Real-Time Temperature Sensing Using a Ratiometric Dual Fluorescent Protein Biosensor


Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. General Differential Scanning Fluorimetry (DSF) Assays
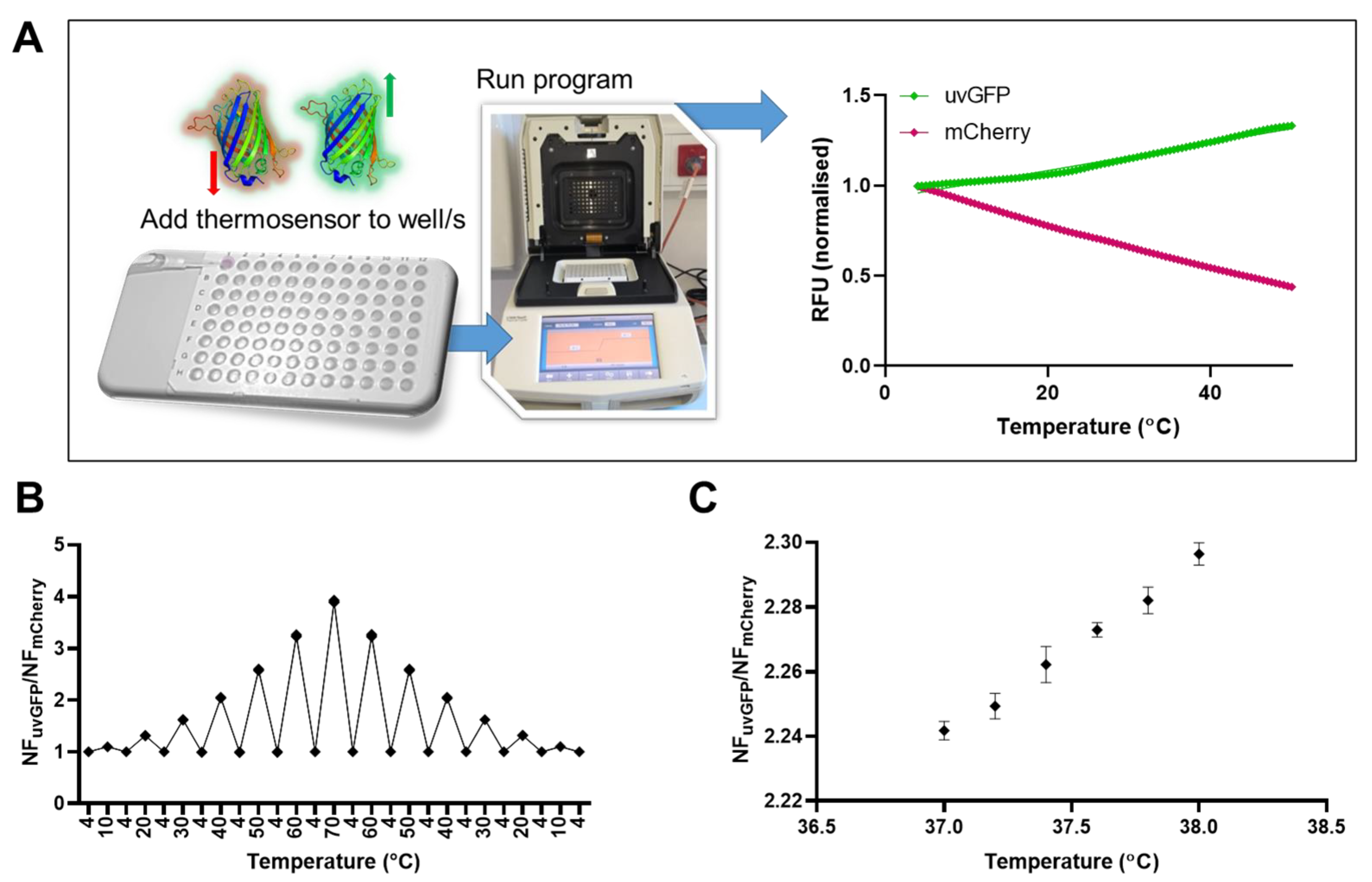
2.3. Sensitivity to Temperature Change and Resolution
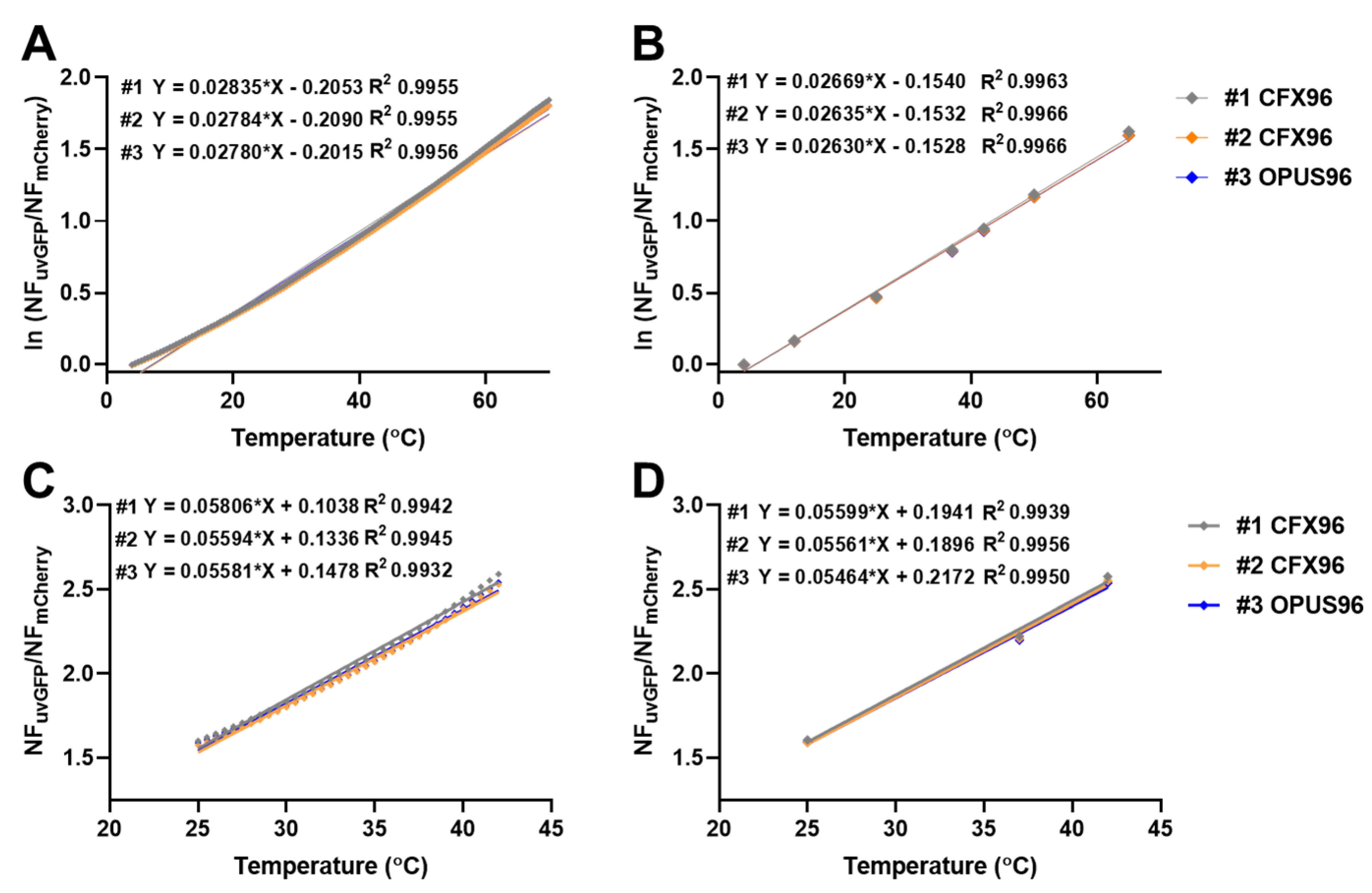
2.4. Relative Temperature Quantification
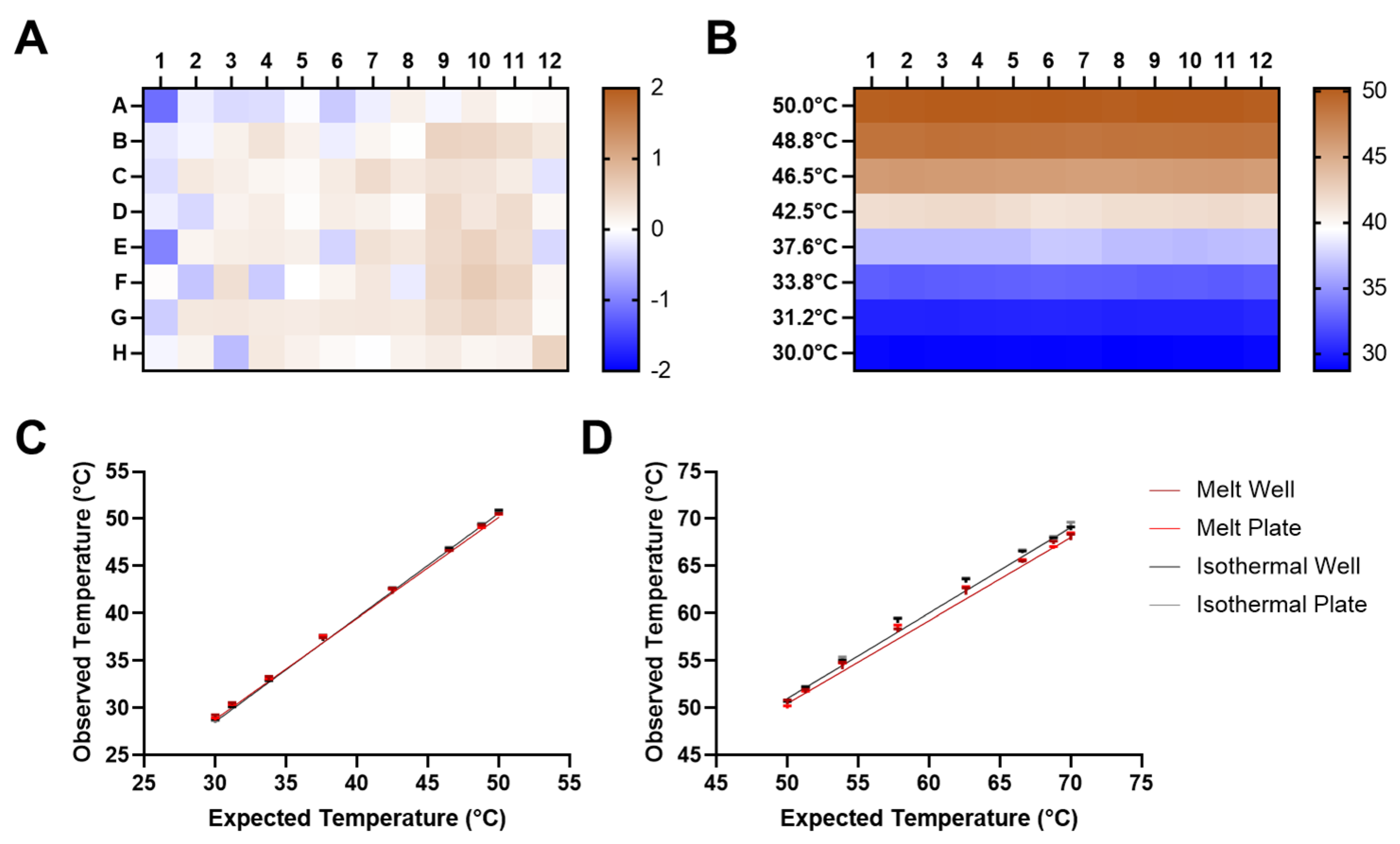
2.5. Temperature and Gradient Uniformity
2.6. Intermachine Variability
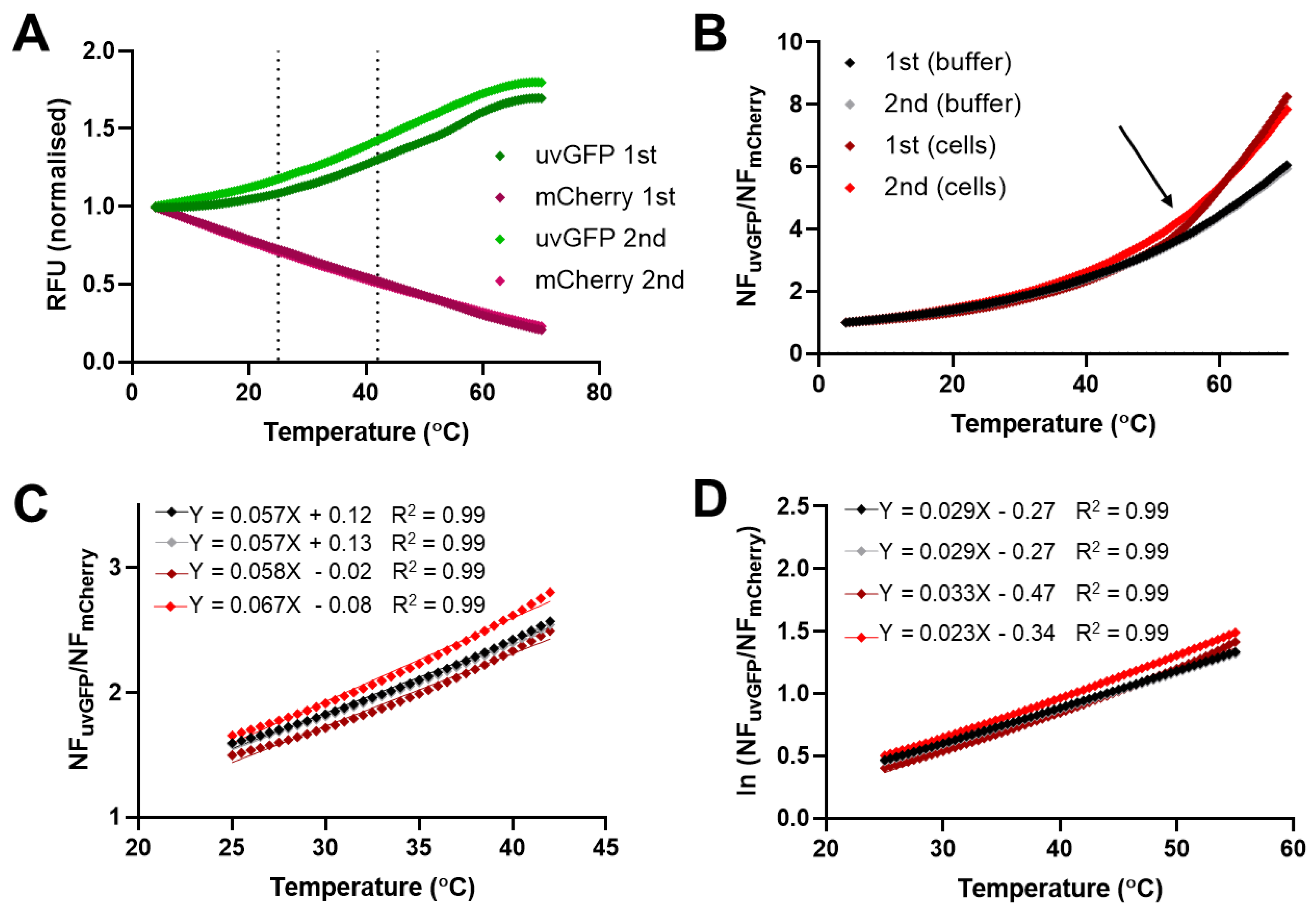
2.7. In Vivo Temperature Sensing in Mixed Bacteria-Encapsulated Format
2.8. Minimum DFPTB Concentration and Detection of Volume Discrepancy
2.9. Absorbance and Fluorescence Spectra
3. Results and Discussion
4. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, H.; Zhang, H.; Xu, Y.; Lassakova, S.; Korabecna, M.; Neuzil, P. PCR past, present and future. Biotechniques 2020, 69, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Kunze, M.; Lattermann, C.; Diederichs, S.; Kroutil, W.; Buchs, J. Minireactor-based high-throughput temperature profiling for the optimization of microbial and enzymatic processes. J. Biol. Eng. 2014, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Zhang, Z.; Grubner, S.; Yurkovich, J.T.; Palsson, B.O.; Zielinski, D.C. Temperature-Dependent Estimation of Gibbs Energies Using an Updated Group-Contribution Method. Biophys. J. 2018, 114, 2691–2702. [Google Scholar] [CrossRef] [Green Version]
- Cianciulli, C.; Watzig, H. Infrared-based temperature measurements in capillary electrophoresis. Electrophoresis 2011, 32, 1530–1536. [Google Scholar] [CrossRef]
- Lao, A.I.K.; Lee, T.M.H.; Hsing, I.; Ip, N.Y. Precise temperature control of microfluidic chamber for gas and liquid phase reactions. Sens. Actuators A 2000, 84, 11–17. [Google Scholar] [CrossRef]
- McKenzie, B.A.; Grover, W.H. A microfluidic thermometer: Precise temperature measurements in microliter- and nanoliter-scale volumes. PLoS ONE 2017, 12, e0189430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, P.; Moghadam, T.T.; Ranjbar, B. Differential scanning calorimetry techniques: Applications in biology and nanoscience. J. Biomol. Tech. 2010, 21, 167–193. [Google Scholar]
- Wong, F.H.; Banks, D.S.; Abu-Arish, A.; Fradin, C. A molecular thermometer based on fluorescent protein blinking. J. Am. Chem. Soc. 2007, 129, 10302–10303. [Google Scholar] [CrossRef]
- Deepankumar, K.; Nadarajan, S.P.; Bae, D.H.; Baek, K.H.; Choi, K.Y.; Yun, H. Temperature sensing using red fluorescent protein. Biotechnol. Bioprocess Eng. 2015, 20, 67–72. [Google Scholar] [CrossRef]
- Savchuk, O.A.; Silvestre, O.F.; Adao, R.M.R.; Nieder, J.B. GFP fluorescence peak fraction analysis based nanothermometer for the assessment of exothermal mitochondria activity in live cells. Sci. Rep. 2019, 9, 7535. [Google Scholar] [CrossRef] [Green Version]
- Ogle, M.M.; McWilliams, A.D.S.; Jiang, B.; Marti, A.A. Latest trends in temperature sensing by molecular probes. ChemPhotoChem 2020, 4, 255–270. [Google Scholar] [CrossRef]
- Zhegalova, N.G.; Aydt, A.; Wang, S.T.; Berezin, M.Y. Molecular thermometers for potential applications in thermal ablation procedures. In Reporters, Markers, Dyes, Nanoparticles, and Molecular Probes for Biomedical Applications; Society of Photo-Optical Instrumentation Engineers: San Francisco, CA, USA, 2013. [Google Scholar]
- Homma, M.; Takei, Y.; Murata, A.; Inoue, T.; Takeoka, S. A ratiometric fluorescent molecular probe for visualisation of mitochondrial temperature in living cells. Chem. Commun. 2015, 51, 6194–6197. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Arai, Y.; Kotera, I.; Okabe, K.; Kamei, Y.; Nagai, T. Genetically encoded ratiometric fluorescent thermometer with wide range and rapid response. PLoS ONE 2017, 12, e0172344. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Wazawa, T.; Sakamoto, J.; Vu, C.Q.; Nakano, M.; Kamei, Y.; Nagai, T. Intracellular Heat Transfer and Thermal Property Revealed by Kilohertz Temperature Imaging with a Genetically Encoded Nanothermometer. Nano Lett. 2022, 22, 5698–5707. [Google Scholar] [CrossRef]
- Feng, G.; Zhang, H.; Zhu, X.; Zhang, J.; Fang, J. Fluorescence thermometers: Intermediation of fundamental temperature and light. Biomater. Sci. 2022, 10, 1855–1882. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wang, Y.; Wu, X.; Chen, M.; Mu, L.; She, G.; Shi, W. An ultrasensitive ratiometric fluorescent thermometer based on frustrated static excimers in the physiological temperature range. Chem. Commun. 2019, 55, 3509–3512. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Z.; Wu, L.; Lu, S.; Ling, S.; Li, G.; Xu, L.; Ma, L.; Hou, Y.; Wang, X.; et al. Emissive Platinum(II) Cages with Reverse Fluorescence Resonance Energy Transfer for Multiple Sensing. J. Am. Chem. Soc. 2020, 142, 2592–2600. [Google Scholar] [CrossRef]
- Tang, S.; Wang, N.; Xu, X.; Feng, S. A ratiometric fluorescent thermometer based on amphiphilic alkynylpyrene derivatives. N. J. Chem. 2019, 43, 6461–6464. [Google Scholar] [CrossRef]
- Shi, L.; Song, W.; Lian, C.; Chen, W.; Mei, J.; Su, J.; Liu, H.; Tian, H. Dual-emitting dihydrophenazines for highly sensitive and ratiometric thermometry over a wide temperature range. Anvanced Opt. Mater. 2018, 6, 1800190. [Google Scholar] [CrossRef]
- Song, W.; Ye, W.; Shi, L.; Huang, J.; Zhang, Z.; Mei, J.; Su, J.; Tian, H. Smart molecular butterfly: An ultrasensitive and range-tunable ratiometric thermometer based on dihydrophenazines. Mater. Horiz. 2020, 7, 615–623. [Google Scholar] [CrossRef]
- Lv, Y.; Jin, Y.; Wu, H.; Liu, D.; Xiong, G.; Ju, G.; Chen, L.; Hu, Y. An all-optical ratiometric thermometer based on reverse thermal response from interplay among diverse emission center and traps with high-temperature sensitivity. Ind. Eng. Chem. Res. 2019, 58, 21242–21251. [Google Scholar] [CrossRef]
- Moreau, M.J.; Morin, I.; Schaeffer, P.M. Quantitative determination of protein stability and ligand binding using a green fluorescent protein reporter system. Mol. Biosyst. 2010, 6, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Moreau, M.J.J.; Morin, I.; Askin, S.; Cooper, A.E.; Moreland, N.J.; Vasudevan, S.G.; Schaeffer, P.M. Rapid determination of protein stability and ligand binding by differential scanning fluorimetry of GFP-tagged proteins. RSC Adv. 2012, 2, 11892–11900. [Google Scholar] [CrossRef]
- Sanford, L.N.; Wittwer, C.T. Monitoring temperature with fluorescence during real-time PCR and melting analysis. Anal. Biochem. 2013, 434, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Crews, N.; Ameel, T.; Wittwer, C.; Gale, B. Flow-induced thermal effects on spatial DNA melting. Lab Chip 2008, 8, 1922–1929. [Google Scholar] [CrossRef]
- Erali, M.; Voelkerding, K.V.; Wittwer, C.T. High resolution melting applications for clinical laboratory medicine. Exp. Mol. Pathol. 2008, 85, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Oerlemans, R.; Groves, M.R. Theory and applications of differential scanning fluorimetry in early-stage drug discovery. Biophys. Rev. 2020, 12, 85–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorenson, A.E.; Schaeffer, P.M. High-Throughput Differential Scanning Fluorimetry of GFP-Tagged Proteins. Methods Mol. Biol. 2020, 2089, 69–85. [Google Scholar] [CrossRef]
- Nagy, A.; Malnasi-Csizmadia, A.; Somogyi, B.; Lorinczy, D. Thermal stability of chemically denatured green fluorescent protein (GFP): A preliminary study. Thermochim. Acta 2004, 410, 161–163. [Google Scholar] [CrossRef]
- Rana, M.S.; Wang, X.; Banerjee, A. An Improved Strategy for Fluorescent Tagging of Membrane Proteins for Overexpression and Purification in Mammalian Cells. Biochemistry 2018, 57, 6741–6751. [Google Scholar] [CrossRef] [Green Version]
- Span, M.; Verblakt, M.; Hendrikx, T. Comparison of temperature dynamics of various thermal cycler calibration methods. In Proceedings of the 19th International Congress of Metrology, Paris, France, 24–26 September 2019; p. 5. [Google Scholar]
- Wood, J.M. Osmosensing by bacteria: Signals and membrane-based sensors. Microbiol. Mol. Biol. Rev. 1999, 63, 230–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleja, N.; Manzoor, O.; Khan, I.; Ahmad, A.; Mohsin, M. Role of green fluorescent proteins and their variants in development of FRET-based sensors. J. Biosci. 2018, 43, 763–784. [Google Scholar] [CrossRef] [PubMed]
- Bajar, B.T.; Wang, E.S.; Zhang, S.; Lin, M.Z.; Chu, J. A Guide to Fluorescent Protein FRET Pairs. Sensors 2016, 16, 1488. [Google Scholar] [CrossRef] [PubMed]
- Spicer, G.; Efeyan, A.; Adam, A.P.; Thompson, S.A. Universal guidelines for the conversion of proteins and dyes into functional nanothermometers. J. Biophotonics 2019, 12, e201900044. [Google Scholar] [CrossRef] [PubMed]
- Maksimov, E.G.; Yaroshevich, I.A.; Tsoraev, G.V.; Sluchanko, N.N.; Slutskaya, E.A.; Shamborant, O.G.; Bobik, T.V.; Friedrich, T.; Stepanov, A.V. A genetically encoded fluorescent temperature sensor derived from the photoactive Orange Carotenoid Protein. Sci. Rep. 2019, 9, 8937. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.; Kim, Y.H.; Byun, J.Y.; Park, S.R. Use of multiplex polymerase chain reactions to indicate the accuracy of the annealing temperature of thermal cycling. Anal. Biochem. 2005, 338, 192–200. [Google Scholar] [CrossRef]
- Ye, F.; Wu, C.; Jin, Y.; Chan, Y.-H.; Zhang, X.; Chiu, D.T. Ratiometric temperature sensing with semiconducting polymer dots. J. Am. Chem. Soc. 2011, 133, 8146–8149. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.-H.; Sun, Y.; Gong, Z.-L.; Li, Z.-Y.; Zhou, Z.; Wang, H.; Li, X.; Saha, M.L.; Zhong, Y.-W.; Stang, P.J. Temperature-Responsive Fluorescent Organoplatinum(II) Metallacycles. J. Am. Chem. Soc. 2018, 140, 7723–7729. [Google Scholar] [CrossRef]
- Cui, Y.; Song, R.; Yu, J.; Liu, M.; Wang, Z.; Wu, C.; Yang, Y.; Wang, Z.; Chen, B.; Qian, G. Dual-emitting MOF supersetdye composite for ratiometric temperature sensing. Adv. Mater. 2015, 27, 1420–1425. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Buffer | pH | (NFuvGFP/NFmCherry) | ln(NFuvGFP/NFmCherry) | ||
|---|---|---|---|---|---|
| Range (°C) | Absolute Sensitivity (%°C−1) | Range (°C) | Absolute Sensitivity (%°C−1) | ||
| HS phosphate ## | 7.8 | 25–42 | 6.7 | 4–70 | 3.0 |
| Phosphate # | 7.8 | 25–42 | 6.0 | 4–75 | 2.8 |
| PBS | 7.2 | 25–42 | 5.7 | 4–70 | 2.7 |
| PBS 1% BSA | 7.2 | 25–42 | 6.0 | 4–75 | 2.6 |
| PBS-T 4% BSA | 7.2 | 25–42 | 5.6 | 4–75 | 2.6 |
| HEPES * | 7.5 | 25–42 | 5.2 | 4–70 | 2.6 |
| Phosphate * | 7.8 | 25–42 | 6.1 | 4–70 | 2.8 |
| Phosphate | 9.1 | 25–42 | 4.8 | 4–70 | 2.4 |
| Phosphate | 6.9 | 25–42 | 5.4 | 4–70 | 2.6 |
| Phosphate | 5.4 | 25–42 | 5.4 | 4–65 | 2.6 |
| Phosphate | 4.8 | 25–42 | 4.8 | 4–45 | 2.3 |
| Citrate * | 6.0 | 25–42 | 6.0 | 4–70 | 2.7 |
| Bis-Tris * | 6.0 | 25–42 | 5.4 | 4–60 | 2.5 |
| Ammonium sulfate * | 5.7 | 25–42 | 5.4 | 4–42 | 2.5 |
| HS Phosphate ##^ | 7.8 | 25–42 | 5.9 | 4–55 | 2.8 |
| Phosphate #^ | 7.8 | 25–42 | 6.0 | 4–55 | 2.9 |
| PBS ^ | 7.2 | 25–42 | 5.9 | 4–55 | 2.8 |
| Phosphate ^ | 4.8 | 25–42 | 4.9 | 4–45 | 2.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorenson, A.E.; Schaeffer, P.M. Real-Time Temperature Sensing Using a Ratiometric Dual Fluorescent Protein Biosensor. Biosensors 2023, 13, 338. https://doi.org/10.3390/bios13030338
Sorenson AE, Schaeffer PM. Real-Time Temperature Sensing Using a Ratiometric Dual Fluorescent Protein Biosensor. Biosensors. 2023; 13(3):338. https://doi.org/10.3390/bios13030338
Chicago/Turabian StyleSorenson, Alanna E., and Patrick M. Schaeffer. 2023. "Real-Time Temperature Sensing Using a Ratiometric Dual Fluorescent Protein Biosensor" Biosensors 13, no. 3: 338. https://doi.org/10.3390/bios13030338