
Occurrence and Molecular Characterization of Multidrug-Resistant Vegetable-Borne Listeria monocytogenes Isolates
 ,
,
Abstract
:1. Introduction
2. Results
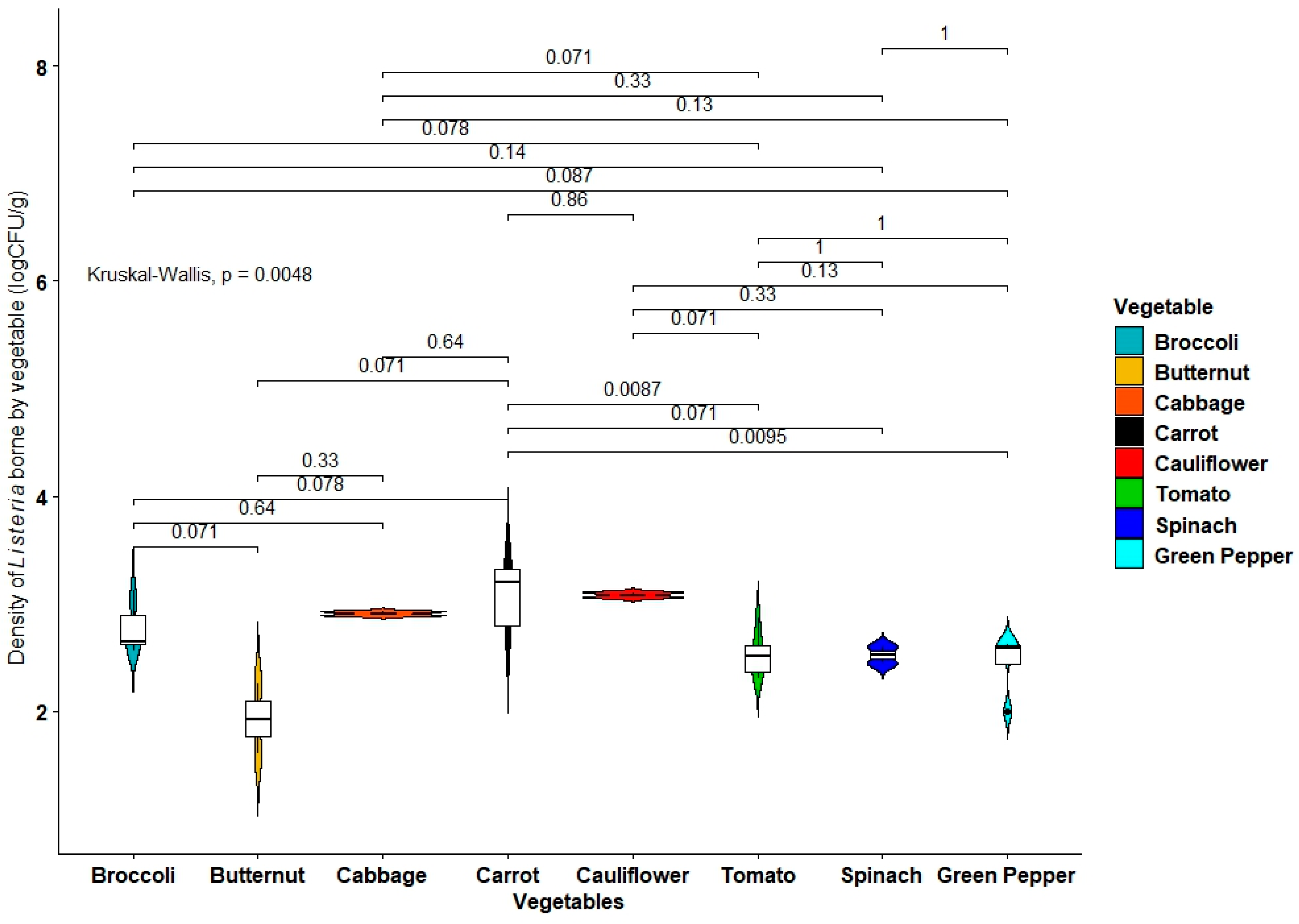
2.1. Prevalence of Listeria Species in Vegetables
2.2. Incidence of Vegetable-Borne Listeria monocytogenes
2.3. Antibiotic Susceptibility Patterns of L. monocytogenes
2.4. Multiple Antibiotic Resistance Phenotypes (MARP) of L. monocytogenes
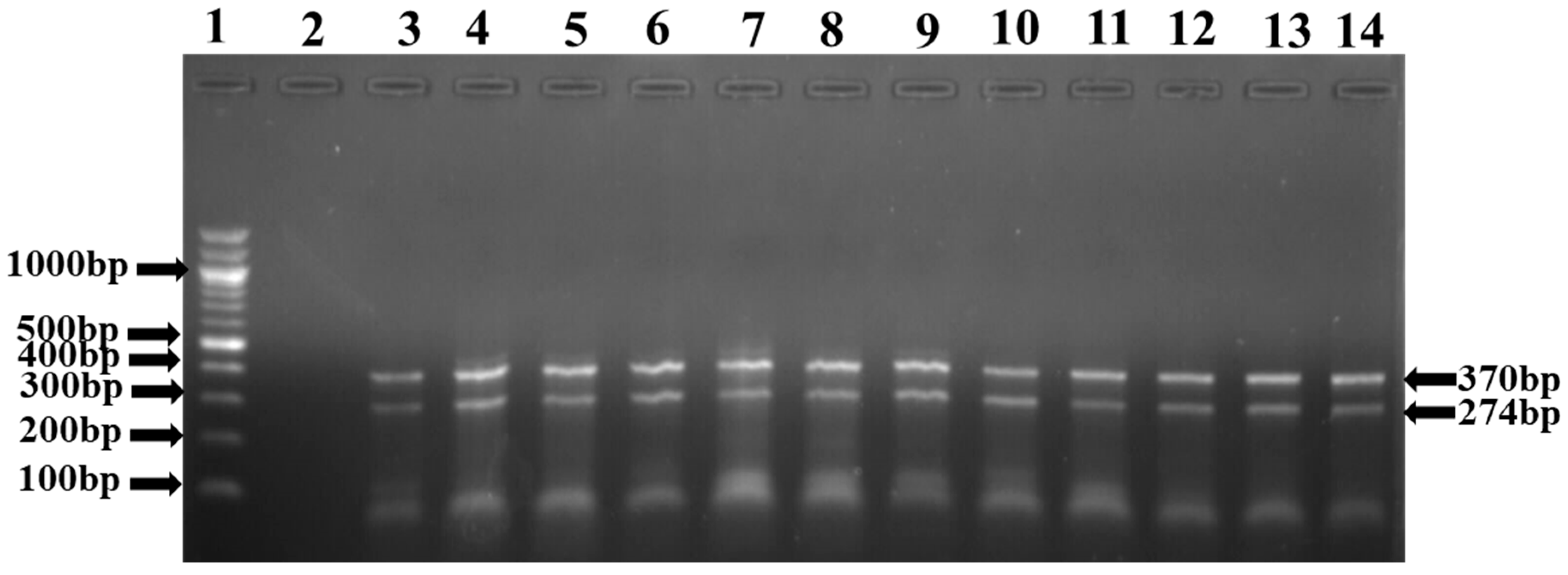
2.5. Prevalence of Clinical Antimicrobial Resistance Gene (ARGs) Subtypes
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Genet. 2017, 16, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, A.; Moura, A.; Vales, G.; Tessaud-Rita, N.; Aguilhon, C.; Lecuit, M. Listeria thailandensis sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 74–81. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of antibiotic re-sistance in Listeria monocytogenes isolated from food products: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeyaletchumi, P.; Tunung, R.; Margaret, S.P.; Son, R.; Ghazali, F.M.; Cheah, Y.K.; Nakaguchi, Y.; Malakar, P.K. Quantification of Listeria monocytogenes in salad vegetables by MPN-PCR. Int. Food Res. J. 2010, 17, 281–286. [Google Scholar]
- Maurice Bilung, L.; Sin Chai, L.; Tahar, A.S.; Ted, C.K.; Apun, K. Prevalence, genetic heterogeneity, and antibiotic resistance profile of Listeria spp. and Listeria monocytogenes at farm level: A highlight of ERIC-and BOX-PCR to reveal genetic diversity. Biomed. Res Int. 2018, 2018, 3067494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, K.J.; Wałecka-Zacharska, E.; Chen, J.C.; Katarzyna, K.-P.; Devlieghere, F.; Van Meervenne, E.; Osek, J.; Wieczorek, K.; Bania, J. Listeria monocytogenes—An examination of food chain factors potentially contributing to antimicrobial resistance. Food Microbiol. 2016, 54, 178–189. [Google Scholar] [CrossRef]
- Gómez, D.; Azón, E.; Marco, N.; Carramiñana, J.J.; Rota, C.; Ariño, A.; Yangüela, J. Antimicrobial resistance of Listeria monocytogenes and Listeria innocua from meat products and meat-processing environment. Food Microbiol. 2014, 42, 61–65. [Google Scholar] [CrossRef]
- Gandhi, M.; Chikindas, M.L. Listeria: A foodborne pathogen that knows how to survive. Int. J. Food Microbiol. 2007, 113, 1–15. [Google Scholar] [CrossRef] [PubMed]
- De Roever, C. Microbiological safety evaluations and recommendations on fresh produce. Food Control 1998, 9, 321–347. [Google Scholar] [CrossRef]
- Şanlıbaba, P.; Tezel, B.U.; Çakmak, G.A. Prevalence and antibiotic resistance of Listeria monocytogenes isolated from ready-to-eat foods in Turkey. J. Food Qual. 2018, 2018, 7693782. [Google Scholar] [CrossRef] [Green Version]
- Al-Nabulsi, A.A.; Osaili, T.M.; Shaker, R.R.; Olaimat, A.N.; Jaradat, Z.W.; Elabedeen, N.A.Z.; Holley, R.A. Effects of osmotic pressure, acid, or cold stresses on antibiotic susceptibility of Listeria monocytogenes. Food Microbiol. 2015, 46, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Meloni, D.; Galluzzo, P.; Mureddu, A.; Piras, F.; Griffiths, M.; Mazzette, R. Listeria monocytogenes in RTE foods marketed in Italy: Prevalence and automated EcoRI ribotyping of the isolates. Int. J. Food Microbiol. 2009, 129, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Gooneratne, S.R.; Hussain, M.A. Listeria monocytogenes in Fresh Produce: Outbreaks, Prevalence and Contamination Levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meldrum, R.; Little, C.; Sagoo, S.; Mithani, V.; McLauchlin, J.; de Pinna, E. Assessment of the microbiological safety of salad vegetables and sauces from kebab take-away restaurants in the United Kingdom. Food Microbiol. 2009, 26, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Krumperman, P.H. Multiple Antibiotic Resistance Indexing of Escherichia coli to Identify High-Risk Sources of Fecal Contamination of Foodst. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Goñi, P.; López, P.; Sánchez, C.; Gómez-Lus, R.; Becerril, R.; Nerín, C. Antimicrobial activity in the vapour phase of a combination of cinnamon and clove essential oils. Food Chem. 2009, 116, 982–989. [Google Scholar] [CrossRef]
- Sangeetha, M.S.; Shubha, G. Molecular Identification of Listeria Species from Vegetables Marketed in Mysore, Karnataka, India. Res. J. Chem. Sci. 2014, 3, 1–4. [Google Scholar]
- Onyilokwu, S.A.; Lawan, F.A.; Hambali, I.U.; Mailafiya, S.; Adamu, N.B.; Atsanda, N.N.; Jauro, S. Phenotypic Characterisation and Distribution Pattern of Listeria Species Isolated from Food Samples Retailed in Markets and Central Abattoir in Maiduguri, Nigeria. Alex. J. Vet. Sci. 2016, 51, 122–126. [Google Scholar]
- Rahimi, E.; Momtaz, H.; Sharifzadeh, A.; Behzadnia, A.; Ashtari, M.S.; Esfahani, S.Z.; Riahi, M.; Momeni, M. Prevalence and antimicrobial resistance of Listeria species isolated from traditional dairy products in Chahar Mahal & Bakhtiyari, Iran. Bulg. J. Vet. Med. 2012, 15, 115–122. [Google Scholar]
- Chitarra, W.; Decastelli, L.; Garibaldi, A.; Gullino, M.L. Potential uptake of Escherichia coli O157: H7 and Listeria monocytogenes from growth substrate into leaves of salad plants and basil grown in soil irrigated with contaminated water. Int. J. Food Microbiol. 2014, 189, 139–145. [Google Scholar] [CrossRef]
- Szabo, E.; Scurrah, K.; Burrows, J. Survey for psychrotrophic bacterial pathogens in minimally processed lettuce. Lett. Appl. Microbiol. 2000, 30, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, E.; Pérez-Rodríguez, F.; Valero, A.; Garcı´a-Gimeno, R.; Zurera, G. Growth of Listeria monocytogenes on shredded, ready-to-eat iceberg lettuce. Food Control 2008, 19, 487–494. [Google Scholar] [CrossRef]
- Moreno, L.Z.; Paixão, R.; Gobbi, D.D.S.; Raimundo, D.C.; Ferreira, T.P.; Moreno, A.M.; Reis, C.M.F.; Matté, G.R.; Matté, M.H. Original Article Characterization of antibiotic resistance in Listeria spp. isolated from slaughterhouse environments, pork and human infections. J. Infect. Dev. Ctries 2014, 8, 416–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cetinkaya, F.; Elal Mus, T.; Yibar, A.; Guclu, N.; Tavsanli, H.; Cibik, R. Prevalence, serotype identification by multiplex polymerase chain reaction and antimicrobial resistance patterns of Listeria monocytogenes isolated from retail foods. J. Food Saf. 2014, 34, 42–49. [Google Scholar] [CrossRef]
- Li, Q.; Sherwood, J.; Logue, C. Antimicrobial resistance of Listeria spp. recovered from processed bison. Lett. Appl. Microbiol. 2007, 44, 86–91. [Google Scholar] [CrossRef]
- Kuan, C.H.; Rukayadi, Y.; Ahmad, S.H.; Wan Mohamed Radzi, C.W.J.; Kuan, C.S.; Yeo, S.K.; Thung, T.Y.; New, C.Y.; Chang, W.S.; Loo, Y.Y.; et al. Antimicrobial resistance of Listeria monocytogenes and Salmonella Enteritidis isolated from vegetable farms and retail markets in Malaysia. Int. Food Res. J. 2017, 24, 1831–1839. [Google Scholar]
- Janakiraman, V. Listeriosis in pregnancy: Diagnosis, treatment, and prevention. Rev. Obstet. Gynecol. 2008, 1, 179–185. [Google Scholar] [PubMed]
- Pagliano, P.; Arslan, F.; Ascione, T. Epidemiology and treatment of the commonest form of listeriosis: Meningitis and bacteraemia. Infez. Med. 2017, 25, 210–216. [Google Scholar]
- Adefisoye, M.A.; Okoh, A.I. Identification and antimicrobial resistance prevalence of pathogenic Escherichia coli strains from treated wastewater effluents in Eastern Cape, South Africa. MicrobiologyOpen 2016, 5, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Christopher, A.F.; Hora, S.; Ali, Z. Investigation of plasmid profile, antibiotic susceptibility pattern multiple antibiotic resistance index calculation of Escherichia coli isolates obtained from different human clinical specimens at tertiary care hospital in Bareilly-India. Ann. Trop. Med. Public Health 2013, 6, 285. [Google Scholar] [CrossRef]
- Kayode, A.J.; Okoh, A.I. Incidence and genetic diversity of multi-drug resistant Listeria monocyto-genes isolates recovered from fruits and vegetables in the Eastern Cape Province, South Africa. Int. J. Food Microbiol. 2022, 363, 109513. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Nam, H.; Nguyen, L.; Tamilselvam, B.; Murinda, S.; Oliver, S. Prevalence of Antimicrobial Resistance Genes in Listeria monocytogenes Isolated from Dairy Farms. Foodborne Pathog. Dis. 2005, 2, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, E.; Courvalin, P. Antibiotic resistance in Listeria spp. Antimicrob. Agents Chemother. 1999, 43, 2103–2108. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, T.L.; Carbone, M.; Fera, M.T.; Irrera, G.P.; Gugliandolo, C. Distribution of potentially pathogenic bacteria as free liv-ing and plankton associated in a marine coastal zone. J. Appl. Microbiol. 2004, 97, 354–361. [Google Scholar] [CrossRef]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K. CLSI Methods Development and Standardization Working Group Best Practices for Evaluation of Antimicrobial Susceptibility Tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [Green Version]
- Obaidat, M.M.; Salman, A.E.B.; Lafi, S.Q.; Al-Abboodi, A.R. Characterization of Listeria monocytogenes from three countries and antibiotic resistance differences among countries and Listeria monocytogenes serogroups. Lett. Appl. Microbiol. 2015, 60, 609–614. [Google Scholar] [CrossRef]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baddour, M.M.; Abuelkheir, M.M.; Fatani, A.J. Comparison of mecA Polymerase Chain Reaction with Phenotypic Methods for the Detection of Methicillin-Resistant Staphylococcus aureus. Curr. Microbiol. 2007, 55, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Falbo, V.; Carattoli, A.; Tosini, F.; Pezzella, C.; Dionisi, A.M.; Luzzi, I. Antibiotic Resistance Conferred by a Conjugative Plasmid and a Class I Integron in Vibrio cholerae O1 El Tor Strains Isolated in Albania and Italy. Antimicrob. Agents Chemother. 1999, 43, 693–696. [Google Scholar] [CrossRef] [Green Version]
- Jannine, K.B.; Jeremy, L.P.; Sashindran, A.; Ruth, M.H. Commensal Escherichia coli of healthy humans: A reservoir for antibiotic-resistance determinants. J. Med. Microbiol. 2010, 59, 1331–1339. [Google Scholar]
- Maynard, C.; Fairbrother, J.M.; Bekal, S.; Sanschagrin, F.; Levesque, R.C.; Brousseau, R.; Masson, L.; Lariviëre, S.; Harel, J. Antimicrobial Resistance Genes in Enterotoxigenic Escherichia coli O149:K91 Isolates Obtained over a 23-Year Period from Pigs. Antimicrob. Agents Chemother. 2003, 47, 3214–3221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, L.-K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes 2001, 15, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Post, V.; Hall, R.M. AbaR5, a large multiple-antibiotic resistance region found in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 2667–2671. [Google Scholar] [CrossRef] [Green Version]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR Assay for Simultaneous Detection of Nine Clinically Relevant Antibiotic Resistance Genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [PubMed]
- Velusamy, S.; Barbara, E.G.; Mark, J.L.; Lien, T.N.; Susan, I.H.; Ynte, H.S.; Stephen, P.O. Phenotypic and genotypic antimicrobial resistance patterns of Escherichia coli isolated from dairy cows with mastitis. Vet. Microbiol. 2007, 124, 319–328. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Antibiotics | Resistance Patterns | Frequency | MARI |
|---|---|---|---|
| 3 | E-PG-VA | 3 | 0.18 |
| 4 | E-PG-T-VA | 1 | 0.25 |
| 4 | E-PG-TS-VA | 2 | 0.25 |
| 5 | E-PG-T-TS-VA | 6 | 0.25 |
| 5 | AUG-E-PG-T-VA | 3 | 0.31 |
| 5 | CIP-E-PG-TS-VA | 3 | 0.31 |
| 5 | E-NI-PG-TS-VA | 1 | 0.31 |
| 5 | E-NI-PG-T-VA | 1 | 0.31 |
| 6 | CXM-E-PG-T-TS-VA | 1 | 0.31 |
| 6 | C-E-PG-T-TS-VA | 1 | 0.31 |
| 6 | E-NI-PG-T-TS-VA | 6 | 0.31 |
| 6 | AUG-E-PG-T-TS-VA | 1 | 0.31 |
| 6 | CIP-E-PG-T-TS-VA | 1 | 0.31 |
| 6 | KF-E-PG-T-TS-VA | 4 | 0.38 |
| 6 | AUG-CXM-E-PG-T-VA | 2 | 0.38 |
| 6 | CIP-E-MEM-PG-TS-VA | 1 | 0.38 |
| 6 | CIP-E-LEV-PG-TS-VA | 2 | 0.38 |
| 6 | CIP-E-NI-PG-TS-VA | 1 | 0.38 |
| 6 | AUG-CXM-KF-PG-T-VA | 1 | 0.38 |
| 6 | AUG-CXM-E-NI-PG-T | 1 | 0.38 |
| 6 | CXM-C-E-PG-T-TS | 1 | 0.38 |
| 6 | AUG-CXM-KF-CIP-MEM-PG | 1 | 0.38 |
| 6 | AUG-CXM-LEV-NI-PG-T | 1 | 0.38 |
| 6 | AUG-KF-E-PG-T-VA | 1 | 0.38 |
| 6 | AUG-C-E-PG-T-VA | 1 | 0.38 |
| 6 | AUG-CXM-KF-E-PG-T | 1 | 0.38 |
| 6 | AUG-CXM-KF-CIP-MEM-PG | 1 | 0.38 |
| 6 | AUG-CXM-LEV-NI-PG-T | 1 | 0.38 |
| 6 | AUG-KF-E-PG-T-VA | 1 | 0.38 |
| 6 | AUG-C-E-PG-T-VA | 1 | 0.38 |
| 6 | AUG-CXM-KF-E-PG-T | 1 | 0.38 |
| 7 | AUG-KF-CIP-E-PG-T-VA | 1 | 0.44 |
| 7 | AUG-CXM-KF-E-MEM-NI-PG | 1 | 0.44 |
| 7 | AUG-CIP-E-NI-PG-TS-VA | 1 | 0.44 |
| 7 | C-E-NI-PG-T-TS-VA | 3 | 0.44 |
| 7 | C-CIP-E-LEV-PG-TS-VA | 1 | 0.44 |
| 7 | CIP-E-LEV-NI-PG-TS-VA | 2 | 0.44 |
| 7 | AUG-KF-E-NI-PG-T-VA | 1 | 0.44 |
| 7 | AUG-CXM-E-PG-T-TS-VA | 1 | 0.44 |
| 7 | AUG-E-NI-PG-T-TS-VA | 5 | 0.44 |
| 7 | C-E-MEM-PG-T-TS-VA | 1 | 0.44 |
| 7 | CXM-E-NI-PG-T-TS-VA | 2 | 0.44 |
| 7 | AUG-CXM-KF-E-NI-PG-VA | 1 | 0.44 |
| 8 | C-CIP-E-NI-PG-T-TS-VA | 1 | 0.5 |
| 8 | CIP-E-MEM-NI-PG-T-TS-VA | 1 | 0.5 |
| 8 | E-K-MEM-NI-PG-T-TS-VA | 2 | 0.5 |
| 8 | KF-C-E-NI-PG-T-TS-VA | 1 | 0.5 |
| 8 | AUG-CXM-KF-E-NI-PG-T-VA | 1 | 0.5 |
| 8 | AUG-CXM-KF-E-PG-T-TS-VA | 2 | 0.5 |
| 8 | AUG-KF-E-NI-PG-T-TS-VA | 1 | 0.5 |
| 8 | AUG-E-MEM-NI-PG-T-TS-VA | 1 | 0.5 |
| 8 | AUG-CXM-E-NI-PG-T-TS-VA | 1 | 0.5 |
| 8 | AUG-CXM-KF-CIP-E-PG-TS-VA | 1 | 0.5 |
| 8 | AUG-CXM-KF-E-NI-PG-TS-VA | 1 | 0.5 |
| 8 | AUG-KF-E-LEV-PG-T-TS-VA | 1 | 0.5 |
| 8 | C-E-LEV-NI-PG-T-TS-VA | 1 | 0.5 |
| 9 | AUG-CXM-KF-E-MEM-NI-PG-T-VA | 1 | 0.56 |
| 9 | AUG-CXM-KF-E-NI-PG-T-TS-VA | 1 | 0.56 |
| 9 | AUG-CXM-E-MEM-NI-PG-T-TS-VA | 1 | 0.56 |
| 9 | AUG-CXM-E-GM-K-PG-T-TS-VA | 1 | 0.56 |
| 9 | AUG-CXM-KF-CIP-E-NI-PG-TS-VA | 1 | 0.56 |
| 9 | AUG-CXM-CIP-E-NI-PG-T-TS-VA | 1 | 0.56 |
| 9 | AUG-CXM-E-LEV-NI-PG-T-TS-VA | 1 | 0.56 |
| 9 | AUG-C-E-LEV-NI-PG-T-TS-VA | 1 | 0.56 |
| 9 | C-E-K-MEM-NI-PG-T-TS-VA | 1 | 0.56 |
| 9 | CXM-CIP-E-LEV-NI-PG-T-TS-VA | 1 | 0.56 |
| 10 | AUG-CXM-KF-E-LEV-NI-PG-T-TS-VA | 1 | 0.63 |
| 10 | AK-AUG-KF-E-MEM-NI-PG-T-TS-VA | 1 | 0.63 |
| 10 | AK-AUG-KF-CIP-E-LEV-PG-T-TS-VA | 1 | 0.63 |
| 11 | AUG-CXM-KF-C-E-K-NI-PG-T-TS-VA | 6 | 0.69 |
| 13 | AUG-CXM-KF-CIP-E-GM-K-LEV-NI-PG-T-TS-VA | 1 | 0.81 |
| Antimicrobial Family | Antimicrobial Agent | Antimicrobial Resistance Gene | No. of Positive Isolates | Percentage (%) |
|---|---|---|---|---|
| Tetracyclines | Tetracycline (n = 86) | tetA | 51 | 59.3 |
| tetB | 0 | 0 | ||
| tetC | 37 | 43 | ||
| tetD | 37 | 43 | ||
| tetK | 1 | 1.2 | ||
| tetM | 47 | 54.7 | ||
| Aminoglycosides | Amikacin (n = 1) Gentamycin (n = 2) Kanamycin (n = 11) | aacC2 | 0 | 0 |
| aphA1 | 0 | 0 | ||
| aphA2 | 5 | 41.7 | ||
| aadA | 4 | 33.3 | ||
| strA | 0 | 0 | ||
| Beta-lactams | Amoxicillin/Clavulanic Acid (n = 53) Penicillin G (n = 108) | blaTEM | 19 | 17.6 |
| blaZ | 0 | 0 | ||
| ampC | 4 | 3.7 | ||
| TEM | 83 | 76.9 | ||
| SHV | 0 | 0 | ||
| OXA1-like | 0 | 0 | ||
| GES | 0 | 0 | ||
| PER | 6 | 0 | ||
| VEB | 0 | 0 | ||
| Cephems | Meropenem (n = 50) | ACC | 0 | 0 |
| FOX | 16 | 32 | ||
| MOX | 0 | 0 | ||
| DHA | 3 | 6 | ||
| CIT | 24 | 48 | ||
| EBC | 0 | 0 | ||
| Phenicols | Chloramphenicol (n = 20) | cmlA1 | 7 | 35 |
| catI | 0 | 0 | ||
| catII | 1 | 5 | ||
| Sulfanomides | Trimethoprim-Sulfamethoxazole (n = 86) | SulI | 86 | 100 |
| sulII | 0 | 0 |
| PCR Name | Primer | Primer Sequence | Amplicon Size (bp) | Cycling Conditions |
|---|---|---|---|---|
| Multiplex I TEM, SHV, and OXA-1-like | blaTEM, blaSHV, blaOXA-1 | F: ATTTCCGTGTCGCCCTTATTC R: CGTTCATCCATAGTTGCCTGAC F: AGCCGCTTGAGCAAATTAAAC R: ATCCCGCAGATAAATCACCAC F: GGCACCAGATTCAACTTTCAAG R: GACCCCAAGTTTCCTGTAAGTG | 800 713 564 | Initial denaturation at 94 °C for 10 min; 30 cycles of 94 °C for 40 s, 60 °C for 40 s and 72 °C for 60 s; and a final elongation step at 72 °C for 7 min |
| Multiplex II FOX, CIT, and EBC | blaFOX blaCIT blaEBC | F: CTACAGTGCGGGTGGTTT R: CTATTTGCGGCCAGGTGA F: CGAAGAGGCAATGACCAGAC R: ACGGACAGGGTTAGGATAGY b F: CGGTAAAGCCGATGTTGCG R: AGCCTAACCCCTGATACA | 162 538 683 | Initial denaturation at 94 °C for 10 min; 30 cycles of 94 °C for 40 s, 60 °C for 40 s and 72 °C for 60 s; and a final elongation step at 72 °C for 7 min |
| Simplex CTX_M group 8/2 | blaCTX-M | F: AACRCRCAGACGCTCTAC b R: TCGAGCCGGAASGTGTYAT b | 326 | Initial denaturation at 94 °C for 10 min; 30 cycles of 94 °C for 40 s, 60 °C for 40 s and 72 °C for 60 s; and a final elongation step at 72 °C for 7 min |
| Multiplex III IMP, VIM, and KPC | blaIMP blaVIM blaKPC | F: TTGACACTCCATTTACDG b R: GATYGAGAATTAAGCCACYCT b F: GATGGTGTTTGGTCGCATA R: CGAATGCGCAGCACCAG F: CATTCAAGGGCTTTCTTGCTGC R: ACGACGGCATAGTCATTTGC | 139 390 538 | Initial denaturation at 94 °C for 10 min; 30 cycles of 94 °C for 40 s, 55 °C for 40 s and 72 °C for 60 s; and a final elongation step at 72 °C for 7 min |
| Multiplex IV GES and PER | blaGES BlaPER | F: AGTCGGCTAGACCGGAAAG R: TTTGTCCGTGCTCAGGAT F: GCTCCGATAATGAAAGCGT R: TTCGGCTTGACTCGGCTGA | 399 520 | Initial denaturation at 94 °C for 10 min; 30 cycles of 94 °C for 40 s, 60 °C for 40 s and 72 °C for 60 s; and a final elongation step at 72 °C for 7 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntshanka, Z.; Ekundayo, T.C.; du Plessis, E.M.; Korsten, L.; Okoh, A.I. Occurrence and Molecular Characterization of Multidrug-Resistant Vegetable-Borne Listeria monocytogenes Isolates. Antibiotics 2022, 11, 1353. https://doi.org/10.3390/antibiotics11101353
Ntshanka Z, Ekundayo TC, du Plessis EM, Korsten L, Okoh AI. Occurrence and Molecular Characterization of Multidrug-Resistant Vegetable-Borne Listeria monocytogenes Isolates. Antibiotics. 2022; 11(10):1353. https://doi.org/10.3390/antibiotics11101353
Chicago/Turabian StyleNtshanka, Zizipho, Temitope C. Ekundayo, Erika M. du Plessis, Lise Korsten, and Anthony I. Okoh. 2022. "Occurrence and Molecular Characterization of Multidrug-Resistant Vegetable-Borne Listeria monocytogenes Isolates" Antibiotics 11, no. 10: 1353. https://doi.org/10.3390/antibiotics11101353