
Targeting of Tomato Bushy Stunt Virus with a Genetically Fused C-End Rule Peptide
 , , , and
, , , and 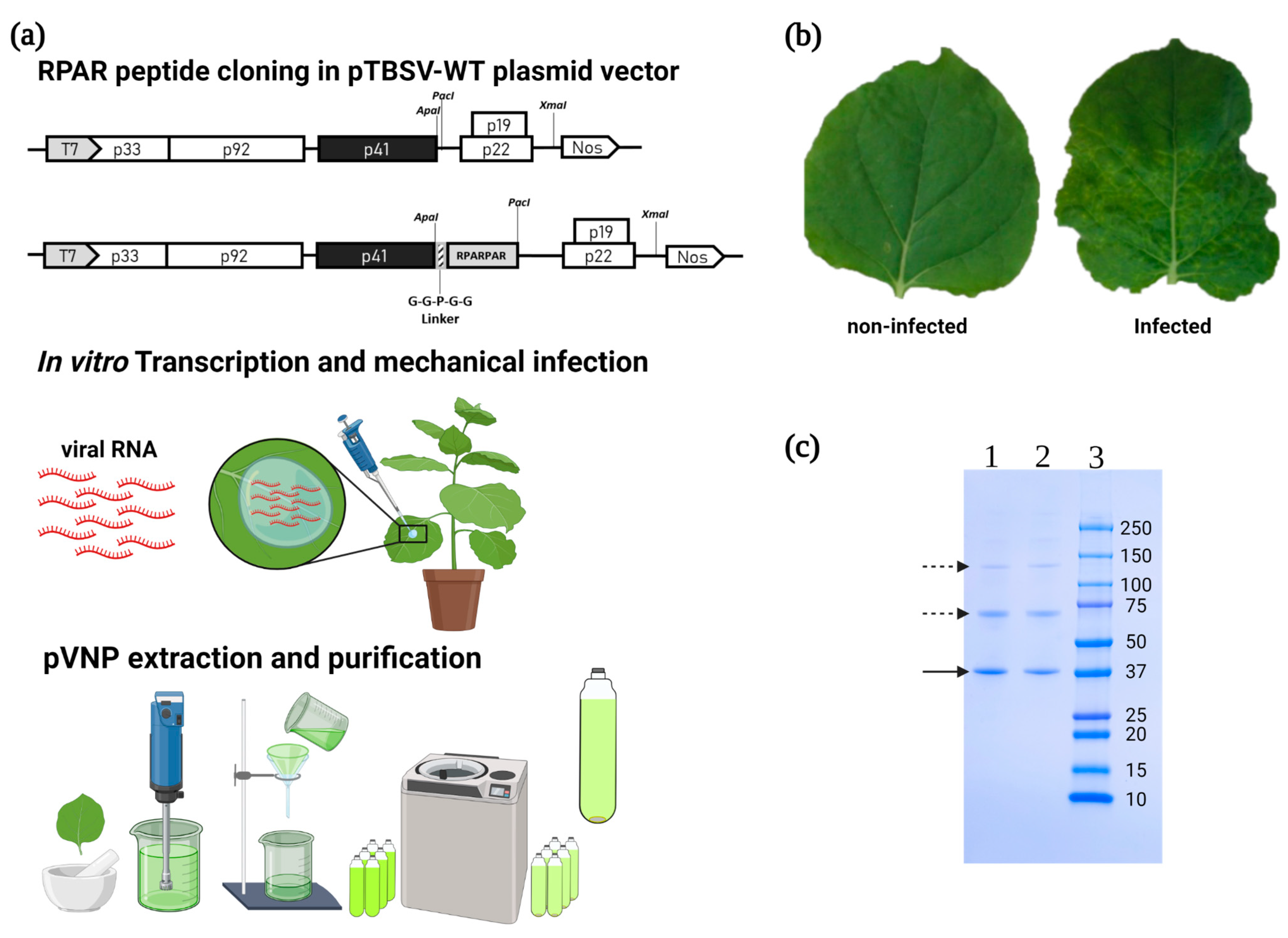{kind=link}
{kind=link}
{kind=link}
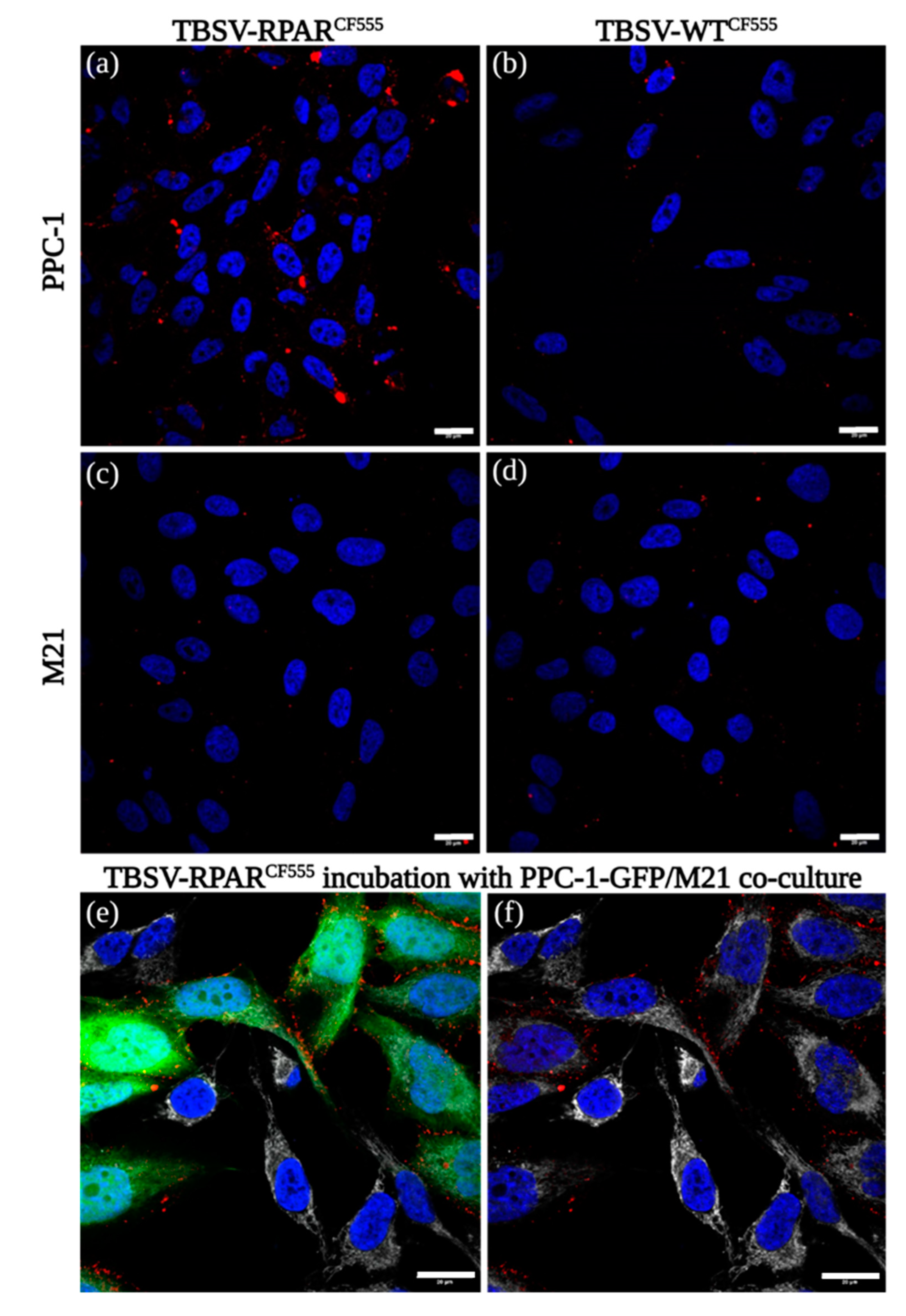{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Engineering of TBSV and Its Production in Plants
2.2. Chemical Conjugation of CF555 Dye on TBSV NPs
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Flow Cytometry
2.5. Confocal Imaging of Cultured Cells
2.6. Doxorubicin Loading of TBSV NPs
2.7. Cell Viability Assay
2.8. In Vivo Biodistribution Analysis of TBSV
2.9. Statistical Analysis
3. Results
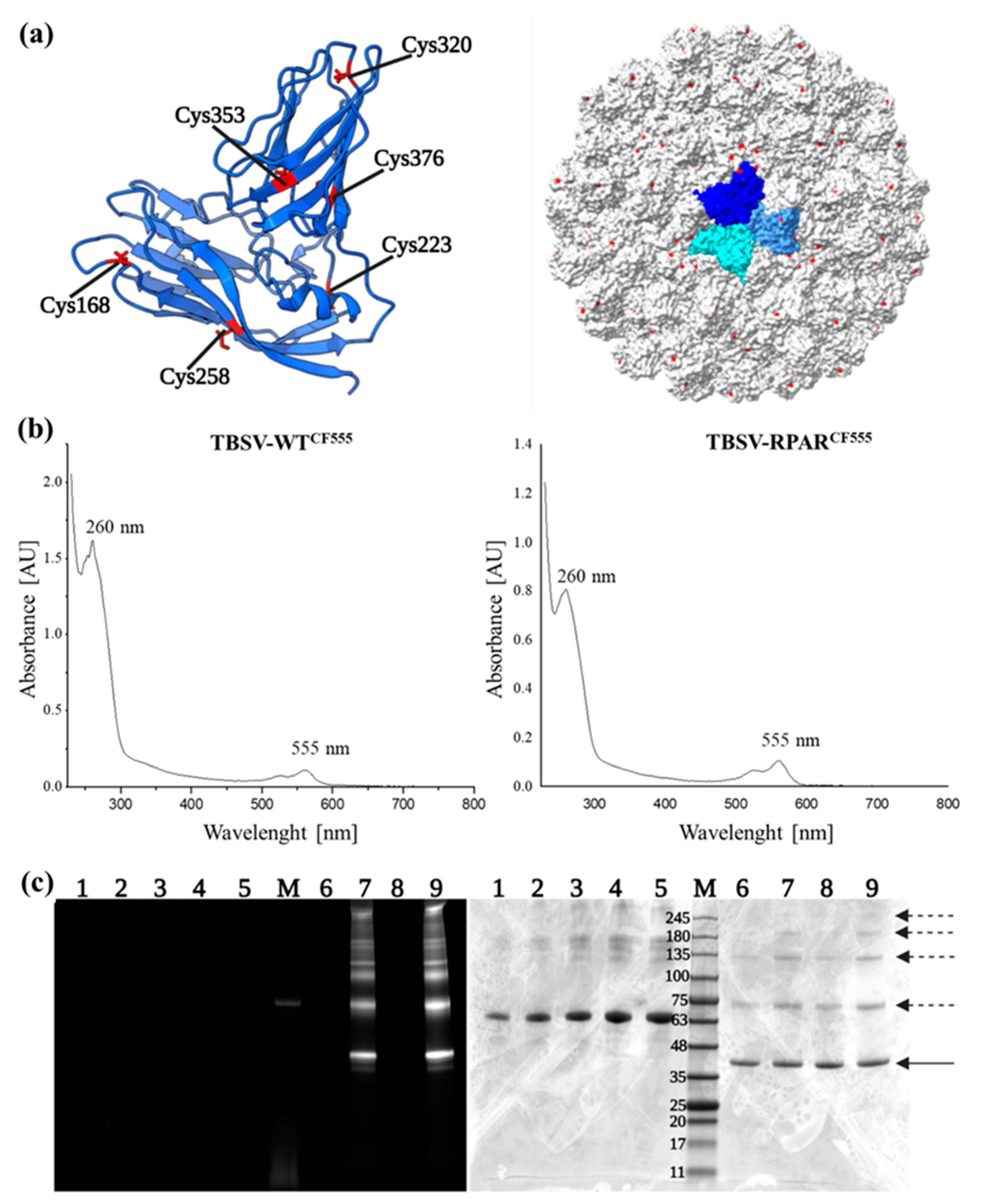
3.1. Construction, Production, and Purification of Wild-Type (WT) and Chimeric TBSV NPs
3.2. Fluorophore Labeling of TBSV NPs
3.3. In Vitro Binding of TBSV-RPAR to NRP-1
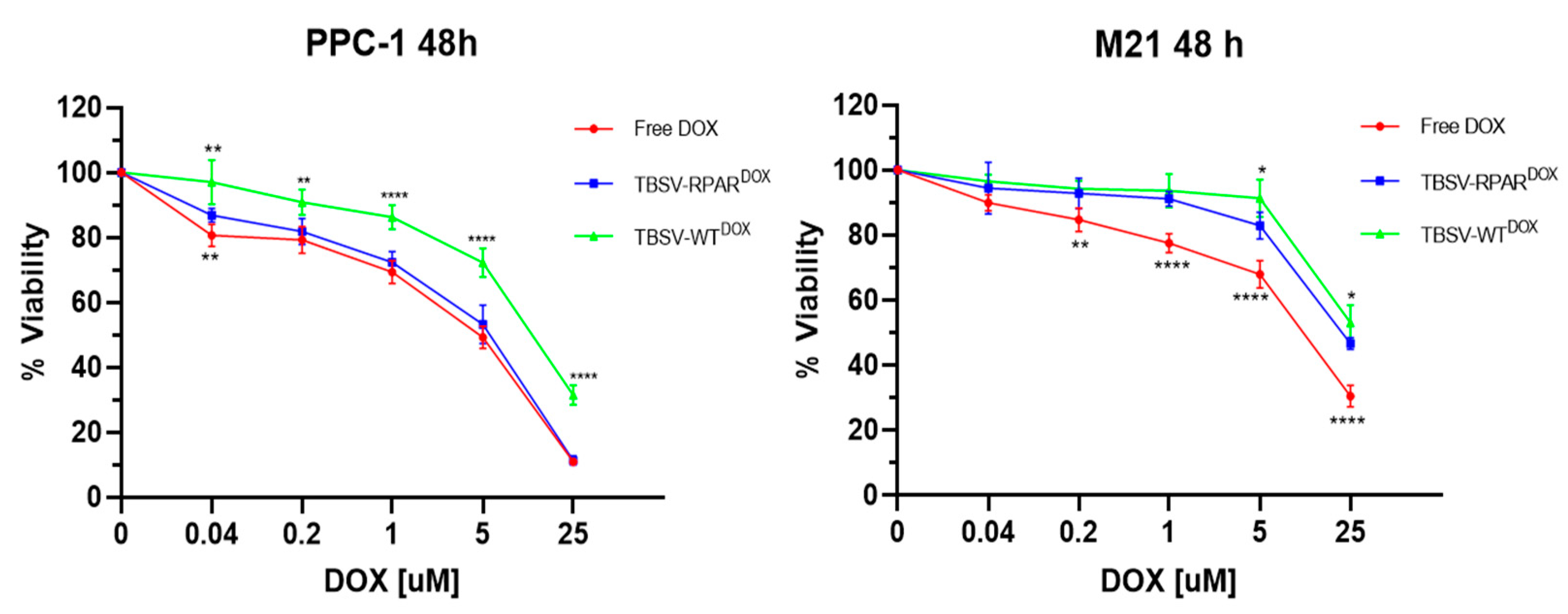
3.4. TBSV-RPAR Loading with DOX and Cytotoxicity Test In Vitro
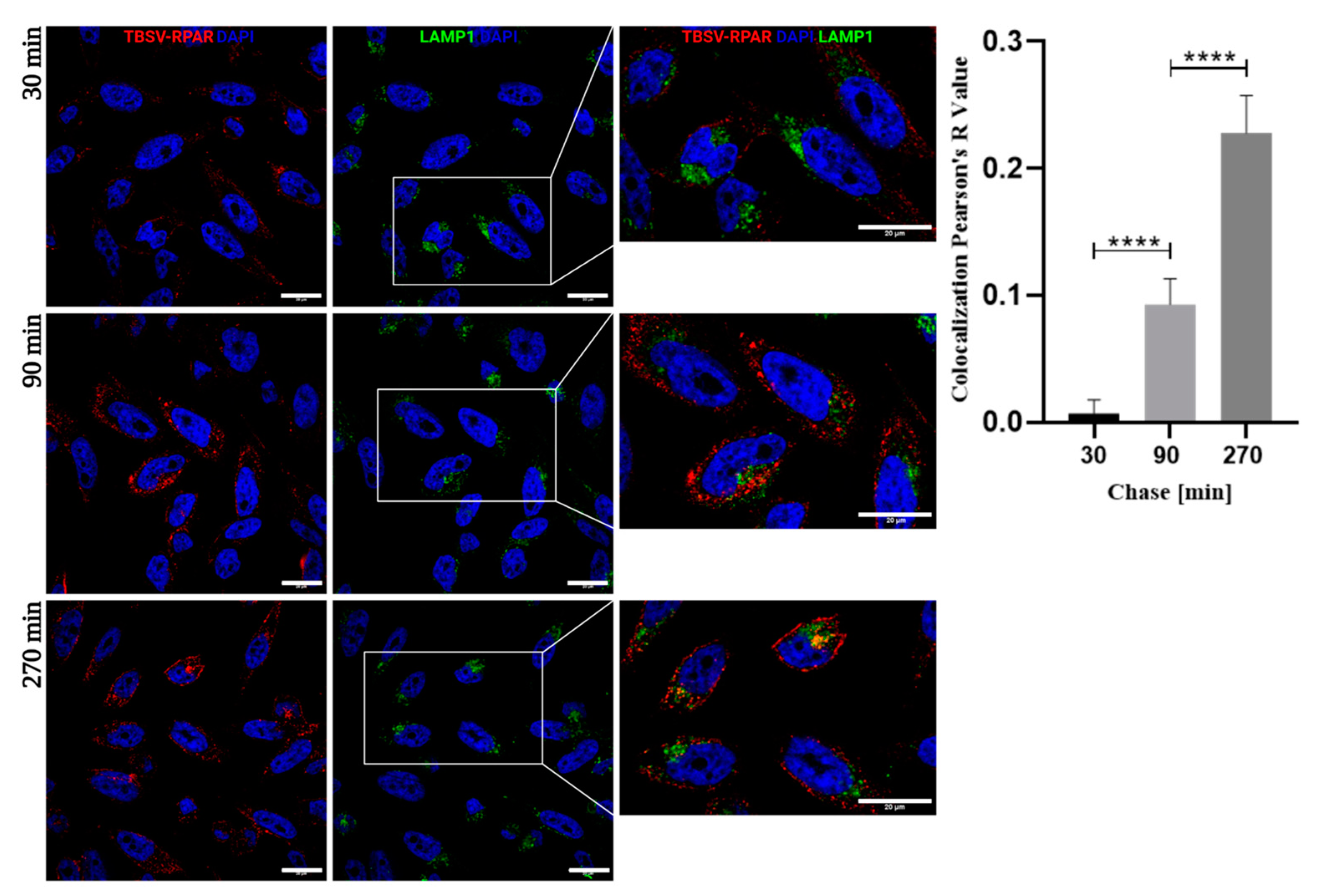
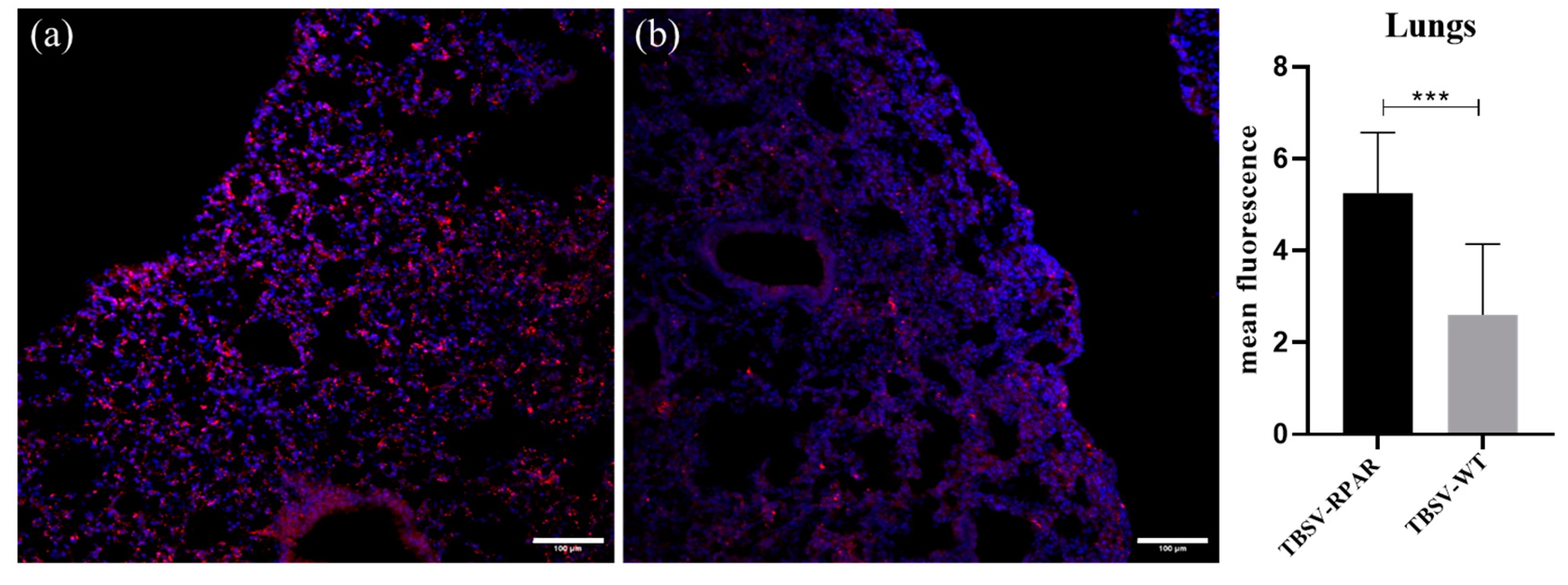
3.5. Systemic TBSV-RPAR NPs Home to Pulmonary Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olshefsky, A.; Richardson, C.; Pun, S.H.; King, N.P. Engineering Self-Assembling Protein Nanoparticles for Therapeutic Delivery. Bioconjug. Chem. 2022, 33, 2018–2034. [Google Scholar] [CrossRef] [PubMed]
- Rohovie, M.J.; Nagasawa, M.; Swartz, J.R. Virus-like Particles: Next-generation Nanoparticles for Targeted Therapeutic Delivery. Bioeng. Transl. Med. 2017, 2, 43. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Nolte, R.J.M.; Cornelissen, J.J.L.M. Virus-Based Nanocarriers for Drug Delivery. Adv. Drug Deliv. Rev. 2012, 64, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Czapar, A.E.; Steinmetz, N.F. Plant Viruses and Bacteriophages for Drug Delivery in Medicine and Biotechnology. Curr. Opin. Chem. Biol. 2017, 38, 108–116. [Google Scholar] [CrossRef]
- Koudelka, K.J.; Pitek, A.S.; Manchester, M.; Steinmetz, N.F. Virus-Based Nanoparticles as Versatile Nanomachines. Annu. Rev. Virol. 2015, 2, 379–401. [Google Scholar] [CrossRef]
- Mateu, M.G. Virus Engineering: Functionalization and Stabilization. Protein Eng. Des. Sel. 2011, 24, 53–63. [Google Scholar] [CrossRef]
- Pokorski, J.K.; Steinmetz, N.F. The Art of Engineering Viral Nanoparticles. Mol. Pharm. 2011, 8, 29–43. [Google Scholar] [CrossRef]
- Sainsbury, F.; Cañizares, M.C.; Lomonossoff, G.P. Cowpea Mosaic Virus: The Plant Virus-Based Biotechnology Workhorse. Annu. Rev. Phytopathol. 2010, 48, 437–455. [Google Scholar] [CrossRef]
- Steinmetz, N.F.; Lin, T.; Lomonossoff, G.P.; Johnson, J.E. Structure-Based Engineering of an Icosahedral Virus for Nanomedicine and Nanotechnology. Curr. Top. Microbiol. Immunol. 2009, 327, 23–58. [Google Scholar]
- Lomonossoff, G.P.; Wege, C. TMV Particles: The Journey From Fundamental Studies to Bionanotechnology Applications. Adv. Virus Res. 2018, 102, 149. [Google Scholar] [CrossRef]
- Chu, S.; Brown, A.D.; Culver, J.N.; Ghodssi, R. Tobacco Mosaic Virus as a Versatile Platform for Molecular Assembly and Device Fabrication. Biotechnol. J. 2018, 13, 1800147. [Google Scholar] [CrossRef] [PubMed]
- Lico, C.; Benvenuto, E.; Baschieri, S. The Two-Faced Potato Virus X: From Plant Pathogen to Smart Nanoparticle. Front. Plant Sci. 2015, 6, 1009. [Google Scholar] [CrossRef] [PubMed]
- Röder, J.; Dickmeis, C.; Commandeur, U. Small, Smaller, Nano: New Applications for Potato Virus X in Nanotechnology. Front. Plant Sci. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, I.; Lee, K.L.; Chen, K.; Shukla, S.; Steinmetz, N.F. Infusion of Imaging and Therapeutic Molecules into the Plant Virus-Based Carrier Cowpea Mosaic Virus: Cargo-Loading and Delivery. J. Control. Release 2013, 172, 568–578. [Google Scholar] [CrossRef]
- Taylor, K.M.; Porta, C.; Lin, T.; Johnson, J.E.; Barker, P.J.; Lomonossoff, G.P. Position-Dependent Processing of Peptides Presented on the Surface of Cowpea Mosaic Virus. Biol. Chem. 1999, 380, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Grasso, S.; Lico, C.; Imperatori, F.; Santi, L. A Plant Derived Multifunctional Tool for Nanobiotechnology Based on Tomato Bushy Stunt Virus. Transgenic Res. 2013, 22, 519–535. [Google Scholar] [CrossRef]
- Lico, C.; Tanno, B.; Marchetti, L.; Novelli, F.; Giardullo, P.; Arcangeli, C.; Pazzaglia, S.; Podda, M.S.; Santi, L.; Bernini, R.; et al. Tomato Bushy Stunt Virus Nanoparticles as a Platform for Drug Delivery to Shh-Dependent Medulloblastoma. Int. J. Mol. Sci. 2021, 22, 10523. [Google Scholar] [CrossRef]
- Harrison, S.C.; Olson, A.J.; Schutt, C.E.; Winkler, F.K.; Bricogne, G. Tomato Bushy Stunt Virus at 2.9 Å Resolution. Nature 1978, 276, 368–373. [Google Scholar] [CrossRef]
- White, K.A.; Nagy, P.D. Advances in the Molecular Biology of Tombusviruses: Gene Expression, Genome Replication, and Recombination. Prog. Nucleic Acid Res. Mol. Biol. 2004, 78, 187–226. [Google Scholar] [CrossRef]
- Olson, A.J.; Bricogne, G.; Harrison, S.C. Structure of Tomato Busy Stunt Virus IV. The Virus Particle at 2.9 A Resolution. J. Mol. Biol. 1983, 171, 61–93. [Google Scholar] [CrossRef]
- Simón-Gracia, L.; Hunt, H.; Teesalu, T. Peritoneal Carcinomatosis Targeting with Tumor Homing Peptides. Mol. A J. Synth. Chem. Nat. Prod. Chem. 2018, 23, 1190. [Google Scholar] [CrossRef]
- Ruoslahti, E. Peptides as Targeting Elements and Tissue Penetration Devices for Nanoparticles. Adv. Mater. 2012, 24, 3747. [Google Scholar] [CrossRef] [PubMed]
- Põšnograjeva, K.; Pleiko, K.; Haugas, M.; Teesalu, T. New Tools for Streamlined In Vivo Homing Peptide Identification. Methods Mol. Biol. 2022, 2383, 385–412. [Google Scholar] [CrossRef] [PubMed]
- Teesalu, T.; Sugahara, K.N.; Ruoslahti, E. Mapping of Vascular ZIP Codes by Phage Display. Methods Enzymol. 2012, 503, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, K.N.; Teesalu, T.; Karmali, P.P.; Kotamraju, V.R.; Agemy, L.; Girard, O.M.; Hanahan, D.; Mattrey, R.F.; Ruoslahti, E. Tissue-Penetrating Delivery of Compounds and Nanoparticles into Tumors. Cancer Cell 2009, 16, 510. [Google Scholar] [CrossRef] [PubMed]
- Teesalu, T.; Sugahara, K.N.; Ruoslahti, E. Tumor-Penetrating Peptides. Front. Oncol. 2013, 3, 216. [Google Scholar] [CrossRef]
- Teesalu, T.; Sugahara, K.N.; Kotamraju, V.R.; Ruoslahti, E. C-End Rule Peptides Mediate Neuropilin-1-Dependent Cell, Vascular, and Tissue Penetration. Proc. Natl. Acad. Sci. USA 2009, 106, 16157–16162. [Google Scholar] [CrossRef]
- Simon-Gracia, L.; Savier, E.; Parizot, C.; Brossas, J.Y.; Loisel, S.; Teesalu, T.; Conti, F.; Charlotte, F.; Scatton, O.; Aoudjehane, L.; et al. Bifunctional Therapeutic Peptides for Targeting Malignant B Cells and Hepatocytes: Proof of Concept in Chronic Lymphocytic Leukemia. Adv. Ther. 2020, 3, 2000131. [Google Scholar] [CrossRef]
- Savier, E.; Simon-Gracia, L.; Charlotte, F.; Tuffery, P.; Teesalu, T.; Scatton, O.; Rebollo, A. Bi-Functional Peptides as a New Therapeutic Tool for Hepatocellular Carcinoma. Pharmaceutics 2021, 13, 1631. [Google Scholar] [CrossRef]
- Graziani, G.; Lacal, P.M. Neuropilin-1 as Therapeutic Target for Malignant Melanoma. Front. Oncol. 2015, 5, 125. [Google Scholar] [CrossRef]
- Jubb, A.M.; Strickland, L.A.; Liu, S.D.; Mak, J.; Schmidt, M.; Koeppen, H. Neuropilin-1 Expression in Cancer and Development. J. Pathol. 2012, 226, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Wonder, E.; Simón-Gracia, L.; Scodeller, P.; Majzoub, R.N.; Kotamraju, V.R.; Ewert, K.K.; Teesalu, T.; Safinya, C.R. Competition of Charge-Mediated and Specific Binding by Peptide-Tagged Cationic Liposome-DNA Nanoparticles in Vitro and in Vivo. Biomaterials 2018, 166, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Tobi, A.; Willmore, A.M.A.; Kilk, K.; Sidorenko, V.; Braun, G.B.; Soomets, U.; Sugahara, K.N.; Ruoslahti, E.; Teesalu, T. Silver Nanocarriers Targeted with a CendR Peptide Potentiate the Cytotoxic Activity of an Anticancer Drug. Adv. Ther. 2021, 4, 2000097. [Google Scholar] [CrossRef]
- Simón-Gracia, L.; Hunt, H.; Scodeller, P.; Gaitzsch, J.; Kotamraju, V.R.; Sugahara, K.N.; Tammik, O.; Ruoslahti, E.; Battaglia, G.; Teesalu, T. IRGD Peptide Conjugation Potentiates Intraperitoneal Tumor Delivery of Paclitaxel with Polymersomes. Biomaterials 2016, 104, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.B.; Braun, G.B.; Ruoslahti, E. Neuropilin-1 and Heparan Sulfate Proteoglycans Cooperate in Cellular Uptake of Nanoparticles Functionalized by Cationic Cell-Penetrating Peptides. Sci. Adv. 2015, 1, e1500821. [Google Scholar] [CrossRef] [PubMed]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Toome, K.; Willmore, A.M.A.; Paiste, P.; Tobi, A.; Sugahara, K.N.; Kirsimäe, K.; Ruoslahti, E.; Braun, G.B.; Teesalu, T. Ratiometric in Vivo Auditioning of Targeted Silver Nanoparticles. Nanoscale 2017, 9, 10094–10100. [Google Scholar] [CrossRef] [PubMed]
- Blandino, A.; Lico, C.; Baschieri, S.; Barberini, L.; Cirotto, C.; Blasi, P.; Santi, L. In Vitro and in Vivo Toxicity Evaluation of Plant Virus Nanocarriers. Colloids Surf. B Biointerfaces 2015, 129, 130–136. [Google Scholar] [CrossRef]
- Lico, C.; Giardullo, P.; Mancuso, M.; Benvenuto, E.; Santi, L.; Baschieri, S. A Biodistribution Study of Two Differently Shaped Plant Virus Nanoparticles Reveals New Peculiar Traits. Colloids Surf. B Biointerfaces 2016, 148, 431–439. [Google Scholar] [CrossRef]
- Milton Harris, J.; Chess, R.B. Effect of Pegylation on Pharmaceuticals. Nat. Rev. Drug Discov. 2003, 2, 214–221. [Google Scholar] [CrossRef]
- Wattendorf, U.; Merkle, H.P. PEGylation as a Tool for the Biomedical Engineering of Surface Modified Microparticles. J. Pharm. Sci. 2008, 97, 4655–4669. [Google Scholar] [CrossRef] [PubMed]
- Rudnick, S.I.; Lou, J.; Shaller, C.C.; Tang, Y.; Klein-Szanto, A.J.P.; Weiner, L.M.; Marks, J.D.; Adams, G.P. Influence of Affinity and Antigen Internalization on the Uptake and Penetration of Anti-HER2 Antibodies in Solid Tumors. Cancer Res. 2011, 71, 2250. [Google Scholar] [CrossRef] [PubMed]
- Diaz Bessone, M.I.; Simón-Gracia, L.; Scodeller, P.; Ramirez, M.D.L.A.; Lago Huvelle, M.A.; Soler-Illia, G.J.A.A.; Simian, M. IRGD-Guided Tamoxifen Polymersomes Inhibit Estrogen Receptor Transcriptional Activity and Decrease the Number of Breast Cancer Cells with Self-Renewing Capacity. J. Nanobiotechnology 2019, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Simón-Gracia, L.; Scodeller, P.; Fuentes, S.S.; Vallejo, V.G.; Ríos, X.; Sebastián, E.S.; Sidorenko, V.; Di Silvio, D.; Suck, M.; De Lorenzi, F.; et al. Application of Polymersomes Engineered to Target P32 Protein for Detection of Small Breast Tumors in Mice. Oncotarget 2018, 9, 18682–18697. [Google Scholar] [CrossRef]
- Hunt, H.; Simón-Gracia, L.; Tobi, A.; Teesalu, T.; Kotamraju, V.R.; Sharma, S.; Sugahara, K.N.; Ruoslahti, E.; Teesalu, T.; Nigul, M.; et al. Targeting of P32 in Peritoneal Carcinomatosis with Intraperitoneal LinTT1 Peptide-Guided pro-Apoptotic Nanoparticles. J. Control. Release 2017, 260, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Säälik, P.; Lingasamy, P.; Toome, K.; Mastandrea, I.; Rousso-Noori, L.; Tobi, A.; Simón-Gracia, L.; Hunt, H.; Paiste, P.; Kotamraju, V.R.; et al. Peptide-Guided Nanoparticles for Glioblastoma Targeting. J. Control. Release 2019, 308, 109–118. [Google Scholar] [CrossRef]
- Simón-Gracia, L.; Sidorenko, V.; Uustare, A.; Ogibalov, I.; Tasa, A.; Tshubrik, O.; Teesalu, T. Novel Anthracycline Utorubicin for Cancer Therapy. Angew. Chem. Int. Ed. Engl. 2021, 60, 17018–17027. [Google Scholar] [CrossRef]
- Zern, B.J.; Chacko, A.M.; Liu, J.; Greineder, C.F.; Blankemeyer, E.R.; Radhakrishnan, R.; Muzykantov, V. Reduction of Nanoparticle Avidity Enhances the Selectivity of Vascular Targeting and PET Detection of Pulmonary Inflammation. ACS Nano 2013, 7, 2461. [Google Scholar] [CrossRef]
- Patel, S.; Kim, J.; Herrera, M.; Mukherjee, A.; Kabanov, A.V.; Sahay, G. Brief Update on Endocytosis of Nanomedicines. Adv. Drug Deliv. Rev. 2019, 144, 90. [Google Scholar] [CrossRef]
- Plummer, E.M.; Manchester, M. Endocytic Uptake Pathways Utilized by CPMV Nanoparticles. Mol. Pharm. 2013, 10, 26–32. [Google Scholar] [CrossRef]
- Masarapu, H.; Patel, B.K.; Chariou, P.L.; Hu, H.; Gulati, N.M.; Carpenter, B.L.; Ghiladi, R.A.; Shukla, S.; Steinmetz, N.F. Physalis Mottle Virus-Like Particles as Nanocarriers for Imaging Reagents and Drugs. Biomacromolecules 2017, 18, 4141–4153. [Google Scholar] [CrossRef]
- Vishnu Vardhan, G.P.; Hema, M.; Sushmitha, C.; Savithri, H.S.; Natraj, U.; Murthy, M.R.N. Development of Sesbania Mosaic Virus Nanoparticles for Imaging. Arch. Virol. 2019, 164, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Simón-Gracia, L.; Hunt, H.; Scodeller, P.D.; Gaitzsch, J.; Braun, G.B.; Willmore, A.M.A.; Ruoslahti, E.; Battaglia, G.; Teesalu, T. Paclitaxel-Loaded Polymersomes for Enhanced Intraperitoneal Chemotherapy. Mol. Cancer Ther. 2016, 15, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, B.; Daly, J.L.; Simón-Gracia, L.; Klein, K.; Weeratunga, S.; Antón-Plágaro, C.; Tobi, A.; Hodgson, L.; Lewis, P.A.; Heesom, K.J.; et al. ESCPE-1 Mediates Retrograde Endosomal Sorting of the SARS-CoV-2 Host Factor Neuropilin-1. Proc. Natl. Acad. Sci. USA 2022, 119, e2201980119. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchetti, L.; Simon-Gracia, L.; Lico, C.; Mancuso, M.; Baschieri, S.; Santi, L.; Teesalu, T. Targeting of Tomato Bushy Stunt Virus with a Genetically Fused C-End Rule Peptide. Nanomaterials 2023, 13, 1428. https://doi.org/10.3390/nano13081428
Marchetti L, Simon-Gracia L, Lico C, Mancuso M, Baschieri S, Santi L, Teesalu T. Targeting of Tomato Bushy Stunt Virus with a Genetically Fused C-End Rule Peptide. Nanomaterials. 2023; 13(8):1428. https://doi.org/10.3390/nano13081428
Chicago/Turabian StyleMarchetti, Luca, Lorena Simon-Gracia, Chiara Lico, Mariateresa Mancuso, Selene Baschieri, Luca Santi, and Tambet Teesalu. 2023. "Targeting of Tomato Bushy Stunt Virus with a Genetically Fused C-End Rule Peptide" Nanomaterials 13, no. 8: 1428. https://doi.org/10.3390/nano13081428