
Virulomic Analysis of Multidrug-Resistant Klebsiella pneumoniae Isolates and Experimental Virulence Model Using Danio rerio (Zebrafish)
 , , and
, , and
Abstract
:1. Introduction
2. Results
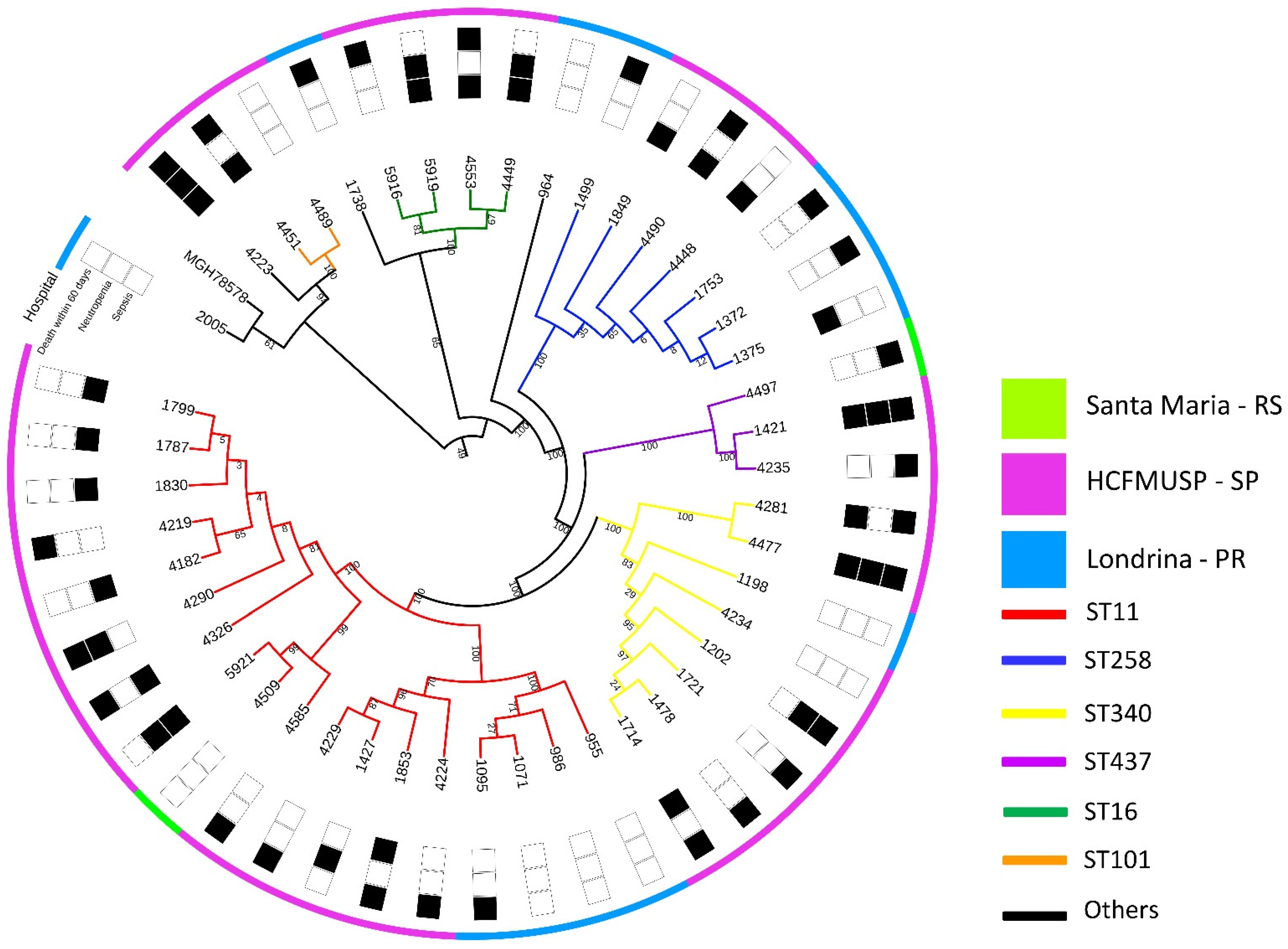
2.1. WGS, Resistance and Virulence Genes
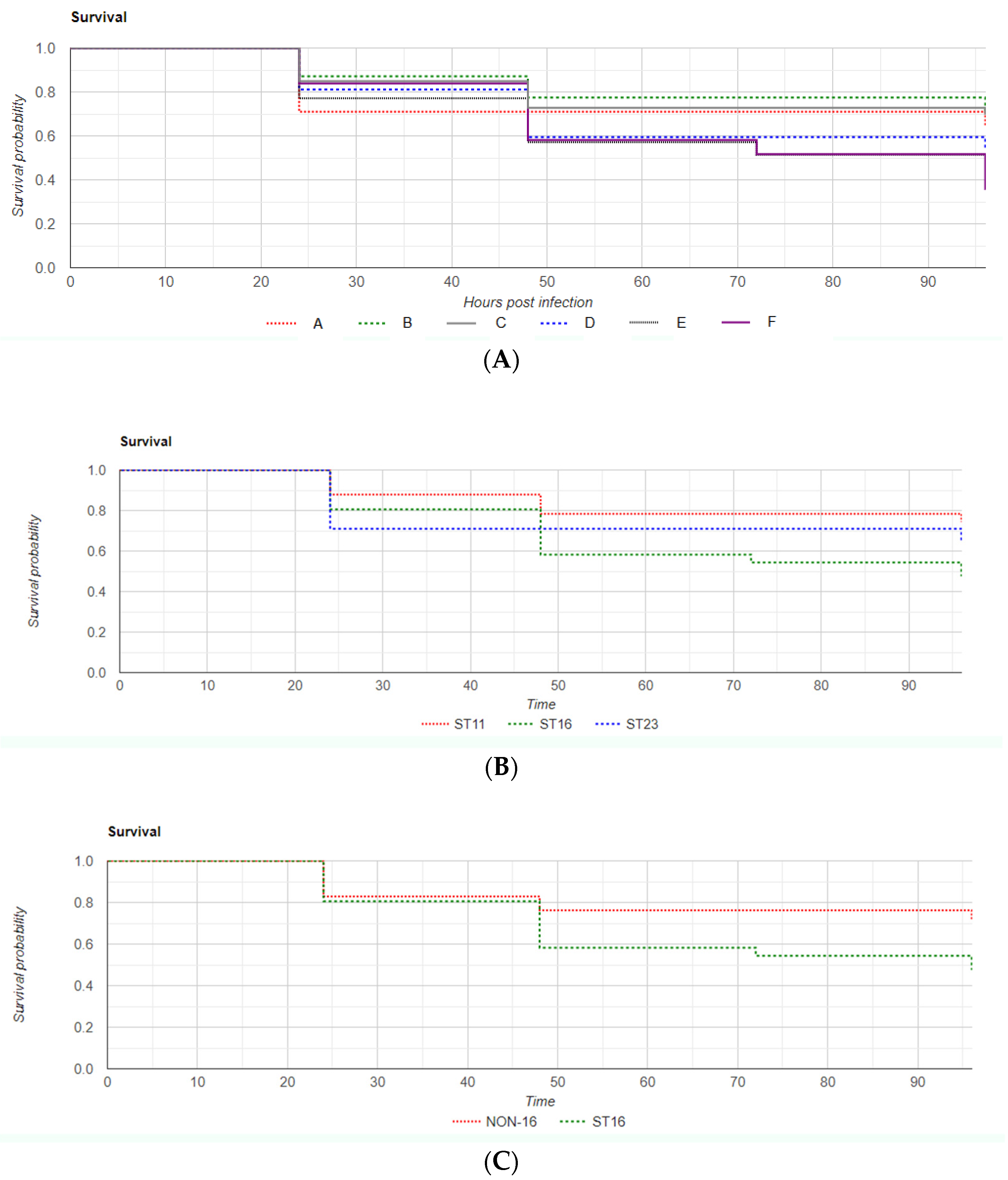
2.2. Zebrafish Virulence Model Preliminary Phase: Determination of Inoculum
2.3. Zebrafish Virulence Model
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Whole-Genome Sequencing, and Genetic Mapping
4.2. Selection of Strains for the In Vivo Experimental Model
4.3. Zebrafish: Inoculum Determination and Virulence Model
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choby, J.E.; Howard-Anderson, J.; Weiss, D.S. Hypervirulent Klebsiella pneumoniae—Clinical and molecular perspectives. J. Intern. Med. 2020, 287, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marr, C.M.; Russo, T.A. Hypervirulent Klebsiella pneumoniae: A new public health threat. Expert Rev. Anti-Infect. Ther. 2019, 17, 71–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcoleta, A.E.; Varas, M.A.; Ortiz-Severín, J.; Vásquez, L.; Berríos-Pastén, C.; Sabag, A.V.; Chávez, F.; Allende, M.L.; Santiviago, C.; Monasterio, O.; et al. Evaluating Different Virulence Traits of Klebsiella pneumoniae Using Dictyostelium discoideum and Zebrafish Larvae as Host Models. Front. Cell. Infect. Microbiol. 2018, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Prokesch, B.C.; TeKippe, M.; Kim, J.; Raj, P.D.; TeKippe, E.M.; Greenberg, D.E. Primary osteomyelitis caused by hypervirulent Klebsiella pneumoniae. Lancet Infect. Dis. 2016, 16, e190–e195. [Google Scholar] [CrossRef]
- Howlader, D.R.; Sinha, R.; Nag, D.; Majumder, N.; Mukherjee, P.; Bhaumik, U.; Maiti, S.; Withey, J.H.; Koley, H. Zebrafish as a novel model for non-typhoidal Salmonella pathogenesis, transmission and vaccine efficacy. Vaccine 2016, 34, 5099–5106. [Google Scholar] [CrossRef]
- Duggan, G.M.; Mostowy, S. Use of zebrafish to study Shigella infection. Dis. Model. Mech. 2018, 11, dmm032151. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; MacDonald, U. The Galleria mellonella Infection Model Does Not Accurately Differentiate between Hypervirulent and Classical Klebsiella pneumoniae. mSphere 2020, 5, e00850-19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, Y.; Wu, Q.; Lin, J.; Fang, R.; Bi, W.; Dong, G.; Li, J.; Zhang, Y.; Cao, J.; et al. Zebrafish and Galleria mellonella: Models to Identify the Subsequent Infection and Evaluate the Immunological Differences in Different Klebsiella pneumoniae Intestinal Colonization Strains. Front. Microbiol. 2019, 10, 2750. [Google Scholar] [CrossRef] [Green Version]
- Anes, J.; Dever, K.; Eshwar, A.; Nguyen, S.; Cao, Y.; Sivasankaran, S.K.; Sakalauskaitė, S.; Lehner, A.; Devineau, S.; Daugelavičius, R.; et al. Analysis of the Oxidative Stress Regulon Identifies soxS as a Genetic Target for Resistance Reversal in Multidrug-Resistant Klebsiella pneumoniae. mBio 2021, 12, e00867-21. [Google Scholar] [CrossRef]
- Al-Farsi, H.M.; Al-Adwani, S.; Ahmed, S.; Vogt, C.; Ambikan, A.T.; Leber, A.; Al-Jardani, A.; Al-Azri, S.; Al-Muharmi, Z.; Toprak, M.S.; et al. Effects of the Antimicrobial Peptide LL-37 and Innate Effector Mechanisms in Colistin-Resistant Klebsiella pneumoniae With mgrB Insertions. Front. Microbiol. 2019, 10, 2632. [Google Scholar] [CrossRef]
- Andrey, D.O.; Dantas, P.P.; Martins, W.B.S.; De Carvalho, F.M.; Almeida, L.G.P.; Sands, K.; Portal, E.; Sauser, J.; Cayô, R.; Nicolás, M.F.; et al. An Emerging Clone, Klebsiellapneumoniae Carbapenemase 2–Producing K. pneumoniae Sequence Type 16, Associated with High Mortality Rates in a CC258-Endemic Setting. Clin. Infect. Dis. 2020, 71, e141–e150. [Google Scholar] [CrossRef] [PubMed]
- Cornet, C.; Calzolari, S.; Miñana-Prieto, R.; Dyballa, S.; van Doornmalen, E.; Rutjes, H.; Savy, T.; D’Amico, D.; Terriente, J. ZeGlobalTox: An Innovative Approach to Address Organ Drug Toxicity Using Zebrafish. Int. J. Mol. Sci. 2017, 18, 864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorlien, E.L.; Witucki, M.A.; Ogas, J. Efficient Production and Identification of CRISPR/Cas9-generated Gene Knockouts in the Model System Danio rerio. J. Vis. Exp. 2018, 138, e56969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, H.-C.; Lee, C.-Z.; Ma, L.-C.; Fang, C.-T.; Chang, S.-C.; Wang, J.-T. Isolation of a Chromosomal Region of Klebsiella pneumoniae Associated with Allantoin Metabolism and Liver Infection. Infect. Immun. 2004, 72, 3783–3792. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.-L.; Chan, K.-S.; Ko, W.-C.; Lee, C.-C.; Chuang, Y.-C. Lower Prevalence of Diabetes Mellitus in Patients with Klebsiella pneumoniae Primary Liver Abscess Caused by Isolates of K1/K2 than with Non-K1/K2 Capsular Serotypes. Clin. Infect. Dis. 2007, 45, 1529–1530. [Google Scholar] [CrossRef]
- Russo, T.A.; Marr, C.M. Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 2019, 32, e00001-19. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Shields, R.K.; Clancy, C.J.; Pasculle, A.W.; Press, E.G.; Haidar, G.; Hao, B.; Chen, L.; Kreiswirth, B.N.; Nguyen, M.H. Verification of Ceftazidime-Avibactam and Ceftolozane-Tazobactam Susceptibility Testing Methods against Carbapenem-Resistant Enterobacteriaceae and Pseudomonas aeruginosa. J. Clin. Microbiol. 2018, 56, e01093-17. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontéen, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- OECD. Validation Report (Phase 1) for the Zebrafish Embryo Toxicity Test: Part I and Part II; Series on Testing and Assessment No. 157; OECD: Paris, France, 2011. [Google Scholar]
- OECD. Validation Report (Phase 2) for the Zebrafish Embryo Toxicity Test: Part I and Part II (Annexes); Series on Testing and Assessment No. 179; OECD: Paris, France, 2012. [Google Scholar]
- Schubert, S.; Keddig, N.; Hanel, R.; Kammann, U. Microinjection into zebrafish embryos (Danio rerio)—A useful tool in aquatic toxicity testing? Environ. Sci. Eur. 2014, 26, 1. [Google Scholar] [CrossRef] [Green Version]
- Rizek, C.; Fu, L.; Dos Santos, L.C.; Leite, G.; Ramos, J.; Rossi, F.; Guimaraes, T.; Levin, A.S.; Costa, S.F. Characterization of carbapenem-resistant Pseudomonas aeruginosa clinical isolates, carrying multiple genes coding for this antibiotic resistance. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Statistical Test Calculators—Check Assumptions, Interpret Results. Statistics Kingdom. 2017. Available online: https://www.statskingdom.com/ (accessed on 10 February 2021).



{kind=link}
{kind=link}
| Strain | MLST | Siderophore | Regulation | Allantoin Metabolism | Auto Transporter | Efflux Pump AcrAB | Stress Adaptation |
|---|---|---|---|---|---|---|---|
| “A” | 23 | Aerobactin, Yersiniabactin | rcsA, rcsB | Yes | No | Yes | Yes |
| “B” | 11 | Enterobactin, Yersiniabactin | rcsA, rcsB | No | cah | Yes | Yes |
| “C” | 11 | Enterobactin, Yersiniabactin | rcsA, rcsB | No | cah | Yes | Yes |
| “D” | 16 | None | rcsA, rcsB | No | No | Yes | No |
| “E” | 16 | None | rcsA, rcsB | No | No | Yes | No |
| “F” | 16 | Enterobactin, Yersiniabactin | rcsA, rcsB | No | No | Yes | No |
| Group ID | ST | Injected Embryos | Alive 24 hpi | PMR | Alive 48 hpi | PMR | Alive 72 hpi | PMR | Alive 96 hpi | Endpoint Mortality |
|---|---|---|---|---|---|---|---|---|---|---|
| “A” | 23 | 31 | 22 | 29% | 22 | 29% | 22 | 29% | 20 | 35% |
| “B” | 11 | 31 | 27 | 13% | 24 | 22% | 24 | 22% | 22 | 29% |
| “C” | 11 | 33 | 28 | 15% | 24 | 27% | 24 | 27% | 23 | 30% |
| “D” | 16 | 37 | 30 | 19% | 22 | 40% | 22 | 40% | 20 | 46% |
| “E” | 16 | 35 | 27 | 23% | 20 | 43% | 18 | 48% | 18 | 48% |
| “F” | 16 | 31 | 26 | 16% | 18 | 42% | 16 | 48% | 11 | 64% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, E.L.T.; Rizek, C.F.; Espinoza, E.S.; Marchi, A.P.; Noguera, S.V.; Côrtes, M.F.; Fernandes, B.H.V.; Guimarães, T.; de Maio Carrilho, C.M.D.; Neto, L.V.P.; et al. Virulomic Analysis of Multidrug-Resistant Klebsiella pneumoniae Isolates and Experimental Virulence Model Using Danio rerio (Zebrafish). Antibiotics 2022, 11, 1567. https://doi.org/10.3390/antibiotics11111567
Duarte ELT, Rizek CF, Espinoza ES, Marchi AP, Noguera SV, Côrtes MF, Fernandes BHV, Guimarães T, de Maio Carrilho CMD, Neto LVP, et al. Virulomic Analysis of Multidrug-Resistant Klebsiella pneumoniae Isolates and Experimental Virulence Model Using Danio rerio (Zebrafish). Antibiotics. 2022; 11(11):1567. https://doi.org/10.3390/antibiotics11111567
Chicago/Turabian StyleDuarte, Edson Luiz Tarsia, Camila Fonseca Rizek, Evelyn Sanchez Espinoza, Ana Paula Marchi, Saidy Vasconez Noguera, Marina Farrel Côrtes, Bianca H. Ventura Fernandes, Thais Guimarães, Claudia M. D. de Maio Carrilho, Lauro V. Perdigão Neto, and et al. 2022. "Virulomic Analysis of Multidrug-Resistant Klebsiella pneumoniae Isolates and Experimental Virulence Model Using Danio rerio (Zebrafish)" Antibiotics 11, no. 11: 1567. https://doi.org/10.3390/antibiotics11111567