
Bacteriocins Revitalize Non-Effective Penicillin G to Overcome Methicillin-Resistant Staphylococcus pseudintermedius
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Antibiotic Resistance Profile of Staphylococcus Pseudintermedius Isolates
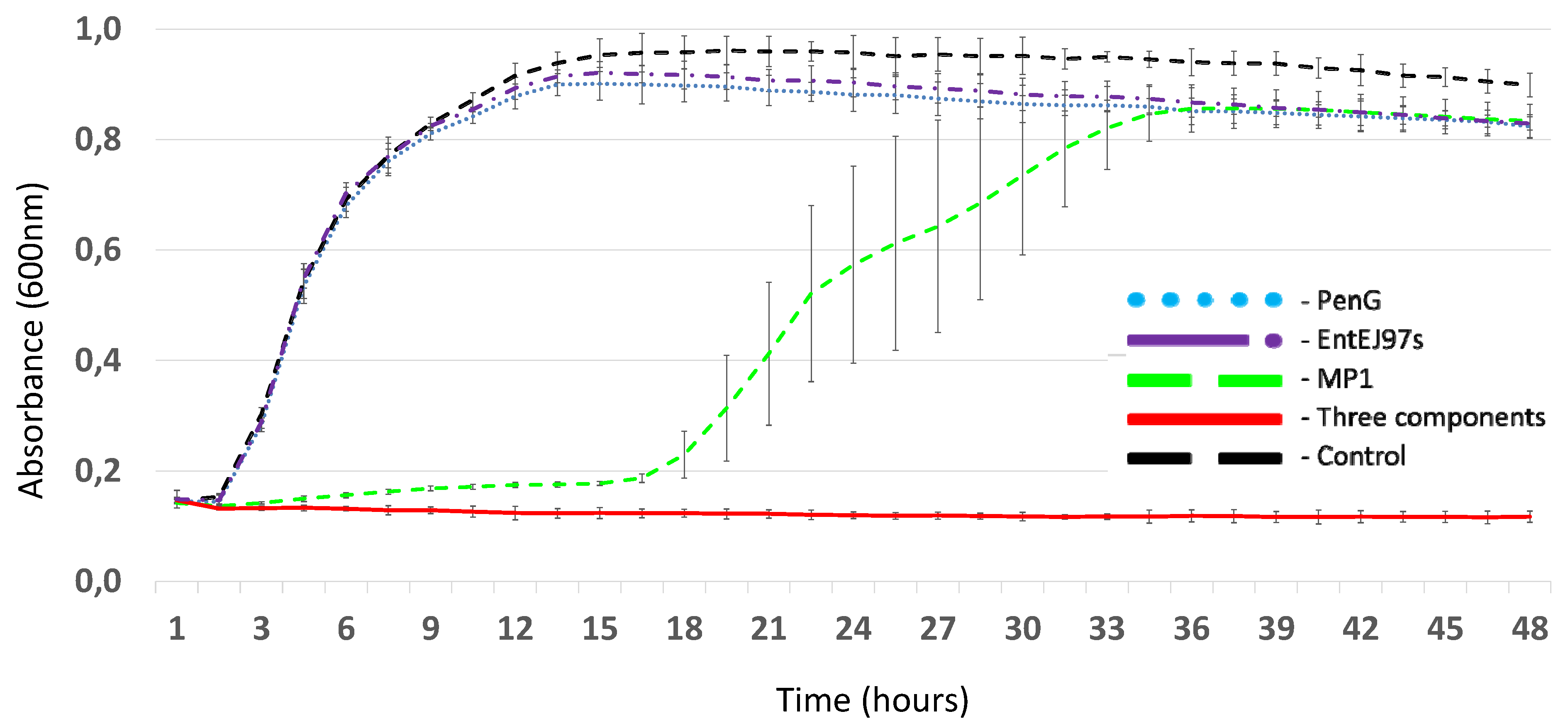
2.2. Synergetic Antimicrobial Activities
2.3. Three-Component Mixture’s Effective Inhibition of MRSP Biofilms
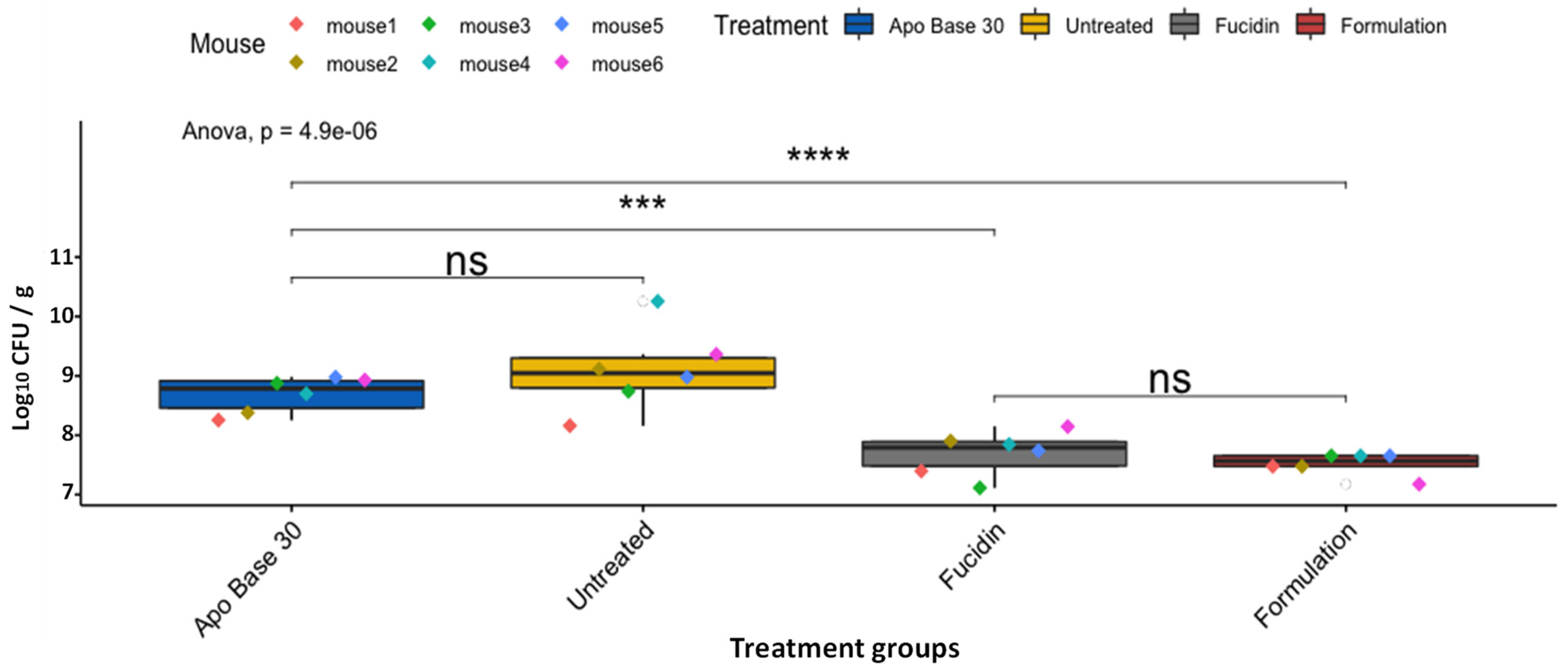
2.4. Formulation Effectiveness against MRSP in a Murine Skin Wound Infection Model
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Susceptibility Testing
4.3. Antimicrobial Agents and Formulation Vector
4.4. Synergy Assessment
4.5. Biofilm-Oriented Antimicrobial Test (BOAT)
4.6. Selection of Suitable Antimicrobial Vehicles
4.7. Murine Experiment
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lynch, S.A.; Helbig, K.J. The complex diseases of Staphylococcus pseudintermedius in canines: Where to next? Vet. Sci. 2021, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, A.; Lloyd, D.H. What has changed in canine pyoderma? A narrative review. Vet. J. 2018, 235, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhooshan, S.; Negi, V.; Khatri, P.K. Staphylococcus pseudintermedius: An undocumented, emerging pathogen in humans. GMS Hyg. Infect. Control. 2020, 15, Doc32. [Google Scholar] [CrossRef] [PubMed]
- Bemis, D.A.; Jones, R.D.; Frank, L.A.; Kania, S.A. Evaluation of Susceptibility Test Breakpoints Used to Predict mecA-Mediated Resistance in Staphylococcus Pseudintevmedius Isolated from Dogs. J. Vet. Diagn. Investig. 2009, 21, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osland, A.M.; Vestby, L.K.; Fanuelsen, H.; Slettemeås, J.S.; Sunde, M. Clonal diversity and biofilm-forming ability of methicillin-resistant Staphylococcus pseudintermedius. J. Antimicrob. Chemother. 2012, 67, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Nes, I.F.; Diep, D.B.; Håvarstein, L.S.; Brurberg, M.B.; Eijsink, V.; Holo, H. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie Leeuwenhoek 1996, 70, 113–128. [Google Scholar] [CrossRef]
- Diep, D.B.; Nes, I.F. Ribosomally synthesized antibacterial peptides in Gram positive bacteria. Curr. Drug Targets 2002, 3, 107–122. [Google Scholar] [CrossRef]
- Schofs, L.; Sparo, M.D.; Sánchez Bruni, S.F. Gram-positive bacteriocins: Usage as antimicrobial agents in veterinary medicine. Vet. Res. Commun. 2020, 44, 89–100. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef]
- Chan, D.C.K.; Burrows, L.L. Thiopeptides: Antibiotics with unique chemical structures and diverse biological activities. J. Antibiot. 2021, 74, 161–175. [Google Scholar] [CrossRef]
- Bagley, M.C.; Dale, J.W.; Merritt, E.A.; Xiong, X. Thiopeptide antibiotics. Chem. Rev. 2005, 105, 685–714. [Google Scholar] [CrossRef] [PubMed]
- Su, T.L. Micrococcin. An antibacterial substance formed by a strain of micrococcus. Br. J. Exp. Pathol. 1948, 29, 473. [Google Scholar] [PubMed]
- Harms, J.M.; Wilson, D.N.; Schluenzen, F.; Connell, S.R.; Stachelhaus, T.; Zaborowska, Z.; Spahn, C.M.; Fucini, P. Translational regulation via L11: Molecular switches on the ribosome turned on and off by thiostrepton and micrococcin. Mol. Cell 2008, 30, 26–38. [Google Scholar] [CrossRef]
- Rogers, M.J.; Cundliffe, E.; McCutchan, T.F. The antibiotic micrococcin is a potent inhibitor of growth and protein synthesis in the malaria parasite. Antimicrob. Agents Chemother. 1998, 42, 715–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, S.; Schoof, S.; Bolten, M.; Haering, C.; Takagi, M.; Shin-ya, K.; Arndt, H.-D. Molecular determinants of microbial resistance to thiopeptide antibiotics. J. Am. Chem. Soc. 2010, 132, 6973–6981. [Google Scholar] [CrossRef] [PubMed]
- Telke, A.A.; Ovchinnikov, K.V.; Vuoristo, K.S.; Mathiesen, G.; Thorstensen, T.; Diep, D.B. Over 2000-fold increased production of the leaderless bacteriocin garvicin KS by increasing gene dose and optimization of culture conditions. Front. Microbiol. 2019, 10, 389. [Google Scholar] [CrossRef]
- Gálvez, A.; Valdivia, E.; Abriouel, H.; Camafeita, E.; Mendez, E.; Martínez-Bueno, M.; Maqueda, M. Isolation and characterization of enterocin EJ97, a bacteriocin produced by Enterococcus faecalis EJ97. Arch. Microbiol. 1998, 171, 59–65. [Google Scholar] [CrossRef]
- Ovchinnikov, K.V.; Kristiansen, P.E.; Straume, D.; Jensen, M.S.; Aleksandrzak-Piekarczyk, T.; Nes, I.F.; Diep, D.B. The leaderless bacteriocin enterocin K1 is highly potent against Enterococcus faecium: A study on structure, target spectrum and receptor. Front. Microbiol. 2017, 8, 774. [Google Scholar] [CrossRef] [Green Version]
- Reinseth, I.; Tønnesen, H.H.; Carlsen, H.; Diep, D.B. Exploring the therapeutic potenital of the leaderless enterocins K1 and EJ97 in the treatment of vancomycin-resistant enterococcal infection. Front. Microbiol. 2021, 12, 649339. [Google Scholar] [CrossRef]
- Ovchinnikov, K.V.; Kranjec, C.; Thorstensen, T.; Carlsen, H.; Diep, D.B. Successful development of bacteriocins into therapeutic formulation for treatment of MRSA skin infection in a murine model. Antimicrob. Agents Chemother. 2020, 64, e00829-20. [Google Scholar] [CrossRef]
- Ovchinnikov, K.V.; Kranjec, C.; Telke, A.; Kjos, M.; Thorstensen, T.; Scherer, S.; Carlsen, H.; Diep, D.B. A strong synergy between the thiopeptide bacteriocin micrococcin P1 and rifampicin against MRSA in a murine skin infection model. Front. Immunol. 2021, 12, 676534. [Google Scholar] [CrossRef] [PubMed]
- Fugaban, J.I.I.; Vazquez Bucheli, J.E.; Holzapfel, W.H.; Todorov, S.D. Assessment of Bacteriocin-Antibiotic Synergy for the Inhibition and Disruption of Biofilms of Listeria monocytogenes and Vancomycin-Resistant Enterococcus. Microbiol. Res. 2022, 13, 480–499. [Google Scholar] [CrossRef]
- Hall, M.J.; Middleton, R.F.; Westmacott, D. The fractional inhibitory concentration (FIC) index as a measure of synergy. J. Antimicrob. Chemother. 1983, 11, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Kranjec, C.; Ovchinnikov, K.V.; Grønseth, T.; Ebineshan, K.; Srikantam, A.; Diep, D.B. A bacteriocin-based antimicrobial formulation to effectively disrupt the cell viability of methicillin-resistant Staphylococcus aureus (MRSA) biofilms. Npj Biofilms Microbiomes 2020, 6, 58. [Google Scholar] [CrossRef]
- Loeffler, A.; Baines, S.J.; Toleman, M.S.; Felmingham, D.; Milsom, S.K.; Edwards, E.A.; Lloyd, D.H. In vitro activity of fusidic acid and mupirocin against coagulase-positive staphylococci from pets. J. Antimicrob. Chemother. 2008, 62, 1301–1304. [Google Scholar] [CrossRef] [Green Version]
- Wegener, A.; Broens, E.M.; Zomer, A.; Spaninks, M.; Wagenaar, J.A.; Duim, B. Comparative genomics of phenotypic antimicrobial resistances in methicillin-resistant Staphylococcus pseudintermedius of canine origin. Vet. Microbiol. 2018, 225, 125–131. [Google Scholar] [CrossRef]
- Kjellman, E.E.; Slettemeås, J.S.; Small, H.; Sunde, M. Methicillin-resistant Staphylococcus pseudintermedius (MRSP) from healthy dogs in Norway—Occurrence, genotypes and comparison to clinical MRSP. Microbiologyopen 2015, 4, 857–866. [Google Scholar] [CrossRef]
- Diep, D.B.; Skaugen, M.; Salehian, Z.; Holo, H.; Nes, I.F. Common mechanisms of target cell recognition and immunity for class II bacteriocins. Proc. Natl. Acad. Sci. USA 2007, 104, 2384–2389. [Google Scholar] [CrossRef] [Green Version]
- Long, B.H. 2. Fusidic acid in skin and soft-tissue infections. Acta Derm. Venereol. 2008, 88, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Bassler, B.L. Surviving as a Community: Antibiotic Tolerance and Persistence in Bacterial Biofilms. Cell Host Microbe 2019, 26, 15–21. [Google Scholar] [CrossRef]
- Spoering, A.L.; Lewis, K.I.M. Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J. Bacteriol. 2001, 183, 6746–6751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, T.; Tegos, G.P.; Zhiyentayev, T.; Mylonakis, E.; Hamblin, M.R. Photodynamic therapy for methicillin-resistant Staphylococcus aureus infection in a mouse skin abrasion model. Lasers Surg. Med. 2010, 42, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Terry, J.; Neethirajan, S. A novel microfluidic wound model for testing antimicrobial agents against Staphylococcus pseudintermedius biofilms. J. Nanobiotechnol. 2014, 12, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, M.; Robres, P.; Moreno, B.; Bolea, R.; Verde, M.T.; Pérez-Laguna, V.; Aspiroz, C.; Gilaberte, Y.; Rezusta, A. Comparison of Antibacterial Activity and Wound Healing in a Superficial Abrasion Mouse Model of Staphylococcus aureus Skin Infection Using Photodynamic Therapy Based on Methylene Blue or Mupirocin or Both. Front. Med. 2021, 8, 673408. [Google Scholar] [CrossRef]
- Pietrocola, G.; Gianotti, V.; Richards, A.; Nobile, G.; Geoghegan, J.A.; Rindi, S.; Monk, I.R.; Bordt, A.S.; Foster, T.J.; Fitzgerald, J.R.; et al. Fibronectin Binding Proteins SpsD and SpsL Both Support Invasion of Canine Epithelial Cells by Staphylococcus pseudintermedius. Infect. Immun. 2015, 83, 4093–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maali, Y.; Martins-Simões, P.; Valour, F.; Bouvard, D.; Rasigade, J.P.; Bes, M.; Haenni, M.; Ferry, T.; Laurent, F.; Trouillet-Assant, S. Pathophysiological Mechanisms of Staphylococcus Non-aureus Bone and Joint Infection: Interspecies Homogeneity and Specific Behavior of S. pseudintermedius. Front. Microbiol. 2016, 7, 1063. [Google Scholar] [CrossRef]
- Bäumer, W.; Bizikova, P.; Jacob, M.; Linder, K.E. Establishing a canine superficial pyoderma model. J. Appl. Microbiol. 2017, 122, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikov, K.V.; Kristiansen, P.E.; Uzelac, G.; Topisirovic, L.; Kojic, M.; Nissen-Meyer, J.; Nes, I.F.; Diep, D.B. Defining the structure and receptor binding domain of the leaderless bacteriocin LsbB. J. Biol. Chem. 2014, 289, 23838–23845. [Google Scholar] [CrossRef] [Green Version]
- Reinseth, I.S.; Ovchinnikov, K.V.; Tønnesen, H.H.; Carlsen, H.; Diep, D.B. The Increasing Issue of Vancomycin-Resistant Enterococci and the Bacteriocin Solution. Probiotics Antimicrob. Proteins 2020, 12, 1203–1217. [Google Scholar] [CrossRef]
- Hastie, J.L.; Ellermeier, C.D. Proteolytic activation of extra cytoplasmic function (ECF) σ factors. Stress Environ. Regul. Gene Expr. Adapt. Bact. 2016, 344–351. [Google Scholar] [CrossRef]
- Paget, M.S. Bacterial Sigma Factors and Anti-Sigma Factors: Structure, Function and Distribution. Biomolecules 2015, 5, 1245–1265. [Google Scholar] [CrossRef] [PubMed]
- Peacock, S.J.; Paterson, G.K. Mechanisms of Methicillin Resistance in Staphylococcus aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, P.D.; Taylor, P.W. Methicillin resistance in Staphylococcus aureus: Mechanisms and modulation. Sci. Prog. 2002, 85, 57–72. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST) Disk Diffusion Method for Antimicrobial Susceptibility Testing. 2022. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Disk_test_documents/2022_manuals/Manual_v_10.0_EUCAST_Disk_Test_2022.pdf (accessed on 1 November 2022).
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K. CLSI methods development and standardization working group best practices for evaluation of antimicrobial susceptibility tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, Y.; Shiohira, C.M.; Yamane, N. Determination of in vitro synergy when three antimicrobial agents are combined against Mycobacterium tuberculosis. Int. J. Antimicrob. Agents 2005, 26, 292–297. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial (A) | Individual MIC, µg/mL | MIC in Mixture (A/MP1), µg/mL | FIC | MIC in Mixture (A/EntEJ97s), µg/mL | FIC |
|---|---|---|---|---|---|
| MP1 | 1.6 | - | - | - | - |
| EntEJ97s | >250 | 4/0.4 | <0.27 | >250 | - |
| Streptomycin | >250 | 4/0.8 | >0.5 | >250/>250 | >0.5 |
| Gentamicin | >250 | 8/0.8 | >0.5 | >250/>250 | >0.5 |
| Erythromycin | >250 | 8/0.8 | >0.5 | >250/>250 | >0.5 |
| Chloramphenicol | 25 | 6/0.2 | 0.38 | 25/25 | >0.5 |
| Kanamycin | 250 | 16/0.4 | 0.38 | 125/125 | >0.5 |
| Fusidic acid | 3 | 0.4/0.4 | 0.38 | 1.6/16 | >0.5 |
| Rifampicin | 0.8 | 0.2/0.2 | 0.38 | 0.4/8 | >0.5 |
| Tetracycline | 62 | 2/0.4 | 0.28 | 31/125 | >0.5 |
| Penicillin G | >250 | 8/0.4 | <0.27 | 62/62 | 0.5 |
| Three Components | (A/EntEJ97s/MP1) | ||||
| Penicillin G/EntEJ97s/MP1 | 5.0/1.0/0.2 | <0.11 | |||
| MRSP Isolate | MP1 | PenG | EntEJ97s | MP1/PenG | MP1/EntEJ97s | PenG/EntEJ97s | MP1/PenG/EntEJ97s | F. acid |
|---|---|---|---|---|---|---|---|---|
| LMGT 4218 | 3.2 | >250 | >250 | 0.4/4 | 0.4/4 | 16/16 | 0.1/1/1 | 0.4 |
| LMGT 4220 | >25 | 250 | >250 | 0.8/8 | 1.6/16 | 120/120 | 0.8/8/8 | >25 |
| LMGT 4221 | >25 | >250 | >250 | 0.4/4 | 1.6/16 | 62/62 | 0.4/4/4 | 3.2 |
| LMGT 4222 | 12 | >250 | >250 | 0.8/8 | 0.8/8 | 32/32 | 0.4/4/4 | 6.4 |
| LMGT 4223 | >25 | >250 | >250 | 0.8/8 | 0.8/8 | 62/62 | 0.4/4/4 | 3.2 |
| MRSP Isolate | MP1 | PenG | EntEJ97s | MP1/PenG | MP1/EntEJ97s | PenG/EntEJ97s | MP1/PenG/EntEJ97s | F. acid |
|---|---|---|---|---|---|---|---|---|
| LMGT 4218 | 0.9 | >3000 | >300 | 0.9/45 | 0.9/45 | 3000/300 | 0.5/25/2.5 | 0.5 |
| LMGT 4219 | 0.9 | >3000 | >300 | 0.9/45 | 0.9/45 | 750/75 | 0.5/25/2.5 | 0.25 |
| LMGT 4220 | 0.9 | >3000 | >300 | 0.9/45 | 0.9/45 | 750/75 | 0.5/12.5/2.5 | >25 |
| LMGT 4221 | 0.9 | >3000 | >300 | 0.9/45 | 0.9/45 | 750/75 | 0.5/25/2.5 | 0.1 |
| LMGT 4222 | 0.9 | >3000 | >300 | 0.9/45 | 0.9/45 | 1500/150 | 0.5/25/2.5 | 0.5 |
| LMGT 4223 | 4 | >3000 | >300 | 1.8/90 | 1.8/90 | 3000/300 | 0.5/25/2.5 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ovchinnikov, K.V.; Kranjec, C.; Thorstensen, T.; Carlsen, H.; Diep, D.B. Bacteriocins Revitalize Non-Effective Penicillin G to Overcome Methicillin-Resistant Staphylococcus pseudintermedius. Antibiotics 2022, 11, 1691. https://doi.org/10.3390/antibiotics11121691
Ovchinnikov KV, Kranjec C, Thorstensen T, Carlsen H, Diep DB. Bacteriocins Revitalize Non-Effective Penicillin G to Overcome Methicillin-Resistant Staphylococcus pseudintermedius. Antibiotics. 2022; 11(12):1691. https://doi.org/10.3390/antibiotics11121691
Chicago/Turabian StyleOvchinnikov, Kirill V., Christian Kranjec, Tage Thorstensen, Harald Carlsen, and Dzung B. Diep. 2022. "Bacteriocins Revitalize Non-Effective Penicillin G to Overcome Methicillin-Resistant Staphylococcus pseudintermedius" Antibiotics 11, no. 12: 1691. https://doi.org/10.3390/antibiotics11121691