
In Vitro Models of Bacterial Biofilms: Innovative Tools to Improve Understanding and Treatment of Infections


Abstract
:1. Introduction
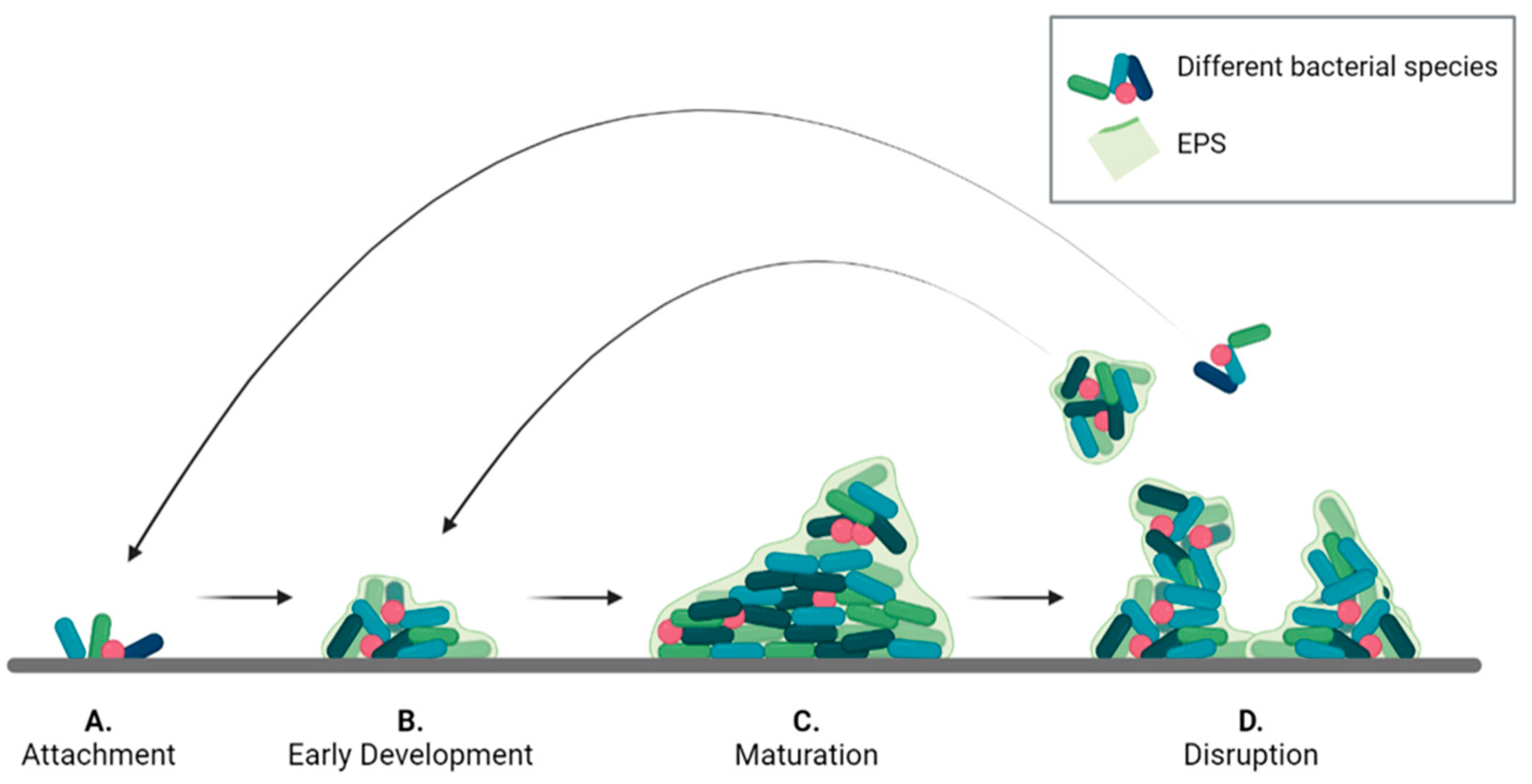
2. Lifecycle and Structure of Biofilms
2.1. The EPS Composition
2.2. Mechanical Properties of Biofilms
2.3. Mechanisms of Antibiotic Resistance in Bacterial Biofilms
3. Biofilm Models
3.1. 3D Models of Bacterial Biofilm
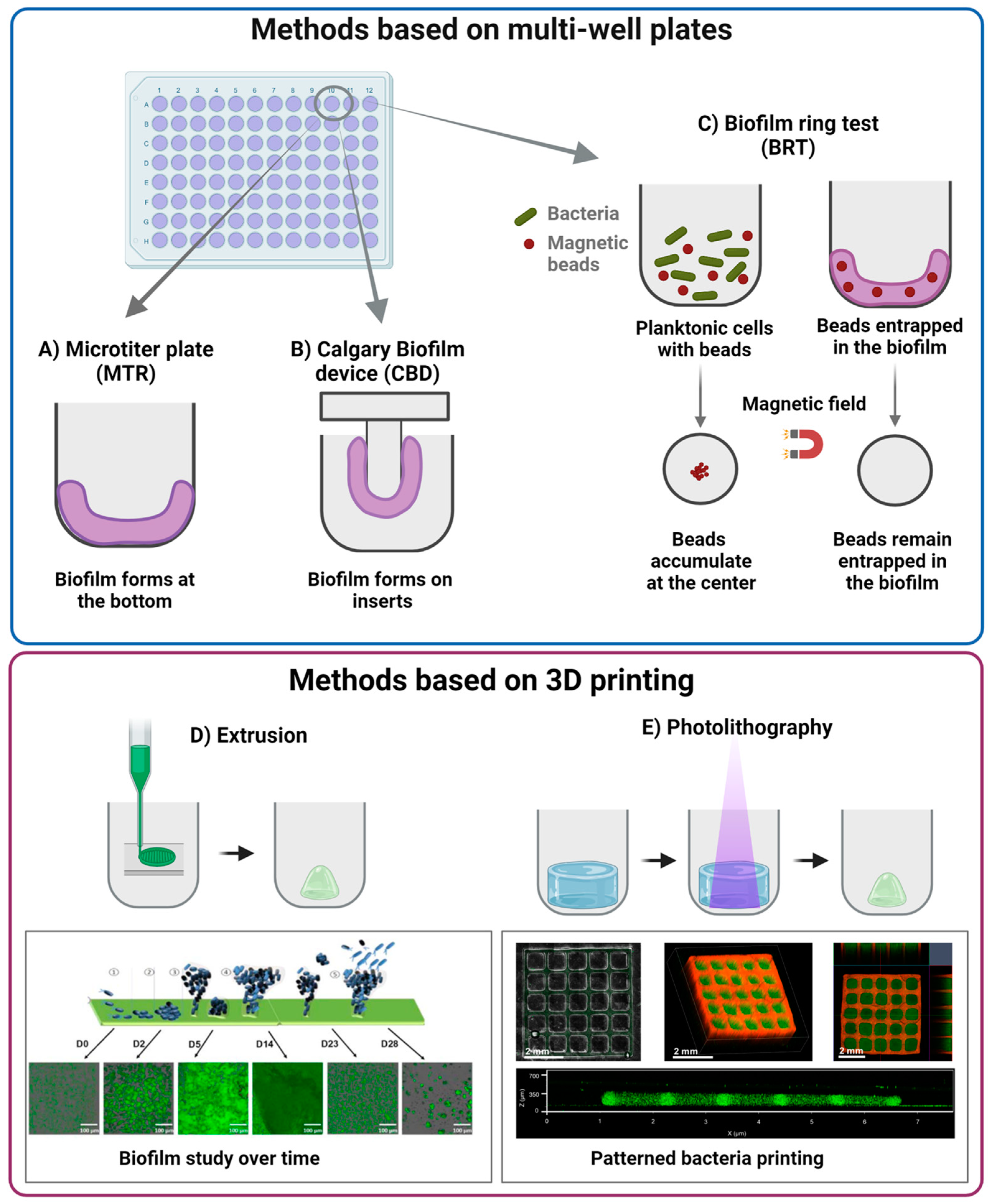
3.2. Static In Vitro Models of Biofilm

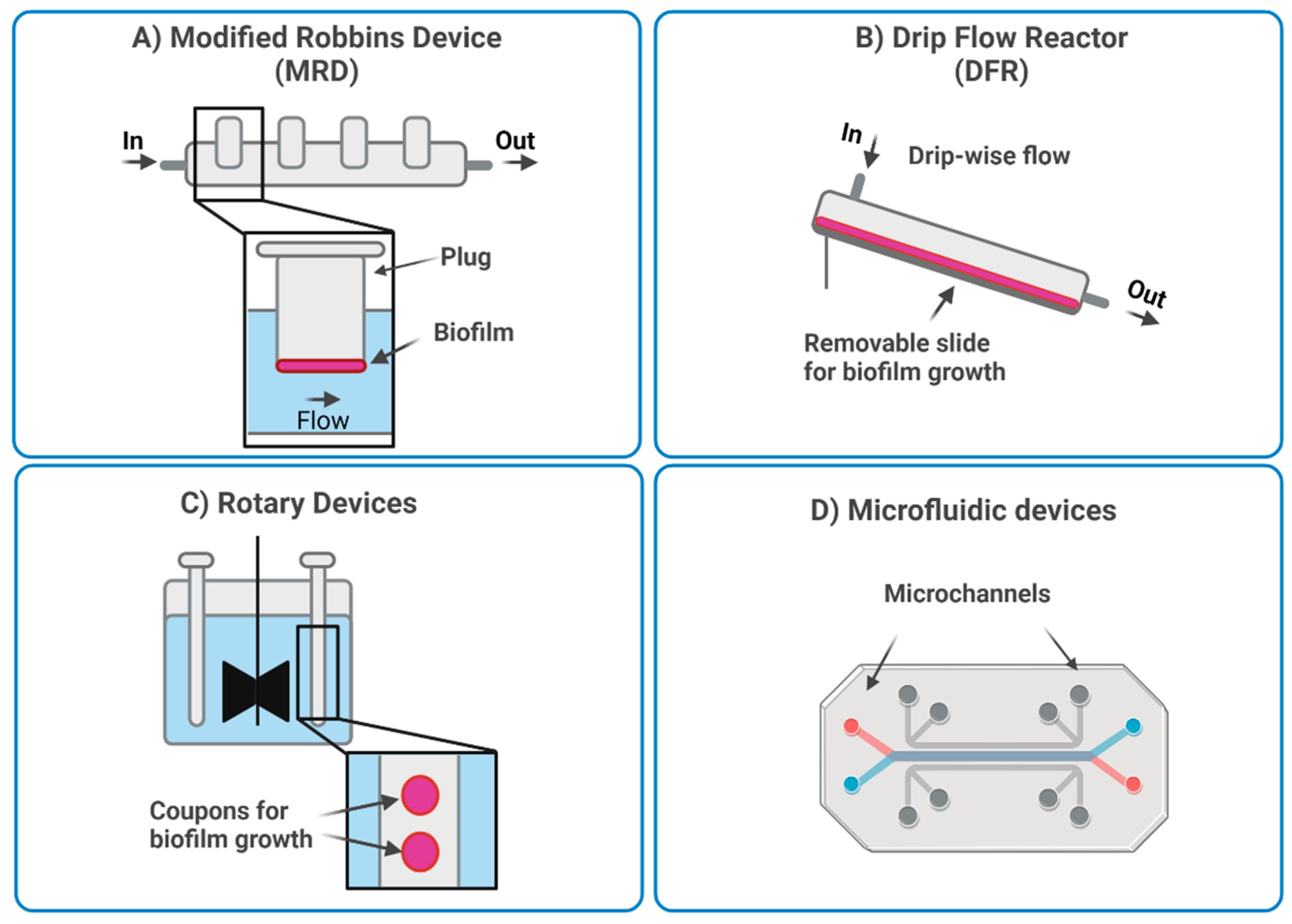
3.3. Dynamic In Vitro Models of Biofilm
3.4. Microcosm Models
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fux, C.A.; Costerton, J.W.; Stewart, P.S.; Stoodley, P. Survival strategies of infectious biofilms. Trends Microbiol. 2005, 13, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Planson, A.-G.; Sauveplane, V.; Dervyn, E.; Jules, M. Bacterial growth physiology and RNA metabolism. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2020, 1863, 194502. [Google Scholar] [CrossRef] [PubMed]
- Hingorani, K.S.; Gierasch, L.M. How bacteria survive an acid trip. Proc. Natl. Acad. Sci. USA 2013, 110, 5279–5280. [Google Scholar] [CrossRef] [Green Version]
- Mathur, J.; Bizzoco, R.W.; Ellis, D.G.; Lipson, D.A.; Poole, A.W.; Levine, R.; Kelley, S.T. Effects of Abiotic Factors on the Phylogenetic Diversity of Bacterial Communities in Acidic Thermal Springs. Appl. Environ. Microbiol. 2007, 73, 2612–2623. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Soto, I.; McTiernan, C.; Gonzalez-Gomez, M.; Ross, A.; Gupta, K.; Suuronen, E.J.; Mah, T.-F.; Griffith, M.; Alarcon, E.I. Mimicking biofilm formation and development: Recent progress in in vitro and in vivo biofilm models. Iscience 2021, 24, 102443. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Dragos, A.; Kovács, A.T. The Peculiar Functions of the Bacterial Extracellular Matrix. Trends Microbiol. 2017, 25, 257–266. [Google Scholar] [CrossRef]
- NIH. NIH Guide: Research on Microbial Biofilms. Available online: https://grants.nih.gov/grants/guide/pa-files/PA-03-047.html (accessed on 14 January 2022).
- Schulze, A.; Mitterer, F.; Pombo, J.P.; Schild, S. Biofilms by bacterial human pathogens: Clinical relevance—development, composition and regulation—therapeutical strategies. Microb. Cell 2021, 8, 28–56. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Moser, C.; Wang, H.-Z.; Høiby, N.; Song, Z.-J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2014, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zimmerli, W.; Sendi, P. Orthopaedic biofilm infections. Apmis 2017, 125, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Pelling, H.; Nzakizwanayo, J.; Milo, S.; Denham, E.; MacFarlane, W.; Bock, L.; Sutton, J.; Jones, B. Bacterial biofilm formation on indwelling urethral catheters. Lett. Appl. Microbiol. 2019, 68, 277–293. [Google Scholar] [CrossRef] [Green Version]
- Nelson, R.E.; Hyun, D.; Jezek, A.; Samore, M.H. Mortality, Length of Stay, and Healthcare Costs Associated with Multidrug-Resistant Bacterial Infections among Elderly Hospitalized Patients in the United States. Clin. Infect. Dis. 2021, 74, 1070–1080. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the Natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Kassinger, S.J.; van Hoek, M.L. Biofilm architecture: An emerging synthetic biology target. Synth. Syst. Biotechnol. 2020, 5, 1–10. [Google Scholar] [CrossRef]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as Complex Differentiated Communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjarnsholt, T.; Whiteley, M.; Rumbaugh, K.P.; Stewart, P.S.; Jensen, P.; Frimodt-Møller, N. The importance of understanding the infectious microenvironment. Lancet Infect. Dis. 2021, 22, e88–e92. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Magana, M.; Sereti, C.; Ioannidis, A.; Mitchell, C.A.; Ball, A.R.; Magiorkinis, E.; Chatzipanagiotou, S.; Hamblin, M.R.; Hadjifrangiskou, M.; Tegos, G.P. Options and Limitations in Clinical Investigation of Bacterial Biofilms. Clin. Microbiol. Rev. 2018, 31, e00084-16. [Google Scholar] [CrossRef] [Green Version]
- Coenye, T.; Nelis, H.J. In vitro and in vivo model systems to study microbial biofilm formation. J. Microbiol. Methods 2010, 83, 89–105. [Google Scholar] [CrossRef]
- Ning, E.; Turnbull, G.; Clarke, J.; Picard, F.; Riches, P.; Vendrell, M.; Graham, D.; Wark, A.W.; Faulds, K.; Shu, W. 3D bioprinting of mature bacterial biofilms for antimicrobial resistance drug testing. Biofabrication 2019, 11, 045018. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.-G.; Choi, J.; Kim, S.-K.; Lee, J.-H.; Kwon, S. Embedded Biofilm, a New Biofilm Model Based on the Embedded Growth of Bacteria. Appl. Environ. Microbiol. 2015, 81, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kaplan, J.B.; Lee, W.Y. Microfluidic devices for studying growth and detachment of Staphylococcus epidermidis biofilms. Biomed. Microdevices 2008, 10, 489–498. [Google Scholar] [CrossRef]
- Chen, X.; Lorenzen, J.; Xu, Y.; Jonikaite, M.; Thaarup, I.C.; Bjarnsholt, T.; Kirketerp-Møller, K.; Thomsen, T.R. A novel chronic wound biofilm model sustaining coexistence of Pseudomonas aeruginosa and Staphylococcus aureus suitable for testing of antibiofilm effect of antimicrobial solutions and wound dressings. Wound Repair Regen. 2021, 29, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Grassi, L.; Batoni, G.; Ostyn, L.; Rigole, P.; Van den Bossche, S.; Rinaldi, A.C.; Maisetta, G.; Esin, S.; Coenye, T.; Crabbe, A. The Antimicrobial Peptide lin-SB056-1 and Its Dendrimeric Derivative Prevent Pseudomonas aeruginosa Biofilm Formation in Physiologically Relevant Models of Chronic Infections. Front. Microbiol. 2019, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa Displays Multiple Phenotypes during Development as a Biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef] [Green Version]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Wu, Y.; Zitelli, J.P.; TenHuisen, K.S.; Yu, X.; Libera, M.R. Differential response of Staphylococci and osteoblasts to varying titanium surface roughness. Biomaterials 2011, 32, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Hospenthal, M.K.; Costa, T.R.D.; Waksman, G. A comprehensive guide to pilus biogenesis in Gram-negative bacteria. Nat. Rev. Microbiol. 2017, 15, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Stones, D.H.; Krachler, A.M. Against the tide: The role of bacterial adhesion in host colonization. Biochem. Soc. Trans. 2016, 44, 1571–1580. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.C.; Blair, K.M.; Salama, N.R. Staying in Shape: The Impact of Cell Shape on Bacterial Survival in Diverse Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, K.; Camper, A.K. Characterization of Phenotypic Changes in Pseudomonas putida in Response to Surface-Associated Growth. J. Bacteriol. 2001, 183, 6579–6589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef] [PubMed]
- Dominiak, D.M.; Nielsen, J.L.; Nielsen, P.H. Extracellular DNA is abundant and important for microcolony strength in mixed microbial biofilms. Environ. Microbiol. 2010, 13, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Xiao, J.; Klein, M.I.; Jeon, J.G. Exopolysaccharides Produced by Streptococcus mutans Glucosyltransferases Modulate the Establishment of Microcolonies within Multispecies Biofilms. J. Bacteriol. 2010, 192, 3024–3032. [Google Scholar] [CrossRef] [Green Version]
- Su, P.-T.; Liao, C.-T.; Roan, J.-R.; Wang, S.-H.; Chiou, A.; Syu, W.-J. Bacterial Colony from Two-Dimensional Division to Three-Dimensional Development. PLoS ONE 2012, 7, e48098. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Hunter, R.C.; Beveridge, T.J. Application of a pH-Sensitive Fluoroprobe (C-SNARF-4) for pH Microenvironment Analysis in Pseudomonas aeruginosa Biofilms. Appl. Environ. Microbiol. 2005, 71, 2501–2510. [Google Scholar] [CrossRef] [Green Version]
- Iii, M.C.W.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of Antibiotic Penetration, Oxygen Limitation, and Low Metabolic Activity to Tolerance of Pseudomonas aeruginosa Biofilms to Ciprofloxacin and Tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Werner, E.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Heydorn, A.; Molin, S.; Pitts, B.; Stewart, P.S. Stratified Growth in Pseudomonas aeruginosa Biofilms. Appl. Environ. Microbiol. 2004, 70, 6066–6075. [Google Scholar] [CrossRef] [Green Version]
- Klausen, M.; Heydorn, A.; Ragas, P.; Lambertsen, L.; Aaes-Jørgensen, A.; Molin, S.; Tolker-Nielsen, T. Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol. Microbiol. 2003, 48, 1511–1524. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-C.; Dean, S.; Marakasova, E.S.; Nwabueze, A.O.; van Hoek, M.L. Chitinases Are Negative Regulators of Francisella novicida Biofilms. PLoS ONE 2014, 9, e93119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, S.; Lebert, I.; Andant, C.; Talon, R. Interaction in dual species biofilms between Staphylococcus xylosus and Staphylococcus aureus. Int. J. Food Microbiol. 2020, 326, 108653. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Barraza, J.P.; Arthur, R.A.; Hara, A.; Lewis, K.; Liu, Y.; Scisci, E.L.; Hajishengallis, E.; Whiteley, M.; Koo, H. Spatial mapping of polymicrobial communities reveals a precise biogeography associated with human dental caries. Proc. Natl. Acad. Sci. USA 2020, 117, 12375–12386. [Google Scholar] [CrossRef]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmølle, M.; Stewart, P.S.; Bjarnsholt, T. The biofilm life cycle: Expanding the conceptual model of biofilm formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. Escaping the biofilm in more than one way: Desorption, detachment or dispersion. Curr. Opin. Microbiol. 2016, 30, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Rumbaugh, K.P.; Sauer, K. Biofilm dispersion. Nat. Rev. Microbiol. 2020, 18, 571–586. [Google Scholar] [CrossRef]
- De Beer, D.; Stoodley, P.; Roe, F.; Lewandowski, Z. Effects of biofilm structures on oxygen distribution and mass transport. Biotechnol. Bioeng. 1994, 43, 1131–1138. [Google Scholar] [CrossRef]
- Lawrence, J.R.; Korber, D.R.; Hoyle, B.D.; Costerton, J.W.; Caldwell, D.E. Optical sectioning of microbial biofilms. J. Bacteriol. 1991, 173, 6558–6567. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Gheorghita Puscaselu, R.; Lobiuc, A.; Dimian, M.; Covasa, M. Alginate: From Food Industry to Biomedical Applications and Management of Metabolic Disorders. Polymers 2020, 12, 2417. [Google Scholar] [CrossRef]
- Ryder, C.; Byrd, M.; Wozniak, D.J. Role of polysaccharides in Pseudomonas aeruginosa biofilm development. Curr. Opin. Microbiol. 2007, 10, 644–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Laue, H.; Schenk, A.; Li, H.; Lambertsen, L.; Neu, T.R.; Molin, S.; Ullrich, M.S. Contribution of alginate and levan production to biofilm formation by Pseudomonas syringae. Microbiology 2006, 152, 2909–2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, Y.; Wang, X.; Hinnebusch, B.J.; Preston, J.F., III; Romeo, T. Depolymerization of β-1,6- N -Acetyl- d -Glucosamine Disrupts the Integrity of Diverse Bacterial Biofilms. J. Bacteriol. 2005, 187, 382–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, J.B.; Ragunath, C.; Velliyagounder, K.; Fine, D.H.; Ramasubbu, N. Enzymatic Detachment of Staphylococcus epidermidis Biofilms. Antimicrob. Agents Chemother. 2004, 48, 2633–2636. [Google Scholar] [CrossRef] [Green Version]
- Waryah, C.B.; Wells, K.; Ulluwishewa, D.; Chen-Tan, N.; Gogoi-Tiwari, J.; Ravensdale, J.; Costantino, P.; Gökçen, A.; Vilcinskas, A.; Wiesner, J.; et al. In Vitro Antimicrobial Efficacy of Tobramycin Against Staphylococcus aureus Biofilms in Combination with or without DNase I and/or Dispersin B: A Preliminary Investigation. Microb. Drug Resist. 2017, 23, 384–390. [Google Scholar] [CrossRef]
- Vats, N.; Lee, S.F. Active detachment of Streptococcus mutans cells adhered to epon–hydroxylapatite surfaces coated with salivary proteins in vitro. Arch. Oral Biol. 2000, 45, 305–314. [Google Scholar] [CrossRef]
- Diggle, S.P.; Stacey, R.E.; Dodd, C.; Camara, M.; Williams, P.; Winzer, K. The galactophilic lectin, LecA, contributes to biofilm development in Pseudomonas aeruginosa. Environ. Microbiol. 2006, 8, 1095–1104. [Google Scholar] [CrossRef]
- Tielker, D.; Hacker, S.; Loris, R.; Strathmann, M.; Wingender, J.; Wilhelm, S.; Rosenau, F.; Jaeger, K.-E. Pseudomonas aeruginosa lectin LecB is located in the outer membrane and is involved in biofilm formation. Microbiology 2005, 151, 1313–1323. [Google Scholar] [CrossRef]
- Lasa, I.; Penadés, J.R. Bap: A family of surface proteins involved in biofilm formation. Res. Microbiol. 2006, 157, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA Required for Bacterial Biofilm Formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef] [PubMed]
- Neu, T.R.; Poralla, K. An amphiphilic polysaccharide from an adhesive Rhodococcus strain. FEMS Microbiol. Lett. 1988, 49, 389–392. [Google Scholar] [CrossRef]
- Kundukad, B.; Seviour, T.; Liang, Y.; Rice, S.A.; Kjelleberg, S.; Doyle, P.S. Mechanical properties of the superficial biofilm layer determine the architecture of biofilms. Soft Matter 2016, 12, 5718–5726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böl, M.; Ehret, A.; Albero, A.B.; Hellriegel, J.; Krull, R. Recent advances in mechanical characterisation of biofilm and their significance for material modelling. Crit. Rev. Biotechnol. 2012, 33, 145–171. [Google Scholar] [CrossRef]
- Körstgens, V.; Flemming, H.-C.; Wingender, J.; Borchard, W. Uniaxial compression measurement device for investigation of the mechanical stability of biofilms. J. Microbiol. Methods 2001, 46, 9–17. [Google Scholar] [CrossRef]
- Koza, A.; Hallett, P.D.; Moon, C.D.; Spiers, A.J. Characterization of a novel air–liquid interface biofilm of Pseudomonas fluorescens SBW25. Microbiology 2009, 155, 1397–1406. [Google Scholar] [CrossRef] [Green Version]
- Klapper, I.; Rupp, C.J.; Cargo, R.; Purvedorj, B.; Stoodley, P. Viscoelastic fluid description of bacterial biofilm material properties. Biotechnol. Bioeng. 2002, 80, 289–296. [Google Scholar] [CrossRef]
- Stoodley, P.; Lewandowski, Z.; Boyle, J.D.; Lappin-Scott, H.M. Structural Deformation of Bacterial Biofilms Caused by Short-Term Fluctuations in Fluid Shear: An in situ Investigation of Biofilm Rheology. Biotechnol. Bioeng. 1999, 65, 83–92. [Google Scholar] [CrossRef]
- Galy, O.; Latour-Lambert, P.; Zrelli, K.; Ghigo, J.-M.; Beloin, C.; Henry, N. Mapping of Bacterial Biofilm Local Mechanics by Magnetic Microparticle Actuation. Biophys. J. 2012, 103, 1400–1408. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.; Melo, L.; Oliveira, A.; Salgueiro, R. Polysaccharide Production and Biofilm Formation by Pseudomonas fluorescens: Effects of PH and Surface Material. Colloids Surf. B Biointerfaces 1994, 2, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Tallawi, M.; Opitz, M.; Lieleg, O. Modulation of the mechanical properties of bacterial biofilms in response to environmental challenges. Biomater. Sci. 2017, 5, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-R.; Li, C.-M.; Yu, C.-H.; Lin, Y.-J.; Wu, C.-M.; Harn, I.-C.; Tang, M.-J.; Chen, Y.-T.; Shen, F.-C.; Lu, C.-Y.; et al. The enhancement of biofilm formation in Group B streptococcal isolates at vaginal pH. Med. Microbiol. Immunol. 2012, 202, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.L.; Sutton, M.P.; McKittrick, L.; Stewart, P.S. Chemical and antimicrobial treatments change the viscoelastic properties of bacterial biofilms. Biofouling 2011, 27, 207–215. [Google Scholar] [CrossRef]
- Körstgens, V.; Flemming, H.-C.; Wingender, J.; Borchard, W. Influence of calcium ions on the mechanical properties of a model biofilm of mucoid Pseudomonas aeruginosa. Water Sci. Technol. 2001, 43, 49–57. [Google Scholar] [CrossRef]
- Boudarel, H.; Mathias, J.-D.; Blaysat, B.; Grédiac, M. Towards standardized mechanical characterization of microbial biofilms: Analysis and critical review. npj Biofilms Microbiomes 2018, 4, 1–15. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Harrison, J.J.; Turner, R.J.; Marques, L.L.R.; Ceri, H. Biofilms: A New Understanding of These Microbial Communities Is Driving a Revolution That May Transform the Science of Microbiology. Am. Sci. 2005, 93, 508–515. [Google Scholar] [CrossRef]
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Thaarup, I.C.; Bjarnsholt, T. Current In Vitro Biofilm-Infected Chronic Wound Models for Developing New Treatment Possibilities. Adv. Wound Care 2021, 10, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Alhede, M.; Alhede, M.; Eickhardt-Sørensen, S.R.; Moser, C.; Kühl, M.; Jensen, P.; Høiby, N. The in vivo biofilm. Trends Microbiol. 2013, 21, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Coenye, T. In Vitro and In Vivo Biofilm Wound Models and Their Application. In Advances in Microbiology, Infectious Diseases and Public Health; Donelli, G., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2015; Volume 897, pp. 15–32. [Google Scholar] [CrossRef]
- Lebeaux, D.; Chauhan, A.; Rendueles, O.; Beloin, C. From in vitro to in vivo Models of Bacterial Biofilm-Related Infections. Pathogens 2013, 2, 288–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelo-Branco, D.D.S.C.M.; Amando, B.R.; Ocadaque, C.J.; de Aguiar, L.; Paiva, D.D.D.Q.; Diógenes, E.M.; Guedes, G.M.D.M.; Costa, C.L.; Santos-Filho, A.S.P.; de Andrade, A.R.C.; et al. Mini-review: From in vitro to ex vivo studies: An overview of alternative methods for the study of medical biofilms. Biofouling 2020, 36, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Mi, G.; Wang, M.; Webster, T.J. In vitro and ex vivo systems at the forefront of infection modeling and drug discovery. Biomaterials 2018, 198, 228–249. [Google Scholar] [CrossRef] [PubMed]
- Raic, A.; Riedel, S.; Kemmling, E.; Bieback, K.; Overhage, J.; Lee-Thedieck, C. Biomimetic 3D in vitro model of biofilm triggered osteomyelitis for investigating hematopoiesis during bone marrow infections. Acta Biomater. 2018, 73, 250–262. [Google Scholar] [CrossRef]
- Spittaels, K.-J.; Coenye, T. Developing an in vitro artificial sebum model to study Propionibacterium acnes biofilms. Anaerobe 2017, 49, 21–29. [Google Scholar] [CrossRef]
- Marsh, P.D.; Zaura, E. Dental biofilm: Ecological interactions in health and disease. J. Clin. Periodontol. 2017, 44, S12–S22. [Google Scholar] [CrossRef]
- Smith, W.P.J.; Davit, Y.; Osborne, J.M.; Kim, W.; Foster, K.R.; Pitt-Francis, J.M. Cell morphology drives spatial patterning in microbial communities. Proc. Natl. Acad. Sci. USA 2016, 114, E280–E286. [Google Scholar] [CrossRef] [Green Version]
- Pavlovsky, L.; Younger, J.G.; Solomon, M.J. In siturheology of Staphylococcus epidermidis bacterial biofilms. Soft Matter 2013, 9, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Pavlovsky, L.; Sturtevant, R.A.; Younger, J.G.; Solomon, M.J. Effects of Temperature on the Morphological, Polymeric, and Mechanical Properties of Staphylococcus epidermidis Bacterial Biofilms. Langmuir 2015, 31, 2036–2042. [Google Scholar] [CrossRef] [Green Version]
- Manetti, A.G.O.; Köller, T.; Becherelli, M.; Buccato, S.; Kreikemeyer, B.; Podbielski, A.; Grandi, G.; Margarit, I. Environmental Acidification Drives S. pyogenes Pilus Expression and Microcolony Formation on Epithelial Cells in a FCT-Dependent Manner. PLoS ONE 2010, 5, e13864. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.R. Influence of pH on bacterial gene expression. Mol. Microbiol. 1993, 8, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Dunsmore, B.C.; Jacobsen, A.; Hall-Stoodley, L.; Bass, C.J.; Lappin-Scott, H.M.; Stoodley, P. The influence of fluid shear on the structure and material properties of sulphate-reducing bacterial biofilms. J. Ind. Microbiol. Biotechnol. 2002, 29, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, P.; Hall-Stoodley, L.; Lappin-Scott, H.M. [21] Detachment, surface migration, and other dynamic behavior in bacterial biofilms revealed by digital time-lapse imaging. In Methods in Enzymology; Doyle, R.J., Ed.; Microbial Growth in Biofilms—Part B: Special Environments and Physicochemical Aspects; Academic Press: Cambridge, MA, USA, 2001; Volume 337, pp. 306–319. [Google Scholar] [CrossRef]
- DePas, W.H.; Hufnagel, D.A.; Lee, J.S.; Blanco, L.P.; Bernstein, H.C.; Fisher, S.T.; James, G.A.; Stewart, P.S.; Chapman, M.R. Iron induces bimodal population development by Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 2629–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attinger, C.; Wolcott, R. Clinically Addressing Biofilm in Chronic Wounds. Adv. Wound Care 2012, 1, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Wolcott, R.; Rhoads, D.; Bennett, M.; Wolcott, B.; Gogokhia, L.; Costerton, J.; Dowd, S. Chronic wounds and the medical biofilm paradigm. J. Wound Care 2010, 19, 45–53. [Google Scholar] [CrossRef]
- Wilson, C.; Lukowicz, R.; Merchant, S.; Valquier-Flynn, H.; Caballero, J.; Sandoval, J.; Okuom, M.; Huber, C.; Brooks, T.D.; Wilson, E.; et al. Quantitative and Qualitative Assessment Methods for Biofilm Growth: A Mini-Review. Res. Rev. J. Eng. Technol. 2017, 6. Available online: http://www.rroij.com/open-access/quantitative-and-qualitative-assessment-methods-for-biofilm-growth-a-minireview-.pdf (accessed on 19 February 2023).
- Sweeney, E.; Lovering, A.; Bowker, K.; MacGowan, A.; Nelson, S. An in vitro biofilm model of Staphylococcus aureus infection of bone. Lett. Appl. Microbiol. 2019, 68, 294–302. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.C.; Haney, E.F.; Akhoundsadegh, N.; Pletzer, D.; Trimble, M.J.; Adriaans, A.E.; Nibbering, P.H.; Hancock, R.E.W. Human organoid biofilm model for assessing antibiofilm activity of novel agents. npj Biofilms Microbiomes 2021, 7, 1–13. [Google Scholar] [CrossRef]
- Merritt, J.H.; Kadouri, D.E.; O'Toole, G.A. Growing and Analyzing Static Biofilms. Curr. Protoc. Microbiol. 2011, 22, 1B.1.1–1B.1.18. [Google Scholar] [CrossRef] [Green Version]
- Bahamondez-Canas, T.F.; Heersema, L.A.; Smyth, H.D.C. Current Status of In Vitro Models and Assays for Susceptibility Testing for Wound Biofilm Infections. Biomedicines 2019, 7, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tambone, E.; Bonomi, E.; Ghensi, P.; Maniglio, D.; Ceresa, C.; Agostinacchio, F.; Caciagli, P.; Nollo, G.; Piccoli, F.; Caola, I.; et al. Rhamnolipid coating reduces microbial biofilm formation on titanium implants: An in vitro study. BMC Oral Health 2021, 21, 1–13. [Google Scholar] [CrossRef]
- Tambone, E.; Marchetti, A.; Ceresa, C.; Piccoli, F.; Anesi, A.; Nollo, G.; Caola, I.; Bosetti, M.; Fracchia, L.; Ghensi, P.; et al. Counter-Acting Candida albicans-Staphylococcus aureus Mixed Biofilm on Titanium Implants Using Microbial Biosurfactants. Polymers 2021, 13, 2420. [Google Scholar] [CrossRef]
- Ceresa, C.; Rinaldi, M.; Tessarolo, F.; Maniglio, D.; Fedeli, E.; Tambone, E.; Caciagli, P.; Banat, I.M.; De Rienzo, M.A.D.; Fracchia, L. Inhibitory Effects of Lipopeptides and Glycolipids on C. albicans–Staphylococcus spp. Dual-Species Biofilms. Front. Microbiol. 2021, 11, 545654. [Google Scholar] [CrossRef] [PubMed]
- Ceresa, C.; Rinaldi, M.; Chiono, V.; Carmagnola, I.; Allegrone, G.; Fracchia, L. Lipopeptides from Bacillus subtilis AC7 inhibit adhesion and biofilm formation of Candida albicans on silicone. Antonie Van Leeuwenhoek 2016, 109, 1375–1388. [Google Scholar] [CrossRef]
- Ceresa, C.; Tessarolo, F.; Maniglio, D.; Tambone, E.; Carmagnola, I.; Fedeli, E.; Caola, I.; Nollo, G.; Chiono, V.; Allegrone, G.; et al. Medical-Grade Silicone Coated with Rhamnolipid R89 Is Effective against Staphylococcus spp. Biofilms. Molecules 2019, 24, 3843. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.-X.; Tu, B.; Shen, Y.; Du, L.; Wang, L.; Guo, S.-R.; Tang, T.-T. Quaternized Chitosan Inhibits icaA Transcription and Biofilm Formation by Staphylococcus on a Titanium Surface. Antimicrob. Agents Chemother. 2011, 55, 860–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceresa, C.; Fracchia, L.; Williams, M.; Banat, I.; De Rienzo, M.D. The effect of sophorolipids against microbial biofilms on medical-grade silicone. J. Biotechnol. 2019, 309, 34–43. [Google Scholar] [CrossRef]
- Allegrone, G.; Ceresa, C.; Rinaldi, M.; Fracchia, L. Diverse Effects of Natural and Synthetic Surfactants on the Inhibition of Staphylococcus aureus Biofilm. Pharmaceutics 2021, 13, 1172. [Google Scholar] [CrossRef]
- Gupta, S.; Thakur, J.; Pal, S.; Gupta, R.; Mishra, D.; Kumar, S.; Yadav, K.; Saini, A.; Yavvari, P.S.; Vedantham, M.; et al. Cholic Acid-Peptide Conjugates as Potent Antimicrobials against Interkingdom Polymicrobial Biofilms. Antimicrob. Agents Chemother. 2019, 63, e00520-19. [Google Scholar] [CrossRef]
- Adam, B.; Baillie, G.S.; Douglas, L.J. Mixed species biofilms of Candida albicans and Staphylococcus epidermidis. J. Med. Microbiol. 2002, 51, 344–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Chen, H.; Sun, Y.; Zhang, X.; Lu, H.; Li, J.; Cao, J.; Zhou, T. Characterization of the mechanism and impact of staphylokinase on the formation of Candida albicans and Staphylococcus aureus polymicrobial biofilms. J. Med. Microbiol. 2019, 68, 355–367. [Google Scholar] [CrossRef]
- Budzyńska, A.; Różalska, S.; Sadowska, B.; Różalska, B. Candida albicans/Staphylococcus aureus Dual-Species Biofilm as a Target for the Combination of Essential Oils and Fluconazole or Mupirocin. Mycopathologia 2017, 182, 989–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werthén, M.; Henriksson, L.; Jensen, P.; Sternberg, C.; Givskov, M.; Bjarnsholt, T. An in vitro model of bacterial infections in wounds and other soft tissues. Apmis 2010, 118, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Garcia-Fernandez, M.J.; Lenoir, J.; De Meyer, L.; Remon, J.-P.; De Beer, T.; Concheiro, A.; Alvarez-Lorenzo, C.; Coenye, T. Dressings Loaded with Cyclodextrin-Hamamelitannin Complexes Increase Staphylococcus aureus Susceptibility Toward Antibiotics Both in Single as well as in Mixed Biofilm Communities. Macromol. Biosci. 2016, 16, 859–869. [Google Scholar] [CrossRef]
- Percival, S.L.; Bowler, P.G.; Dolman, J. Antimicrobial activity of silver-containing dressings on wound microorganisms using an in vitro biofilm model. Int. Wound J. 2007, 4, 186–191. [Google Scholar] [CrossRef]
- Townsend, E.M.; Sherry, L.; Rajendran, R.; Hansom, D.; Butcher, J.; Mackay, W.G.; Williams, C.; Ramage, G. Development and characterisation of a novel three-dimensional inter-kingdom wound biofilm model. Biofouling 2016, 32, 1259–1270. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.L.; Ritschdorff, E.T.; Whiteley, M.; Shear, J.B. 3D printing of microscopic bacterial communities. Proc. Natl. Acad. Sci. USA 2013, 110, 18380–18385. [Google Scholar] [CrossRef] [Green Version]
- Schaffner, M.; Rühs, P.A.; Coulter, F.; Kilcher, S.; Studart, A.R. 3D printing of bacteria into functional complex materials. Sci. Adv. 2017, 3, eaao6804. [Google Scholar] [CrossRef] [Green Version]
- Schmieden, D.T.; Vázquez, S.J.B.; Sangüesa, H.; van der Does, M.; Idema, T.; Meyer, A.S. Printing of Patterned, Engineered E. coli Biofilms with a Low-Cost 3D Printer. ACS Synth. Biol. 2018, 7, 1328–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubbin, K.; Dong, Z.; Park, D.M.; Alvarado, J.; Su, J.; Wasson, E.; Robertson, C.; Jackson, J.; Bose, A.; Moya, M.L.; et al. Projection Microstereolithographic Microbial Bioprinting for Engineered Biofilms. Nano Lett. 2021, 21, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Aubin-Tam, M.-E.; Meyer, A.S. 3D Printing for the Fabrication of Biofilm-Based Functional Living Materials. ACS Synth. Biol. 2019, 8, 1564–1567. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yuk, H.; Lin, S.; Parada, G.A.; Tang, T.-C.; Tham, E.; de la Fuente-Nunez, C.; Lu, T.K.; Zhao, X. 3D Printing of Living Responsive Materials and Devices. Adv. Mater. 2017, 30, 1704821. [Google Scholar] [CrossRef] [PubMed]
- Raad, I.; Hanna, H.; Dvorak, T.; Chaiban, G.; Hachem, R. Optimal Antimicrobial Catheter Lock Solution, Using Different Combinations of Minocycline, EDTA, and 25-Percent Ethanol, Rapidly Eradicates Organisms Embedded in Biofilm. Antimicrob. Agents Chemother. 2007, 51, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Blanc, V.; Isabal, S.; Sánchez, M.C.; Llama-Palacios, A.; Herrera, D.; Sanz, M.; León, R. Characterization and application of a flow system for in vitro multispecies oral biofilm formation. J. Periodontal Res. 2013, 49, 323–332. [Google Scholar] [CrossRef]
- Goeres, D.M.; Hamilton, M.A.; Beck, N.A.; Buckingham-Meyer, K.; Hilyard, J.D.; Loetterle, L.R.; Lorenz, L.A.; Walker, D.K.; Stewart, P.S. A method for growing a biofilm under low shear at the air–liquid interface using the drip flow biofilm reactor. Nat. Protoc. 2009, 4, 783–788. [Google Scholar] [CrossRef]
- Xu, K.D.; Stewart, P.S.; Xia, F.; Huang, C.-T.; McFeters, G.A. Spatial Physiological Heterogeneity in Pseudomonas aeruginosa Biofilm Is Determined by Oxygen Availability. Appl. Environ. Microbiol. 1998, 64, 4035–4039. [Google Scholar] [CrossRef] [Green Version]
- Agostinho, A.; James, G.; Wazni, O.; Citron, M.; Wilkoff, B.D. Inhibition of Staphylococcus aureus Biofilms by a Novel Antibacterial Envelope for Use with Implantable Cardiac Devices. Clin. Transl. Sci. 2009, 2, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Plazonic, F.; LuTheryn, G.; Hind, C.; Clifford, M.; Gray, M.; Stride, E.; Glynne-Jones, P.; Hill, M.; Sutton, J.M.; Carugo, D. Bactericidal Effect of Ultrasound-Responsive Microbubbles and Sub-inhibitory Gentamicin against Pseudomonas aeruginosa Biofilms on Substrates with Differing Acoustic Impedance. Ultrasound Med. Biol. 2022, 48, 1888–1898. [Google Scholar] [CrossRef]
- Duckworth, P.F.; Rowlands, R.S.; Barbour, M.E.; Maddocks, S.E. A novel flow-system to establish experimental biofilms for modelling chronic wound infection and testing the efficacy of wound dressings. Microbiol. Res. 2018, 215, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.; Neethirajan, S.; Weng, X. Microfluidic wound model for studying the behaviors of Pseudomonas aeruginosa in polymicrobial biofilms. Biotechnol. Bioeng. 2015, 112, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Ren, B.; He, J.; Peng, X.; Guo, Q.; Zheng, L.; Li, J.; Dai, H.; Chen, V.; Zhang, L.; et al. Candida albicans promotes tooth decay by inducing oral microbial dysbiosis. ISME J. 2020, 15, 894–908. [Google Scholar] [CrossRef]
- Sánchez, M.; Llama-Palacios, A.; Fernández, E.; Figuero, E.; Marín, M.; León, R.; Blanc, V.; Herrera, D.; Sanz, M. An in vitro biofilm model associated to dental implants: Structural and quantitative analysis of in vitro biofilm formation on different dental implant surfaces. Dent. Mater. 2014, 30, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Carterson, A.J.; zu Bentrup, K.H.; Ott, C.M.; Clarke, M.S.; Pierson, D.L.; Vanderburg, C.R.; Buchanan, K.L.; Nickerson, C.A.; Schurr, M.J. A549 Lung Epithelial Cells Grown as Three-Dimensional Aggregates: Alternative Tissue Culture Model for Pseudomonas aeruginosa Pathogenesis. Infect. Immun. 2005, 73, 1129–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crabbé, A.; Liu, Y.; Matthijs, N.; Rigole, P.; De La Fuente-Nùñez, C.; Davis, R.; Ledesma, M.A.; Sarker, S.; Van Houdt, R.; Hancock, R.E.W.; et al. Antimicrobial efficacy against Pseudomonas aeruginosa biofilm formation in a three-dimensional lung epithelial model and the influence of fetal bovine serum. Sci. Rep. 2017, 7, srep43321. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Naha, P.C.; Hwang, G.; Kim, D.; Huang, Y.; Simon-Soro, A.; Jung, H.-I.; Ren, Z.; Li, Y.; Gubara, S.; et al. Topical ferumoxytol nanoparticles disrupt biofilms and prevent tooth decay in vivo via intrinsic catalytic activity. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhou, J.; Yuan, L.; Wu, F.; Xie, L.; Yan, X.; Li, H.; Li, Y.; Shi, L.; Hu, R.; et al. Neighboring Carboxylic Acid Boosts Peroxidase-Like Property of Metal-Phenolic Nano-Networks in Eradicating Streptococcus mutans Biofilms. Small 2022, 19, 2206657. [Google Scholar] [CrossRef]
- Besser, M.; Dietrich, M.; Weber, L.; Rembe, J.D.; Stuermer, E.K. Efficacy of antiseptics in a novel 3-dimensional human plasma biofilm model (hpBIOM). Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Zhou, X.; Huang, X.; Zhu, C.; Weir, M.D.; Melo, M.A.; Bonavente, A.; Lynch, C.D.; Imazato, S.; Oates, T.W.; et al. Antibacterial and remineralizing nanocomposite inhibit root caries biofilms and protect root dentin hardness at the margins. J. Dent. 2020, 97, 103344. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Parameter | Effects on Bacterial Metabolism | Effects on EPS Composition | Effects on Biofilm Mechanical Properties | REF |
|---|---|---|---|---|
| Bacterial species | Different strains have different metabolic activity | Bacteria-specific EPS Quantity of secreted EPS | Biofilms produced by different bacterial species have different mechanical properties | [16,66,73] |
| Multispecies | Bacteria cooperate in production and metabolization of nutrients | Secretion of different EPS components | Cell shape influences bacteria disposition and, thus, biofilm architecture | [16,89,90] |
| Temperature | Affects cell viability and metabolism | Quantity and composition of EPS | Affects biofilm stiffness | [71,91,92] |
| pH | Alters gene expression | Quantity and composition of secreted EPS | Bacteria grown under optimal pH conditions produce stiffer biofilms | [72,74,93,94] |
| Shear flow | Stimulates bacteria detachment | - | Higher shear flow results in higher stiffness | [69,70,95,96] |
| Nutrients and oxygen | Development of different bacteria phenotypes | Bacteria-specific EPS | Biofilm produced by different bacterial species have different mechanical properties. | [5,51] |
| Ion concentration | Alters gene expression | Quantity and composition of secreted EPS | Multivalent cations enhance stiffness | [75,76,97] |
| Tissue | Application | Main Bacterial Species Involved [9] | Phase of Biofilm Development |
|---|---|---|---|
| Skin | Wound healing | S. aureus S. epidermidis P. aeruginosa | Adhesion Dispersion |
| Acne | Propionibacterium acnes Propionibacterium cellulitis Propionibacterium erysipelas. | Adhesion Dispersion | |
| Bone | Osteomyelitis | S. aureus S. epidermidis | Dispersion |
| Dental plaques | Streptococcus mutans Lactobacillus spp. Actinomyces spp. | Adhesion | |
| Lung | Cystic fibrosis | P. aeruginosa | Adhesion Dispersion |
| Implanted devices and catheters | Implant infection | S. aureus S. epidermidis Enterococcus faecalis E. coli Klebsiella pneumoniae Proteus mirabilis P. aeruginosa | Adhesion |
| Bacteria | Bioink | 3D-Printing Method | Pros | Cons | REF |
|---|---|---|---|---|---|
| P. putida, A. xylinum | Hyaluronic acid k-carrageenan fumed silica | Extrusion | Multispecies biofilm. Bacteria produced cellulose to replace the bioink. | No characterization on the biofilm stage. | [123] |
| E. coli | Alginate | Extrusion | Stability >1 week. Bacteria produced their own EPS to replace alginate. | Use of genetically modified bacteria that secrete EPS following external stimulation. | [124] |
| E. coli, P. aeruginosa | Alginate | Extrusion | Stability >4 weeks. Suitable for aerobic and anaerobic bacteria. | Minimum thickness of 0.25 mm (natural biofilm can be <0.1 mm). | [22] |
| S. aureus, P. aeruginosa | Gelatin—A | Photolithography | Multispecies biofilm. Study of cell-cell communication. | Expensive. Low-throughput analyses. | [122] |
| E. coli | PEG diacrylate (PEGDa) | Photolithography | High resolution (10 µm). Complex geometries. Modulation of the mechanical properties of the bioink. | Low stability (3 days). | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crivello, G.; Fracchia, L.; Ciardelli, G.; Boffito, M.; Mattu, C. In Vitro Models of Bacterial Biofilms: Innovative Tools to Improve Understanding and Treatment of Infections. Nanomaterials 2023, 13, 904. https://doi.org/10.3390/nano13050904
Crivello G, Fracchia L, Ciardelli G, Boffito M, Mattu C. In Vitro Models of Bacterial Biofilms: Innovative Tools to Improve Understanding and Treatment of Infections. Nanomaterials. 2023; 13(5):904. https://doi.org/10.3390/nano13050904
Chicago/Turabian StyleCrivello, G., L. Fracchia, G. Ciardelli, M. Boffito, and C. Mattu. 2023. "In Vitro Models of Bacterial Biofilms: Innovative Tools to Improve Understanding and Treatment of Infections" Nanomaterials 13, no. 5: 904. https://doi.org/10.3390/nano13050904