
The Mechanism of Manganese Ferrite Nanomaterials Promoting Drought Resistance in Rice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. MnFe2O4 NM Synthesis and Characterization
2.2. Plant Cultivation and NM Exposure
2.3. Root Morphology, Photosynthesis, Element Content, and Single Particle Concentration
2.4. Determination of IAA and ABA
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. MnFe2O4 NM Characterization
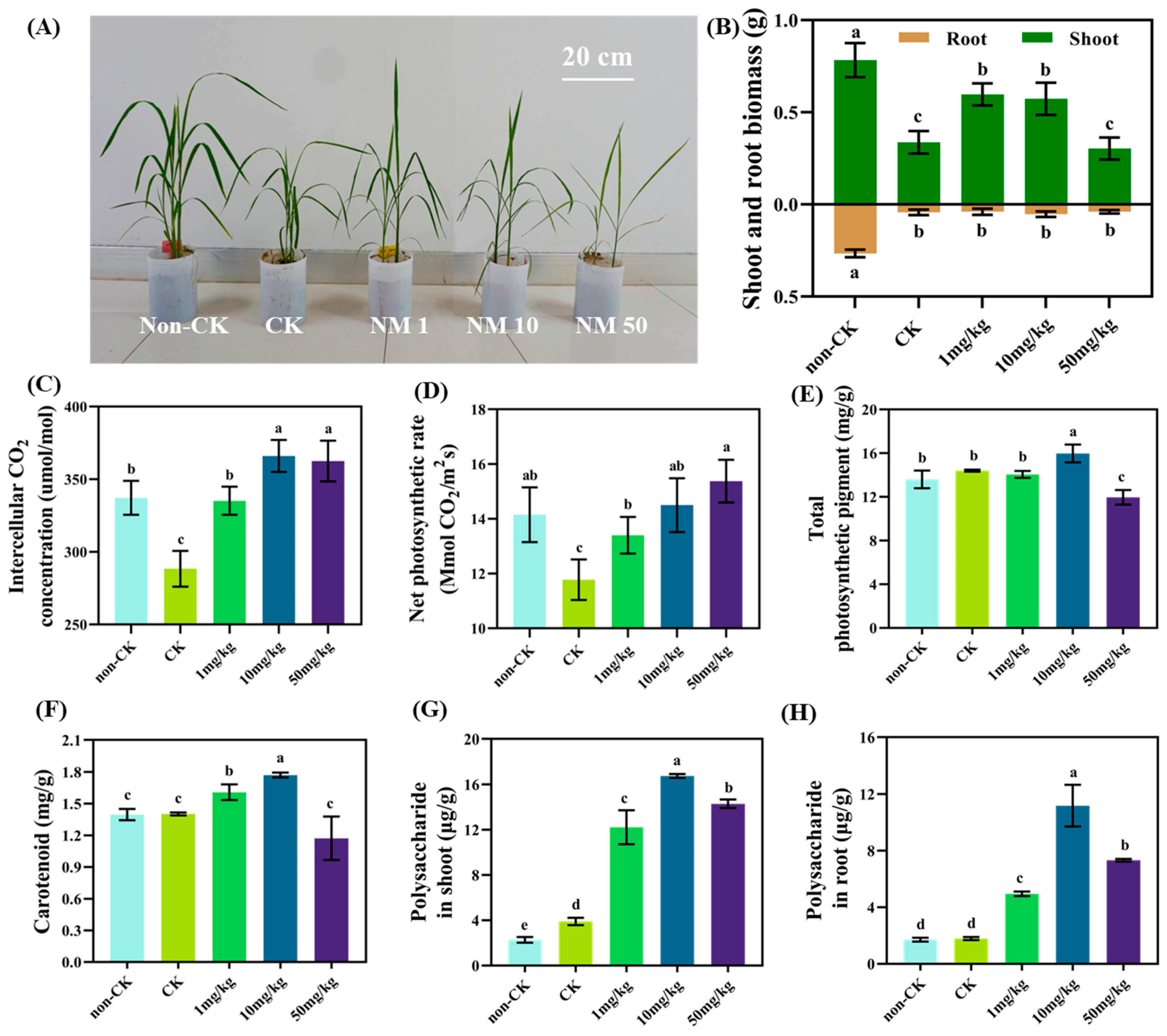
3.2. MnFe2O4 NMs Alleviated the Adverse Effects of Drought on Rice Growth
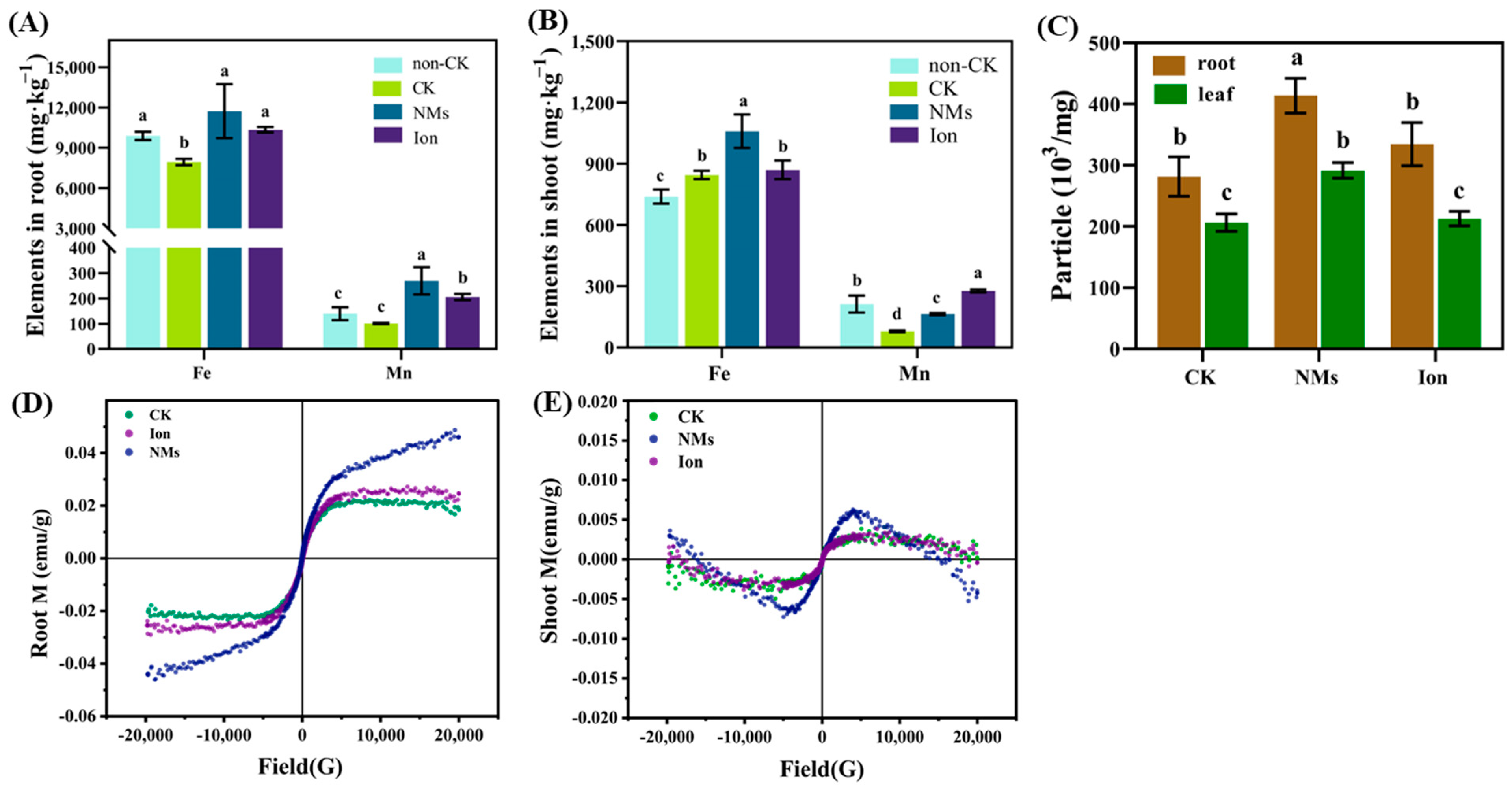
3.3. Accumulation of MnFe2O4 NMs in Rice Plants under Drought Stress
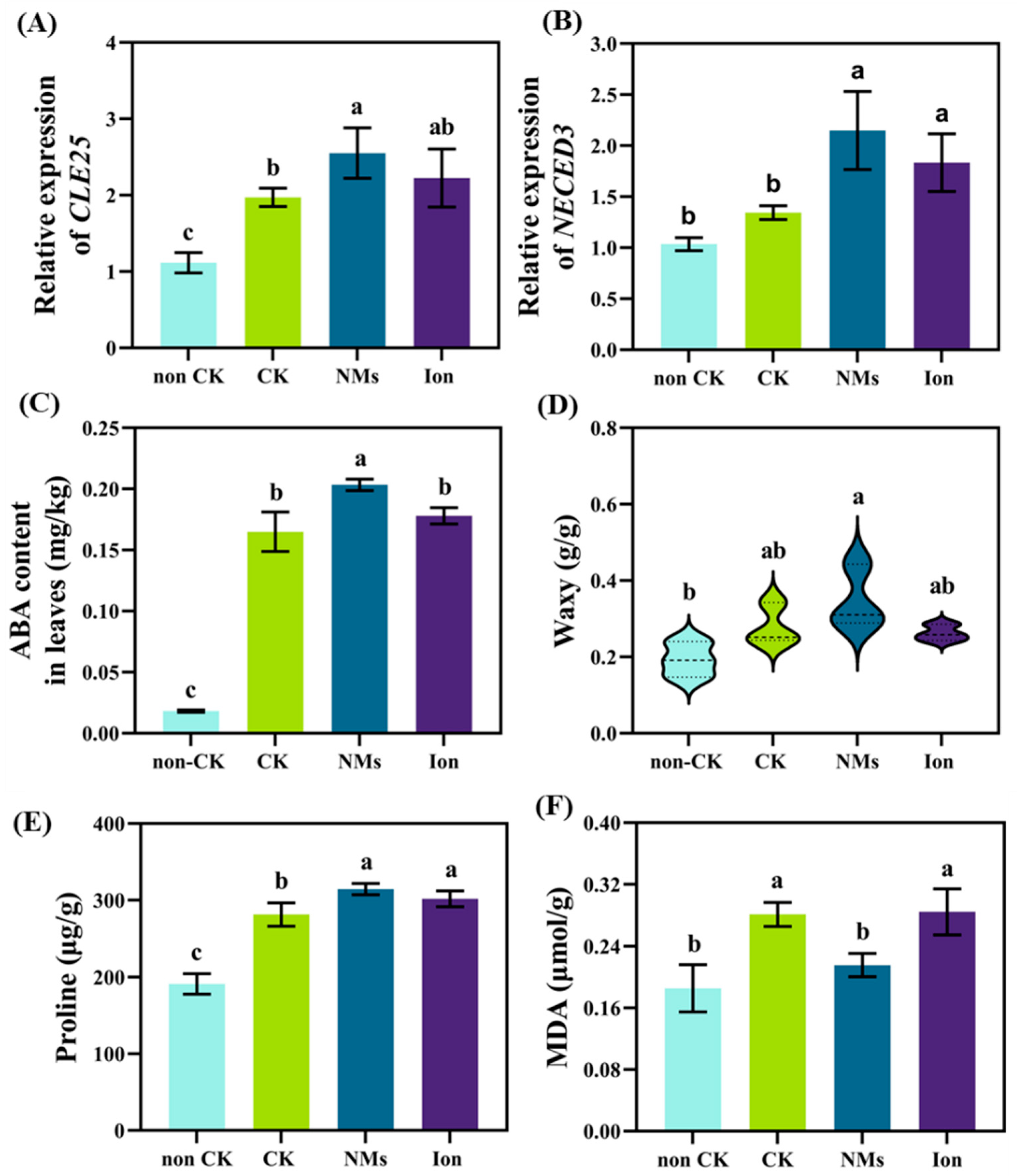
3.4. MnFe2O4 NMs Enhanced Signal Transduction in Rice Roots
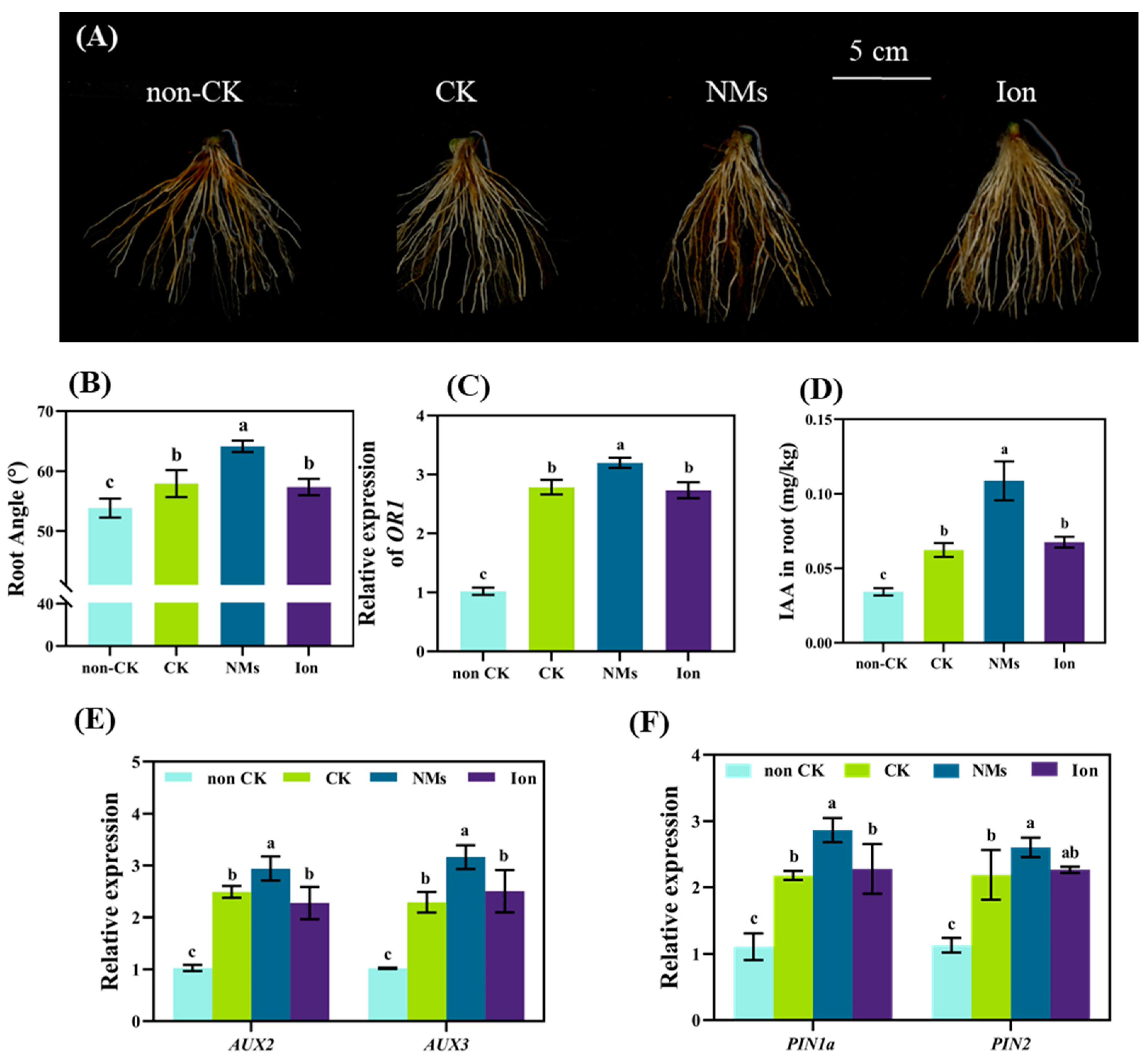
3.5. MnFe2O4 NMs Changed Root Development under Drought Stress
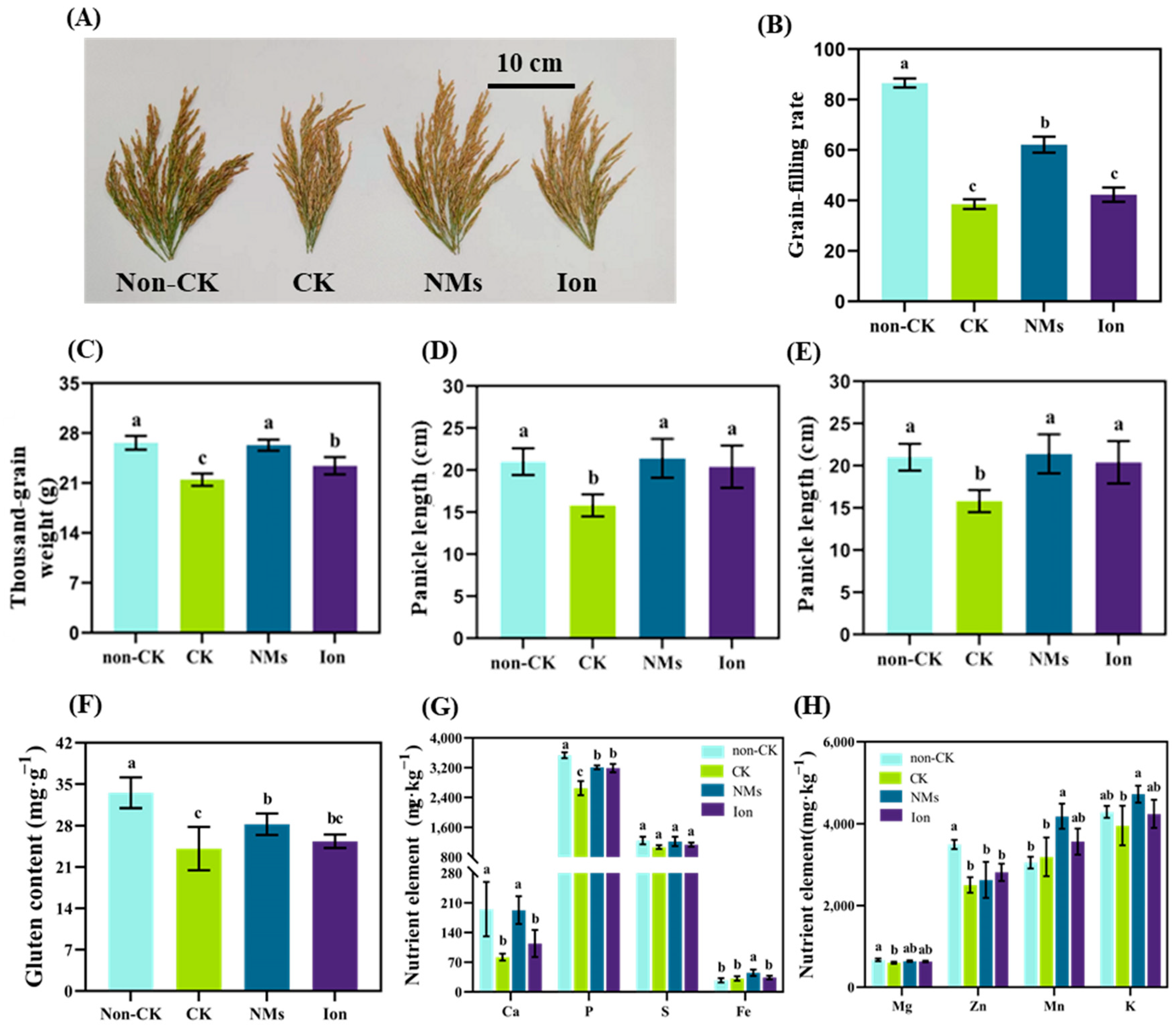
3.6. MnFe2O4 NMs Enhanced the Grain Yield and Quality of Rice under Drought Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Adam, D. How far will global population rise? Researchers can’t agree. Nature 2021, 597, 462–465. [Google Scholar] [CrossRef]
- Challinor, A.J.; Watson, J.; Lobell, D.B.; Howden, S.M.; Smith, D.R.; Chhetri, N. A meta-analysis of crop yield under climate change and adaptation. Nat. Clim. Chang. 2014, 4, 287–291. [Google Scholar] [CrossRef]
- OECD and Food and Agriculture Organization of the United Nations. OECD-FAO Agricultural Outlook 2019-2028; OECD Publishing: Paris, France, 2019. [Google Scholar] [CrossRef]
- Challabathula, D.; Zhang, Q.; Bartels, D. Protection of photosynthesis in desiccation-tolerant resurrection plants. J. Plant Physiol. 2018, 227, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Zu, X.; Lu, Y.; Wang, Q.; La, Y.; Hong, X.; Tan, F.; Niu, J.; Xia, H.; Wu, Y.; Zhou, S.; et al. Increased drought resistance 1 mutation increases drought tolerance of upland rice by altering physiological and morphological traits and limiting ROS levels. Plant Cell Physiol. 2021, 62, 1168–1184. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef]
- Quan, W.; Liu, X.; Wang, H.; Chan, Z. Comparative physiological and transcriptional analyses of two contrasting drought tolerant alfalfa varieties. Front. Plant Sci. 2015, 6, 1256. [Google Scholar] [CrossRef]
- Bristiel, P.; Roumet, C.; Violle, C.; Volaire, F. Coping with drought: Root trait variability within the perennial grass Dactylis glomerata captures a trade-off between dehydration avoidance and dehydration tolerance. Plant Soil 2018, 434, 327–342. [Google Scholar] [CrossRef]
- Sandor, R.; Der, C.; Grosjean, K.; Anca, I.; Noirot, E.; Leborgne-Castel, N.; Gerbeau-Pissot, P. Plasma membrane order and fluidity are diversely triggered by elicitors of plant defence. J. Exp. Bot. 2016, 67, 5173–5185. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 2018, 556, 235–238. [Google Scholar] [CrossRef]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Uga, Y.; Kitomi, Y.; Yamamoto, E.; Kanno, N.; Kawai, S.; Mizubayashi, T.; Fukuoka, S. A QTL for root growth angle on rice chromosome 7 is involved in the genetic pathway of DEEPER ROOTING 1. Rice 2015, 8, 8. [Google Scholar] [CrossRef]
- Kitomi, Y.; Hanzawa, E.; Kuya, N.; Inoue, H.; Hara, N.; Kawai, S.; Kanno, N.; Endo, M.; Sugimoto, K.; Yamazaki, T.; et al. Root angle modifications by the DRO1 homolog improve rice yields in saline paddy fields. Proc. Natl. Acad. Sci. USA 2020, 117, 21242–21250. [Google Scholar] [CrossRef]
- Sisi, N.A.; Ruzicka, K. ER-Localized PIN carriers: Regulators of intracellular auxin homeostasis. Plants 2020, 9, 11. [Google Scholar]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A critical evaluation of nanopesticides and nanofertilizers against their conventional analogues. Nat. Nanotechnol. 2018, 13, 7–684. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Yue, L.; Cao, X.; Chen, F.; Li, J.; Zhang, J.; Xing, B. Carbon dots promoted soybean photosynthesis and amino acid biosynthesis under drought stress: Reactive oxygen species scavenging and nitrogen metabolism. Sci. Total Environ. 2023, 856, 159125. [Google Scholar] [CrossRef]
- Kim, J.-H.; Lee, Y.; Kim, E.-J.; Gu, S.; Sohn, E.J.; Seo, Y.S.; An, H.J.; Chang, Y.-S. Exposure of iron nanoparticles to Arabidopsis thaliana enhances root elongation by triggering cell wall loosening. Environ. Sci. Technol. 2014, 48, 3477–3485. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; Andrews, J.; Fugice, J.; Singh, U.; Bindraban, P.S.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Facile coating of urea with low-dose ZnO nanoparticles promotes wheat performance and enhances Zn uptake under drought stress. Front. Plant Sci. 2020, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Crop Nutrition. Manganese in Crop Production. Crop Nutririon Resource Library. Available online: https://www.cropnutrition.com/resource-library/manganese-in-crop-production (accessed on 1 April 2023).
- Yue, L.; Feng, Y.; Ma, C.; Wang, C.; Chen, F.; Cao, X.; Wang, J.; White, J.C.; Wang, Z.; Xing, B. Molecular mechanisms of early flowering in tomatoes induced by manganese ferrite (MnFe2O4) nanomaterials. ACS Nano 2022, 16, 5636–5646. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Mohammadi, H.; Kariman, K. Nanosilicon-based recovery of barley (Hordeum vulgare) plants subjected to drought stress. Environ. Sci. Nano 2020, 7, 443–461. [Google Scholar] [CrossRef]
- Rigon, J.P.G.; Capuani, S.; Beltrão, N.D.E.M.; Brito Neto, J.F.D.; Sofiatti, V.; França, F.V.D. Non-destructive determination of photosynthetic pigments in the leaves of castor oil plants. Acta Scientiarum. Agron. 2012, 34, 325–329. [Google Scholar] [CrossRef]
- Tombuloglu, H.; Tombuloglu, G.; Slimani, Y.; Ercan, I.; Sozeri, H.; Baykal, A. Impact of manganese ferrite (MnFe2O4) nanoparticles on growth and magnetic character of barley (Hordeum vulgare L.). Environ. Pollut. 2018, 243, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M.U. Does enhanced photosynthesis enhance growth? Lessons learned from CO2 enrichment studies. Plant Physiol. 2011, 155, 117–124. [Google Scholar] [CrossRef]
- Bouranis, D.L.; Malagoli, M.; Avice, J.C.; Bloem, E. Advances in plant sulfur research. Plants 2020, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Rahman, M.M.; Ghosh, T.K.; Kabir, A.H.; Abdelrahman, M.; Khan, A.R.; Mochida, K.; Tran, L.-S.P. Potassium in plant physiological adaptation to abiotic stresses. Plant Physiol. Biochem. 2022, 186, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ma, X.; Zhang, J.; Zhu, J.; Lu, T.; Wang, Q.; Wang, X.; Hua, W.; Xu, S. Meta-analysis of the role of zinc in coordinating absorption of mineral elements in wheat seedlings. Plant Methods 2021, 17, 105. [Google Scholar] [CrossRef]
- Zhang, X.; Cheng, X.; Zhang, Q. Nanostructured energy materials for electrochemical energy conversion and storage: A review. J. Energy Chem. 2016, 25, 967–984. [Google Scholar] [CrossRef]
- Shankramma, K.; Yallappa, S.; Shivanna, M.B.; Manjanna, J. Fe2O3 magnetic nanoparticles to enhance S. lycopersicum (tomato) plant growth and their biomineralization. Appl. Nanosci. 2015, 6, 983–990. [Google Scholar] [CrossRef]
- Song, X.F.; Hou, X.L.; Liu, C.M. CLE peptides: Critical regulators for stem cell maintenance in plants. Planta 2021, 255, 5. [Google Scholar] [CrossRef]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Martin, L.B.; Romero, P.; Fich, E.A.; Domozych, D.S.; Rose, J.K. Cuticle biosynthesis in tomato leaves is developmentally regulated by abscisic acid. Plant Physiol. 2017, 174, 1384–1398. [Google Scholar] [CrossRef]
- Anupama, A.; Bhugra, S.; Lall, B.; Chaudhury, S.; Chugh, A. Assessing the correlation of genotypic and phenotypic responses of indica rice varieties under drought stress. Plant Physiol. Biochem. 2018, 127, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Ding, C.; Hao, X.; Zeng, J.; Yang, Y.; Wang, X.; Wang, L. CsSWEET1a and CsSWEET17 mediate growth and freezing tolerance by promoting sugar transport across the plasma membrane. Plant Cell Physiol. 2020, 61, 1669–1682. [Google Scholar] [CrossRef]
- Hu, Z.; Tang, Z.; Zhang, Y.; Niu, L.; Yang, F.; Zhang, D.; Hu, Y. Rice SUT and SWEET transporters. Int. J. Mol. Sci. 2021, 22, 1198. [Google Scholar] [CrossRef] [PubMed]
- Camilo, S.; Odindo, A.O.; Kondwakwenda, A.; Sibiya, J. Root traits related with drought and phosphorus tolerance in common bean (Phaseolus vulgaris L.). Agronomy 2021, 11, 552. [Google Scholar] [CrossRef]
- Giri, J.; Bhosale, R.; Huang, G.; Pandey, B.K.; Parker, H.; Zappala, S.; Bennett, M.J. Rice auxin influx carrier OsAUX1 facilitates root hair elongation in response to low external phosphate. Nat. Commun. 2018, 9, 1408. [Google Scholar] [CrossRef]
- Vinarao, R.; Proud, C.; Zhang, X.; Snell, P.; Fukai, S.; Mitchell, J. Stable and novel quantitative trait loci (QTL) confer narrow root cone angle in an aerobic rice (Oryza sativa L.) production system. Rice 2021, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, L.; Xie, B.; Cao, X.; Chen, F.; Wang, C.; Xiao, Z.; Jiao, L.; Wang, Z. The Mechanism of Manganese Ferrite Nanomaterials Promoting Drought Resistance in Rice. Nanomaterials 2023, 13, 1484. https://doi.org/10.3390/nano13091484
Yue L, Xie B, Cao X, Chen F, Wang C, Xiao Z, Jiao L, Wang Z. The Mechanism of Manganese Ferrite Nanomaterials Promoting Drought Resistance in Rice. Nanomaterials. 2023; 13(9):1484. https://doi.org/10.3390/nano13091484
Chicago/Turabian StyleYue, Le, Budiao Xie, Xuesong Cao, Feiran Chen, Chuanxi Wang, Zhenggao Xiao, Liya Jiao, and Zhenyu Wang. 2023. "The Mechanism of Manganese Ferrite Nanomaterials Promoting Drought Resistance in Rice" Nanomaterials 13, no. 9: 1484. https://doi.org/10.3390/nano13091484