Quorum Sensing System Represses Persister Formation through Regulation of Phenol Soluble Modulins in Staphylococcus aureus. Front. Microbiol. 2017, 8, 2189. [Google Scholar] [CrossRef]
- Shan, Y.; Brown Gandt, A.; Rowe, S.E.; Deisinger, J.P.; Conlon, B.P.; Lewis, K. ATP-Dependent Persister Formation in Escherichia coli. mBio 2017, 8, e02267-16. [Google Scholar] [CrossRef] [Green Version]
- Hazan, R.; Maura, D.; Que, Y.A.; Rahme, L.G. Assessing Pseudomonas aeruginosa Persister/antibiotic tolerant cells. Methods Mol. Biol. 2014, 1149, 699–707. [Google Scholar] [CrossRef]
- Stapels, D.A.C.; Hill, P.W.S.; Westermann, A.J.; Fisher, R.A.; Thurston, T.L.; Saliba, A.E.; Blommestein, I.; Vogel, J.; Helaine, S. Salmonella persisters undermine host immune defenses during antibiotic treatment. Science 2018, 362, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef]
- Moyed, H.S.; Bertrand, K.P. hipA, a newly recognized gene of Escherichia coli K-12 that affects frequency of persistence after inhibition of murein synthesis. J. Bacteriol. 1983, 155, 768–775. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Sim, S.; Shi, W.; Du, L.; Xing, D.; Zhang, Y. Energy production genes sucB and ubiF are involved in persister survival and tolerance to multiple antibiotics and stresses in Escherichia coli. FEMS Microbiol. Lett. 2010, 303, 33–40. [Google Scholar] [CrossRef]
- Li, J.; Ji, L.; Shi, W.; Xie, J.; Zhang, Y. Trans-translation mediates tolerance to multiple antibiotics and stresses in Escherichia coli. J. Antimicrob. Chemother. 2013, 68, 2477–2481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korch, S.B.; Henderson, T.A.; Hill, T.M. Characterization of the hipA7 allele of Escherichia coli and evidence that high persistence is governed by (p)ppGpp synthesis. Mol. Microbiol. 2003, 50, 1199–1213. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y. PhoU is a persistence switch involved in persister formation and tolerance to multiple antibiotics and stresses in Escherichia coli. Antimicrob. Agents Chemother. 2007, 51, 2092–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debbia, E.A.; Roveta, S.; Schito, A.M.; Gualco, L.; Marchese, A. Antibiotic persistence: The role of spontaneous DNA repair response. Microb. Drug Resist. 2001, 7, 335–342. [Google Scholar] [CrossRef]
- Yee, R.; Cui, P.; Shi, W.; Feng, J.; Wang, J.; Zhang, Y. Identification of a novel gene argJ involved in arginine biosynthesis critical for persister formation in Staphylococcus aureus. Discov. Med. 2020, 29, 65–77. [Google Scholar]
- Yee, R.; Cui, P.; Shi, W.; Feng, J.; Zhang, Y. Genetic Screen Reveals the Role of Purine Metabolism in Staphylococcus aureus Persistence to Rifampicin. Antibiotics 2015, 4, 627–642. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, Y.; Zhu, F.; Cheung, A.L.; Wang, G.; Bai, G.; Proctor, R.A.; Yeaman, M.R.; Bayer, A.S.; Xiong, Y.Q. New Mechanistic Insights into Purine Biosynthesis with Second Messenger c-di-AMP in Relation to Biofilm-Related Persistent Methicillin-Resistant Staphylococcus aureus Infections. mBio 2021, 12, e0208121. [Google Scholar] [CrossRef]
- Xu, T.; Han, J.; Zhang, J.; Chen, J.; Wu, N.; Zhang, W.; Zhang, Y. Absence of Protoheme IX Farnesyltransferase CtaB Causes Virulence Attenuation but Enhances Pigment Production and Persister Survival in MRSA. Front. Microbiol. 2016, 7, 1625. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bojer, M.S.; George, S.E.; Wang, Z.; Jensen, P.R.; Wolz, C.; Ingmer, H. Inactivation of TCA cycle enhances Staphylococcus aureus persister cell formation in stationary phase. Sci. Rep. 2018, 8, 10849. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Chen, J.; Chen, G.; Du, X.; Cui, P.; Wu, J.; Zhao, J.; Wu, N.; Zhang, W.; Li, M.; et al. Transposon Mutagenesis Identifies Novel Genes Associated with Staphylococcus aureus Persister Formation. Front. Microbiol. 2015, 6, 1437. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; He, L.; Shi, W.; Xu, X.; Wang, S.; Zhang, S.; Zhang, Y. Glycerol uptake is important for L-form formation and persistence in Staphylococcus aureus. PLoS ONE 2014, 9, e108325. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Wang, X.; Chen, Z.; Lyu, Z.; Lin, Z.; Zheng, J.; Wu, Y.; Deng, Q.; Yu, Z.; Zhang, Y.; et al. Staphylococcus aureus PhoU Homologs Regulate Persister Formation and Virulence. Front. Microbiol. 2020, 11, 865. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Wang, X.; Meng, L.; Zhu, M.; Wu, J.; Xu, Y.; Zhang, Y.; Zhang, W. Magnesium Links Starvation-Mediated Antibiotic Persistence to ATP. mSphere 2020, 5, e00862-19. [Google Scholar] [CrossRef] [Green Version]
- Mechler, L.; Herbig, A.; Paprotka, K.; Fraunholz, M.; Nieselt, K.; Bertram, R. A novel point mutation promotes growth phase-dependent daptomycin tolerance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2015, 59, 5366–5376. [Google Scholar] [CrossRef] [Green Version]
- Keren, I.; Kaldalu, N.; Spoering, A.; Wang, Y.; Lewis, K. Persister cells and tolerance to antimicrobials. FEMS Microbiol. Lett. 2004, 230, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamble, E.; Pardesi, K. Antibiotic Tolerance in Biofilm and Stationary-Phase Planktonic Cells of Staphylococcus aureus. Microb. Drug Resist. 2021, 27, 3–12. [Google Scholar] [CrossRef]
- Han, J.; Liu, Z.; Xu, T.; Shi, W.; Zhang, Y. A Novel LysR-Type Global Regulator RpvA Controls Persister Formation and Virulence in Staphylococcus aureus. bioRxiv 2019, 12, 861500. [Google Scholar]
- Sahukhal, G.S.; Pandey, S.; Elasri, M.O. msaABCR operon is involved in persister cell formation in Staphylococcus aureus. BMC Microbiol. 2017, 17, 218. [Google Scholar] [CrossRef] [Green Version]
- Nygaard, P.; Smith, J.M. Evidence for a novel glycinamide ribonucleotide transformylase in Escherichia coli. J. Bacteriol. 1993, 175, 3591–3597. [Google Scholar] [CrossRef] [Green Version]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; Clair, G.; Adkins, J.N.; Cheung, A.L.; Lewis, K. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat. Microbiol. 2016, 1, 16051. [Google Scholar] [CrossRef] [Green Version]
- Shan, Y.; Lazinski, D.; Rowe, S.; Camilli, A.; Lewis, K. Genetic basis of persister tolerance to aminoglycosides in Escherichia coli. mBio 2015, 6, e00078-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Zhang, Q.; Fu, Q.; Sun, M.; Huang, X. Disruption of Fis reduces bacterial persister formation by regulating glutamate metabolism in Salmonella. Microb. Pathog. 2021, 152, 104651. [Google Scholar] [CrossRef] [PubMed]
- Castaño, I.; Bastarrachea, F.; Covarrubias, A.A. gltBDF operon of Escherichia coli. J. Bacteriol. 1988, 170, 821–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, A.; Aikawa, S.; Sasaki, K.; Teramura, H.; Hasunuma, T.; Matsuda, F.; Osanai, T.; Hirai, M.Y.; Kondo, A. Rre37 stimulates accumulation of 2-oxoglutarate and glycogen under nitrogen starvation in Synechocystis sp. PCC 6803. FEBS Lett. 2014, 588, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.N.; Méndez-García, C.; Geier, R.R.; Iakiviak, M.; Chang, J.; Cann, I.; Mackie, R.I. Metabolic networks for nitrogen utilization in Prevotella ruminicola 23. Sci. Rep. 2017, 7, 7851. [Google Scholar] [CrossRef] [Green Version]
- Zeden, M.S.; Kviatkovski, I.; Schuster, C.F.; Thomas, V.C.; Fey, P.D.; Gründling, A. Identification of the main glutamine and glutamate transporters in Staphylococcus aureus and their impact on c-di-AMP production. Mol. Microbiol. 2020, 113, 1085–1100. [Google Scholar] [CrossRef]
- Bowman, L.; Zeden, M.S.; Schuster, C.F.; Kaever, V.; Gründling, A. New Insights into the Cyclic Di-adenosine Monophosphate (c-di-AMP) Degradation Pathway and the Requirement of the Cyclic Dinucleotide for Acid Stress Resistance in Staphylococcus aureus. J. Biol. Chem. 2016, 291, 26970–26986. [Google Scholar] [CrossRef] [Green Version]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 7, 29. [Google Scholar] [CrossRef]
- Dunman, P.M.; Murphy, E.; Haney, S.; Palacios, D.; Tucker-Kellogg, G.; Wu, S.; Brown, E.L.; Zagursky, R.J.; Shlaes, D.; Projan, S.J. Transcription profiling-based identification of Staphylococcus aureus genes regulated by the agr and/or sarA loci. J. Bacteriol. 2001, 183, 7341–7353. [Google Scholar] [CrossRef] [Green Version]
- Andrey, D.O.; Jousselin, A.; Villanueva, M.; Renzoni, A.; Monod, A.; Barras, C.; Rodriguez, N.; Kelley, W.L. Impact of the Regulators SigB, Rot, SarA and sarS on the Toxic Shock Tst Promoter and TSST-1 Expression in Staphylococcus aureus. PLoS ONE 2015, 10, e0135579. [Google Scholar] [CrossRef]
- Liu, Q.; Yeo, W.S.; Bae, T. The SaeRS Two-Component System of Staphylococcus aureus. Genes 2016, 7, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, N.; López-Redondo, M.; Miguel-Romero, L.; Kulhankova, K.; Cahill, M.P.; Tran, P.M.; Kinney, K.J.; Kilgore, S.H.; Al-Tameemi, H.; Herfst, C.A.; et al. The SrrAB two-component system regulates Staphylococcus aureus pathogenicity through redox sensitive cysteines. Proc. Natl. Acad. Sci. USA 2020, 117, 10989–10999. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.N.; Crosby, H.A.; Spaulding, A.R.; Salgado-Pabón, W.; Malone, C.L.; Rosenthal, C.B.; Schlievert, P.M.; Boyd, J.M.; Horswill, A.R. The Staphylococcus aureus ArlRS two-component system is a novel regulator of agglutination and pathogenesis. PLoS Pathog. 2013, 9, e1003819. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, A.T.; Calzolari, A.; Cataldi, A.A.; Bogni, C.; Nagel, R. The sae locus of Staphylococcus aureus encodes a two-component regulatory system. FEMS Microbiol. Lett. 1999, 177, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.; Li, C.; Jeong, D.; Sohn, C.; He, C.; Bae, T. In the Staphylococcus aureus two-component system sae, the response regulator SaeR binds to a direct repeat sequence and DNA binding requires phosphorylation by the sensor kinase SaeS. J. Bacteriol. 2010, 192, 2111–2127. [Google Scholar] [CrossRef] [Green Version]
- Mainiero, M.; Goerke, C.; Geiger, T.; Gonser, C.; Herbert, S.; Wolz, C. Differential target gene activation by the Staphylococcus aureus two-component system saeRS. J. Bacteriol. 2010, 192, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Yu, C.; Sun, J.; Liu, H.; Landwehr, C.; Holmes, D.; Ji, Y. Inactivation of a two-component signal transduction system, SaeRS, eliminates adherence and attenuates virulence of Staphylococcus aureus. Infect. Immun. 2006, 74, 4655–4665. [Google Scholar] [CrossRef] [Green Version]
- Geiger, T.; Goerke, C.; Mainiero, M.; Kraus, D.; Wolz, C. The virulence regulator Sae of Staphylococcus aureus: Promoter activities and response to phagocytosis-related signals. J. Bacteriol. 2008, 190, 3419–3428. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Jeong, D.W.; Liu, Q.; Yeo, W.S.; Vogl, T.; Skaar, E.P.; Chazin, W.J.; Bae, T. Calprotectin Increases the Activity of the SaeRS Two Component System and Murine Mortality during Staphylococcus aureus Infections. PLoS Pathog. 2015, 11, e1005026. [Google Scholar] [CrossRef] [Green Version]
- Bae, T.; Schneewind, O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 2006, 55, 58–63. [Google Scholar] [CrossRef]
- Han, J.; Shi, W.; Xu, X.; Wang, S.; Zhang, S.; He, L.; Sun, X.; Zhang, Y. Conditions and mutations affecting Staphylococcus aureus L-form formation. Microbiology 2015, 161, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larzábal, M.; Mercado, E.C.; Vilte, D.A.; Salazar-González, H.; Cataldi, A.; Navarro-Garcia, F. Designed coiled-coil peptides inhibit the type three secretion system of enteropathogenic Escherichia coli. PLoS ONE 2010, 5, e9046. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.M.H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1932, 27, 493–497. [Google Scholar]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmid | Relevant Genotype and Property | Source or Reference |
|---|---|---|
| S. aureus Strains | ||
| Newman | Clinical isolate, ATCC 25904 | ATCC |
| ΔpurN | Newman with a deletion of purN | This study |
| Newman::pRAB11 | Newman with pRAB11 | This study |
| ∆purN::pRAB11 | ∆purN with pRAB11 | This study |
| ∆purN::pRBpurN ∆gltB::pRAB11 ∆gltB::pRBpurN | ∆purN with pRAB11-purN ∆gltB with pRAB11 ∆gltB with pRAB11-purN | This study This study This study |
| Escherichia coli strains | ||
| DC10B | ∆dcm in the DH10B background; Dam methylation only | [33] |
| plasmids | ||
| pMX10 | A pKOR1 derivate for gene knockout, CmR, AmpR | [29] |
| pRAB11 | Atc inducible shuttle plasmid, CmR, AmpR | [15] |
| pRAB11-purN | Overexpression plasmid for purN | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Q.; Guo, L.; Dong, Y.; Bao, T.; Wang, H.; Xu, T.; Zhang, Y.; Han, J. PurN Is Involved in Antibiotic Tolerance and Virulence in Staphylococcus aureus. Antibiotics 2022, 11, 1702. https://doi.org/10.3390/antibiotics11121702
Peng Q, Guo L, Dong Y, Bao T, Wang H, Xu T, Zhang Y, Han J. PurN Is Involved in Antibiotic Tolerance and Virulence in Staphylococcus aureus. Antibiotics. 2022; 11(12):1702. https://doi.org/10.3390/antibiotics11121702
Chicago/Turabian StylePeng, Qi, Lu Guo, Yu Dong, Tingrui Bao, Huiyuan Wang, Tao Xu, Ying Zhang, and Jian Han. 2022. "PurN Is Involved in Antibiotic Tolerance and Virulence in Staphylococcus aureus" Antibiotics 11, no. 12: 1702. https://doi.org/10.3390/antibiotics11121702