
Recent Trends in Nano-Fertilizers for Sustainable Agriculture under Climate Change for Global Food Security

 ,
,  , and
, and
Abstract
:1. Introduction
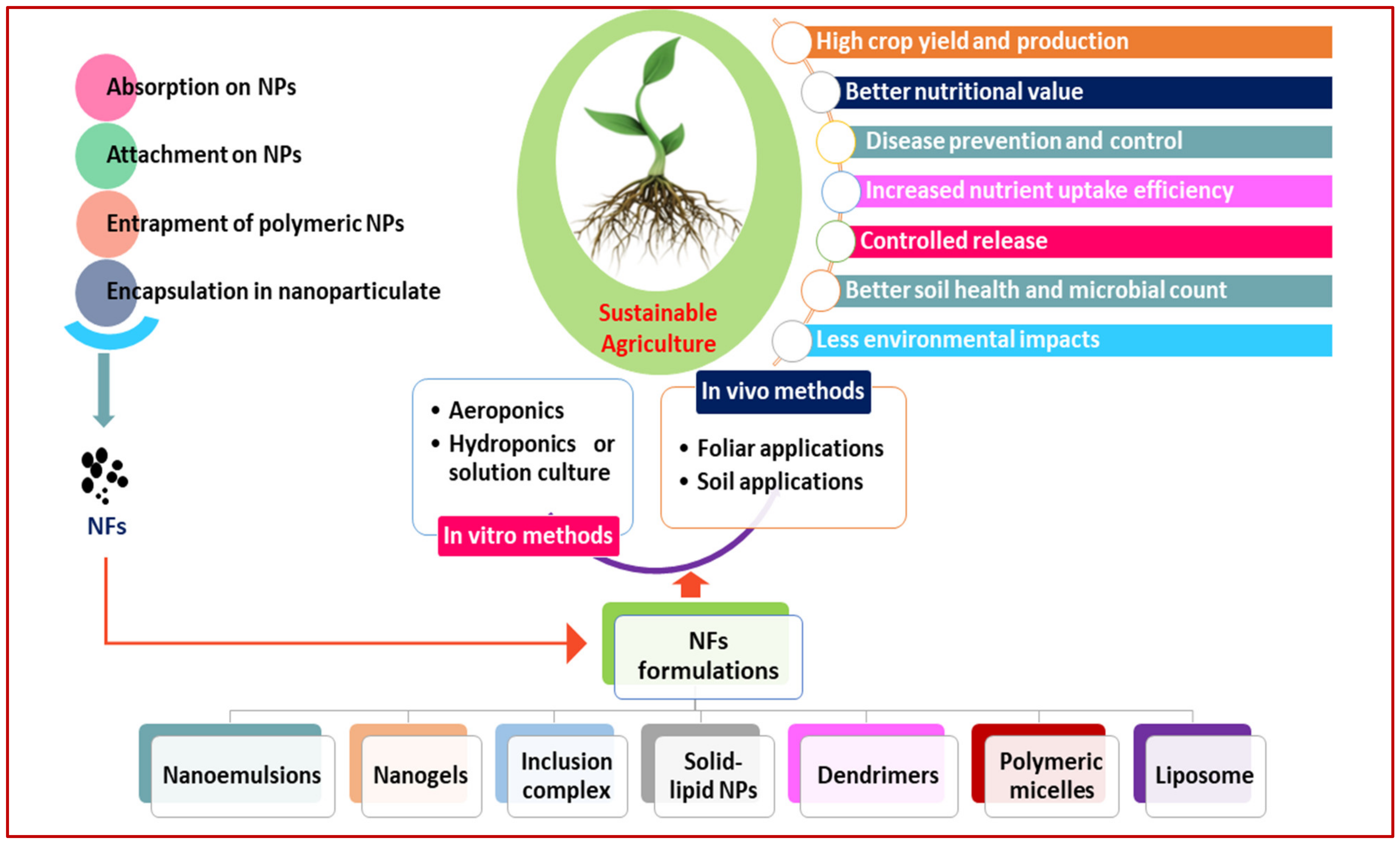
2. Agriculture and Nano-Fertilizers
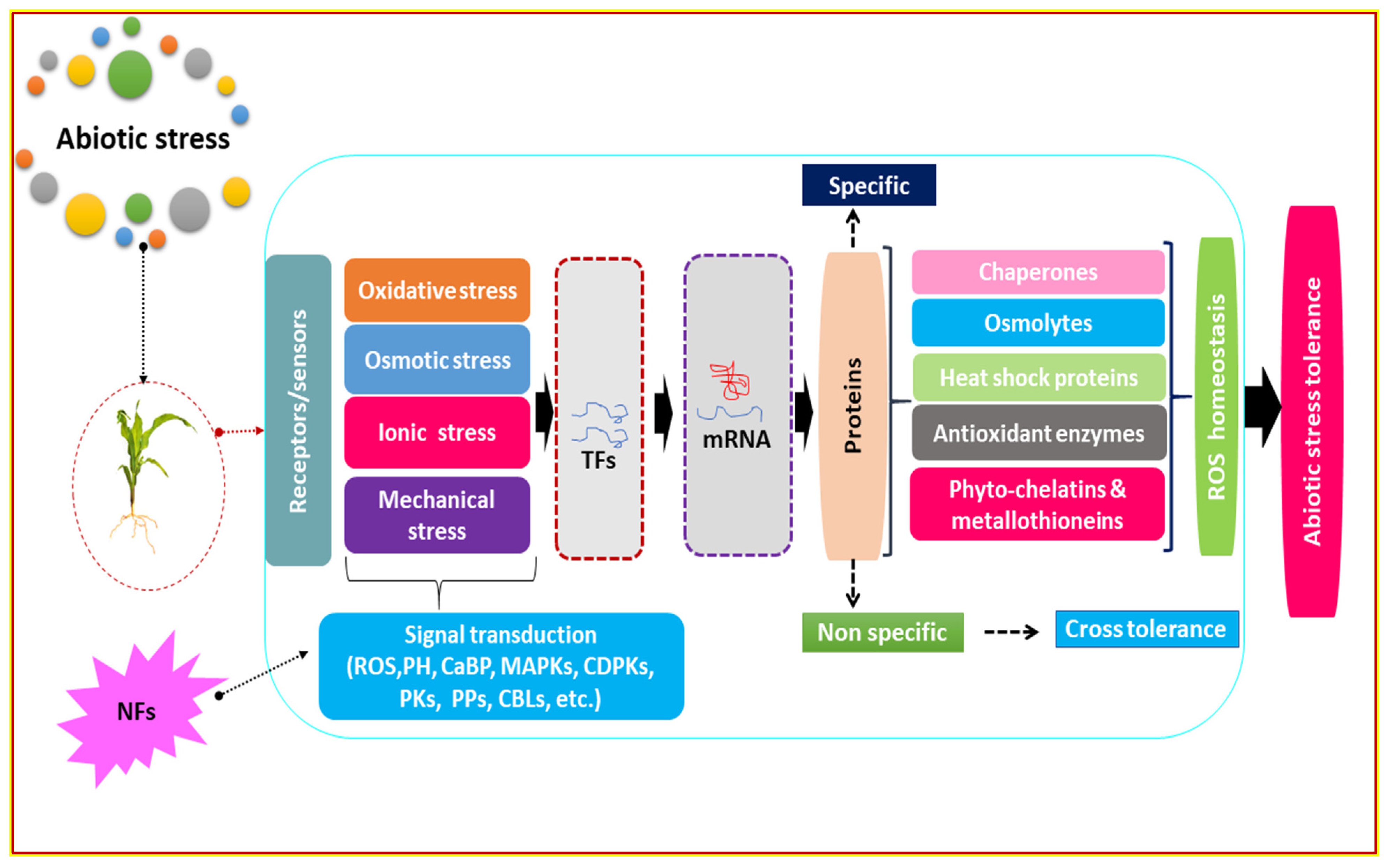
3. Nano-Fertilizers Mitigate Abiotic Stresses
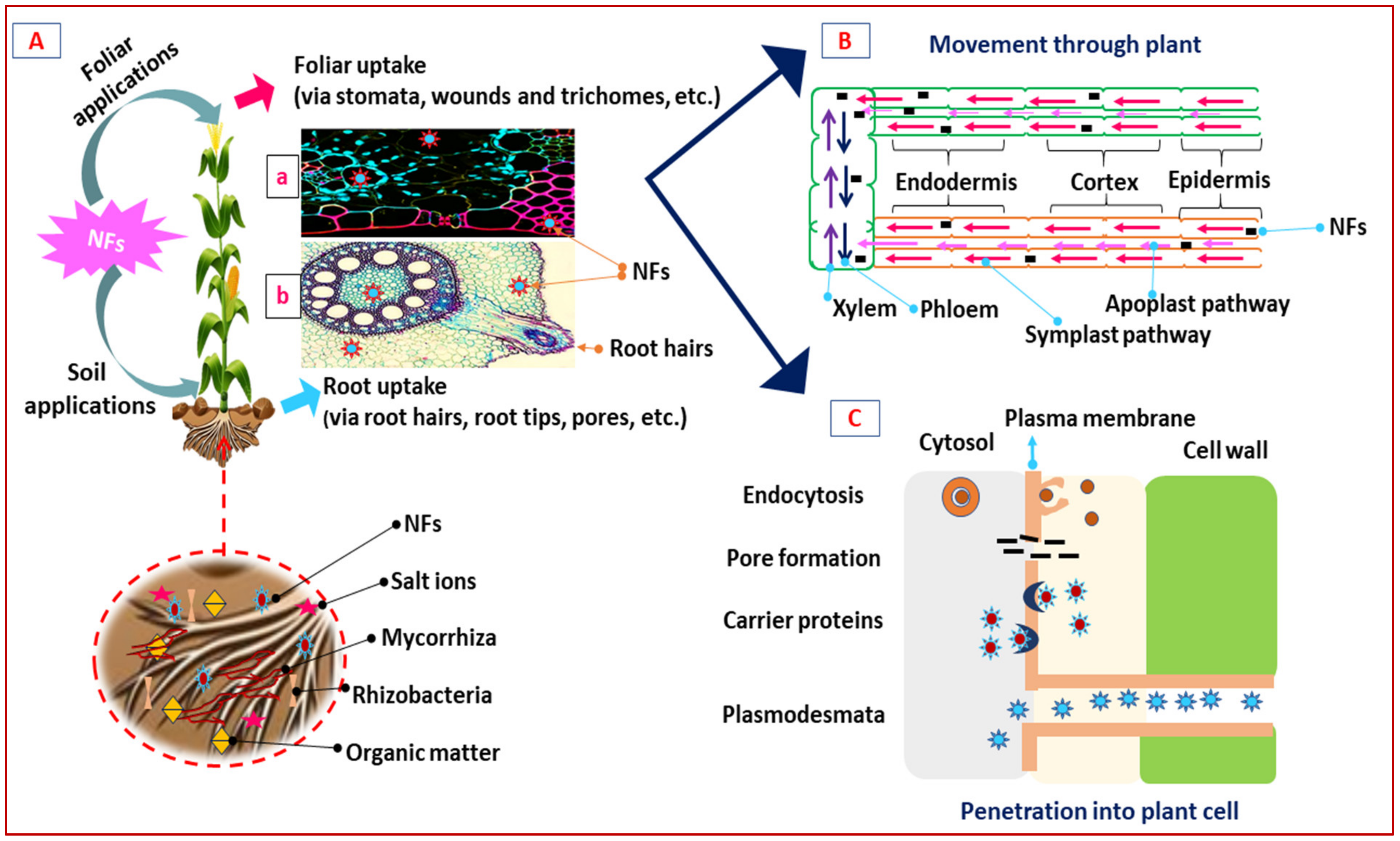
3.1. Uptake and Accumulation Mechanisms of Nano-Fertilizers from Soil to Plants
3.2. The Role of Nano-Fertilizers on Uptake of Water with Minerals
3.3. Impact of Nano-Fertilizer on Photosynthetic Leaf Gas Exchange Capacity
3.4. The Interactive Role of Nano-Fertilizers and Plant Growth—Biomass and Productivity
3.5. Influence of Nano-Fertilizer on the Regulation of Plant Hormones
3.6. Defense Mechanisms
3.7. Stimulation of Enzymatic and Non-Enzymatic Activities
3.8. Role of Nano-Fertilizers on the Expression of Stress-Responsive Genes
4. Long-Term Application of Nano-Fertilizers and Its Responses in Agriculture

{kind=link}
{kind=link}
{kind=link}
| NPs | Plant | Application Type | Concentration Range | Impacts | Source |
|---|---|---|---|---|---|
| nCeO2 | Barley (Hordeum vulgare L.) | Soil | 0–500 mg kg−1 soil | Improved plant performance, enhanced Ce accumulation in grains, and P, K, Ca, Mg, S, Cu, Fe, Zn, Mn, Al, amino acids, fatty acids, methionine, aspartic acid, threonine, tyrosine, arginine, and linolenic acid. | [163] |
| Wheat (Triticum aestivum L.) | Soil | 0–500 mg kg−1 soil | Enhanced overall plant fitness and productivity as compared to normal plants—increased Ce uptake in roots but no change in leaves, hull, and seeds. | [164] | |
| Wheat (Triticum aestivum L.) | Soil | 0–400 mg kg−1 soil | Reduced photosynthetic pigments and seed protein, antioxidant enzyme activities upregulated. No significant effects on plant biomass and productivity. | [165] | |
| Cucumber (Cucumis sativus L.) | Soil | 400 mg kg−1 soil | No change in starch level but changed carbohydrate pattern. Enhanced globulin and reduced glutelin content. | [166] | |
| Cilantro (Coriandrum sativum L.) | Soil | 0–500 mg kg−1 soil | Higher content was found in Ce, CAT in the stem, and APx in roots. | [167] | |
| nCuO | Tomato (Solanum lycopersicum L.) | Foliar | 50–500 ppm (particle size 50 nm) | Enhanced vitamin C, lycopene, ABTS, CAT, and SOD and reduced the APX and GPX activities. Increased Cu accumulation in tomato fruits. | [168] |
| Tomato (Solanum lycopersicum L.) | Soil | 0.02–10 ppm | Improved plant growth, development, productivity, and fruit quality. Enhanced the lycopene and antioxidant capacity. | [169] | |
| Cucumber (Cucumis sativus L.) | Hydroponic | 10–20 ppm | Increased ROS, phenolic components, amino acids, antioxidant enzymatic systems, and decreased citric acid level. | [170] | |
| Cucumber (Cucumis sativus L.) | Soil | 40 nm (particle size) | Fruit metabolites were changed as compared to control plants. Sugars and organic, amino, and fatty acids were enhanced. | [171] | |
| Tomato (Solanum lycopersicum Mill.) | Soil | 10–100 mM | Enhanced plant biomass and growth characteristics. Upregulated photosynthetic pigments, leaf gas exchange responses, and enzymatic activities. | [172] | |
| nCuO, nAl2O3, nTiO2 | Onion (Allium cepa L.) | Petriplate | 0–2000 µg mL−1 | Significantly affected the mitotic index. ROS activities enhanced in onion roots. Enzymatic activities increased, i.e., CAT and SOD in all applied NPs. | [173] |
| nCu/ kinetin | Kidney bean (Phaseolus vulgaris L.) | Soil | 50, 100 mg kg−1 soil | The chlorophyll content and nutrient elements, Ca, Mn, and P, were reduced and root Cu accumulation enhanced. | [174] |
| nCu–chitosan | Tomato (Solanum lycopersicum L.) | Soil | 0.3–0.015 M | Increased plant performance, productivity, stomatal conductance, and leaf CAT and fruit lycopene level. | [175] |
| nCu, nFe, nCo (Metal NPs) | Maize (Zea mays L.) | Soil irrigation | 3–5 ppm | Positively enhanced the seed germination frequency, time, and early growth, enzymatic activities, and metabolism of SOD in plant leaves to stress resistance capacity. | [176] |
| nSiO2 | Maize (Zea mays L.) | Hydroponic | 20–40 nm | Enhanced germination (%) rate, biomass, Si uptake, and nutrient uptake | [177] |
| Soybean (Glycine max L.) | Soil | 30–50 nm (particle size) | Reduced the toxic effects on plant performance and reduced Hg uptake in the epidermis and pericycle of the plant roots and leaves. Increase leaf gas exchange and enzymatic responses. | [178] | |
| Peregrina (Jatropha integerrima) | Foliar | 1–2 mM | Increased growth characteristics, biochemical profile, meanwhile reduced uptake of Na, Cl, total phenolics, and flavonoid contents in the plant leaves. | [179] | |
| Tomato (Solanum lycopersicum L.) | Petriplate | 0.05–2.5 ppm | The germination rate, root morphology, and biomass were significantly enhanced after NPs. Gene expression was upregulated, i.e., in AREB, TAS14, NCED3, CRK1, and RBOH1, APX2, MAPK2, ERF5, MAPK3, and DDF2 decreased. The genes are significantly associated to nSi in plant’s response to enhance stress resistance capacity. | [180] | |
| Mahaleb (Prunus mahaleb L.) | Soil irrigation | 10–100 ppm | Improved photosynthetic performance less impacted by stress when plants were pretreated with NPs at maximum treatment concentrations and upgraded nutritional level, i.e.,N, P, and K content. | [181] | |
| Faba bean (Vicia faba L.) | Soil | 1–3 mM | Improved seed germination rate and duration, plant length, leaf RWC biomass, seed quality, and productivity and nutritional element status, i.e., N, P, K, Ca, and Na. | [182] | |
| Cucumber (Cucumis sativus L.) | Foliar | 15–120 ppm | An enhancement in plant length, leaf number, areaexpansion, biomass, fruit weights, and quality as relative to control plants. | [183] | |
| Strawberry (Fragaria × ananassa) | Foliar and soil irrigation | 20–80 ppm | Significantly enhanced the nutritional content, such as K, Ca, Mg, Fe, Mn, and Si, in plant stem but no changes in Zn and Cu content. | [184] | |
| Sugarcane (Saccharum officinarum L.) | Foliar | 300 ppm | Enhanced photosynthetic efficiency, Fv/Fm variables, chlorophyll content, and PS II apparatus during cold stress conditions. | [185] | |
| Barley (Hordeum vulgare L.) | Soil | 12–250 ppm | Significantly enhanced plant growth performance, chlorophyll content, leaf gas exchange, osmolytes, antioxidative enzyme activities, cell membrane efficiency, and profile of metabolites. | [186] | |
| Wheat (Triticum aestivum L.) | Hydroponic | 10 µM | Alleviates harmful effects of UV radiation on plants. | [187] | |
| Marigold (Tagetes erecta L.) | Soil and foliar | 100–600 ppm | Enhanced biometrics, physiological, biochemical, and flower traits, i.e., fresh and dry mass of flower, flowering duration, and days taken to first bud initiation, etc. | [188] | |
| Biogenic amorphous silica (bASi) | - | Soil | 1–15% | Increases soil water holding capacity (SWHC). Soil management can be modified to increase bASI level, increasing available water content in soils, and to reduce water stress capacity for plant growth and development. | [189] |
| nFe2O3 | Soybean (Glycine max L.) | Foliar | 0.25–1 M | Enhanced leaf biomass with seed weight in comparison to normal plants. | [190] |
| Peanut (Arachis hypogaea L.) | Soil | 2–1000 ppm | Improved plant growth characteristics, root morphology, and productivity. Enhanced photosynthetic pigments, Chl index, plant hormones, enzymatic activities, and Fe uptake. | [191] | |
| Tomato (Solanum lycopersicum L.) | Hydroponic | 50–800 ppm | Improved germination of seeds, morphological traits, dry weight, and Fe uptake as compared to normal plants | [192] | |
| nFeS | Mustard (Brassica juncea L.) | Foliar | 2–10 ppm | Enhanced agronomic traits, photosynthetic pigments, membrane injury, nutrient assimilation, MDA, proline, and enzymatic activities versuswithout NP application. Activation of genes, i.e., rubiscosmall subunit (rubisco S), rubiscolarge subunit (rubisco L), glutamine synthetase (gs), and glutamate synthase (gogat). | [45] |
| nTiO2 | Cucumber (Cucumis sativus L.) | Soil | 0–750 mg kg−1 soil | Enhanced leaf greenness, CAT, and APx activity were reduced. Applied TiO2 increased Kand Plevels. | [193] |
| Barley (Hordeum vulgare L.) | Soil | 500–1000 mg kg−1 soil | Applied NPs found tostimulate plant performance by enhancing germination (%) as compared to normal and treated plants. | [39] | |
| Rice (Oryza sativa L.) | Soil | 0–750 mg kg−1 soil | Enhanced plant performance, P level in roots to grains. Upregulated the level of metabolites, i.e., amino acids, palmitic acids, and glycerol level in rice seeds. | [194] | |
| Tomato (Solanum lycopersicum L.) | Soil | 0–1000 mg kg−1 soil | Improved plant development uptake and accumulation of minerals. | [195] | |
| Tomato (Solanum lycopersicumL.) | Hydroponic | 0.5–4 M | nTiO2 improved plant growth and development (approx. 50%) and significantly enhanced the leaf gas exchange, i.e., quantum yield, performance index, photosynthetic pigments, and expression ofPSIgene compared to normal plant growth conditions. Enhanced expressions of glutathione synthase and glutathioneS-transferase in roots and leaves. Antioxidant activities increased in a dose-dependent method. Nutritional element significantly affected (P, S, Mg, and Fe content). | [196] | |
| Spinach (Spinacia oleracea L.) | - | 0.25% | Enhanced electron transport rate (ETR) and the oxygen-evolving rate (OER) of PS II, enzymatic responses, reduced ROS level. | [197] | |
| Tomato (Solanum lycopersicum L.) | Foliar | 0.05–0.2 M | Increased photosynthetic performance by regulating PS II energy dissipation and slightly reduced the Fv/Fm and electron transport rate in plant leaves. | [198] | |
| Wheat (Triticum vulgare L.) | Hydroponic | 5–40 ppm | No significant effects on plant performance. Leaf photosynthetic pigments were reduced with increasing NP levels. Increased nutrient uptake and accumulation except for K level. | [199] | |
| nTiO2-Activated carbon composite | Tomato (Solanum lycopersicum L.) and mungbean (Vigna radiates L.) | Foliar | 0–500 ppm | Appropriate NP concentrations can enhance the rate of seed germination and minimize the germination period in tomato and mungbean. | [200] |
| nFe3O4 | Cucumber (Cucumis sativus L.) | Hydroponic | 50–2000 ppm | Improved plant growth, development, yield, and enzymatic responses, i.e., SOD and POD. Applied NPs enhance/balance the proper nutrient management to overcome food security and safety. | [201] |
| Barley (Hordeum vulgare L.) | Hydroponic | 125–1000 ppm | Increase plant growth, biomass traits, photosynthetic pigments, total soluble protein, and chloroplasts frequency. No toxic effects were found during the excess dose of NPs. Excess NP application reduced the CAT and H2O2 activities, and alteration was found in the photosynthetic genes of plant leaves. | [202] | |
| nFe | Chili (Capsicum annuum L.) | Foliar | 0.002–2 mM L−1 | Low dose of nFe was noted to play positive role in plant growth and development. Enhanced chloroplast functional capacity and grana stacking. High dose of FeNPs found to have harmful effects on plants and can potentially stop the distribution of Fe nutrient. | [94] |
| nAg | Tomato (Solanum lycopersicum L.) | Seed | 0.05–2.5 ppm | Enhanced the rate of germination (%), root morphology, and plant output. The expression of genes was found to be upregulated (AREB, MAPK2, P5CS, and CRK1), and few genes were noted as downregulated (TAS14, DDF2, and ZFHD1). | [203] |
| Tomato (Solanum lycopersicum Mill.) | Soil irrigation | 10–40 ppm | Applied NPs enhanced the fruit characteristics and plant performance. | [204] | |
| Soybean (Glycine max (L.) Mell.) | Soil | 31.2–62.5 mg kg−1 soil | Negatively affected plant development and fixation of N. | [205] | |
| nZnO | Maize (Zea mays L.) | Foliar | 150–300 ppm | Enhanced maximum growth characteristics, physiological and biochemical activities during high pH treatment. | [206] |
| Mungbean (Vigna radiate L.) | Petriplate | 10–100 ppm | Enhanced germination rate, growth development, and nutritional elements. | [207] | |
| Tomato (Solanum lycopersicum Mill.) | Tissue culture | 15–30 ppm | ZnO NPs alleviated the adverse effects of plants. Lower dose was more appropriate than the higher. Various cultivars found different tolerance capacity for stress. | [208] | |
| Maize (Zea mays L.) | Foliar | 50–2000 ppm | Enhanced seed germination rate, seedling vigor index, biomass, productivity, and accumulation of Zn in grains. | [32] | |
| Peanut (Arachis hypogaea L.) | Soil irrigation | 0–1000 ppm | Increased vegetation growth rate, morphological traits, photosynthetic content, crop productivity, and overall plant performance. | [209] | |
| Sweet basil (Ocimum basilicumL.) | Foliar | 1000 ppm | Improved vegetative growth, development, essential oil productivity, biomass, and accumulation of Zn content. | [210] | |
| Peanut (Arachis hypogaea L.) | Soil | 100–500 ppm | Morphological, yield, and biochemical traits, such as plant length, biomass, and pod numbers/weight. Photosynthetic pigments, total phenols, reducing and total soluble sugar were positively affected by the NP treatment. | [211] | |
| Sorghum (Sorghum bicolor L.) | Soil and foliar | 6 mg kg−1 soil | Enhanced plant performance and yield component, uptake of N and K elements, improved grain nutrient profile and NUE as compared to normal plants. | [212] | |
| nZn–chitosan | Wheat (Triticum durum) | Soil and foliar | 20 mg g−1 soil (w/w) | Increased Zn accumulation in the plants cultivated under Zn-deficient arable land. | [213] |
| nChitosan | Barley (Hordeum vulgareL.) | Soil and foliar | 10–100 ppm | Significantly enhanced the leaf areaexpansion, leaf greenness (Chl index), number of seeds/spikes, productivity, and harvest index relative to normal plants. nChitosan enhanced the LRWC, grain weight, grain protein, proline, and CAT and SOD activity. | [214] |
| nChitosan-NPK | Wheat (Triticum aestivum L.) | Foliar | 500, 60, and 400 ppm (N, P, and K), 10, 25, and 100% | Enhanced growth, yield, and nutritional status as compared to normal plants. | [215] |
| nChitosan | Barley (Hordeum vulgare L.) | Soil and foliar | 30–90 ppm | Positively enhanced the growth parameters, leaf chlorophyll index, RWC, yield, and biochemical activities. | [214] |
| nZ (Zein NPs) | Sugarcane (Saccharum spp.) | Hydroponic | 0.88–1.75 mg mL−1 | Uptake of significant amount of ZNPs in cane roots and the presence of Z particles in the epidermis and endodermis in the roots system of the sugarcane plant. Increased nutrient uptake in the plant system. | [216] |
| nAu | Thale cress (Arabidopsis thaliana L.) | Foliar | 10–80 µg mL−1 | Increased seed germination (%), growth, free radical scavenging responses. Potential approach to increase the seed productivity of plants. | [217] |
| Brown mustard (Brassica juncea L.) | Foliar | 0–100 ppm | Significantly enhanced the growth, biomass parameters, and total sugar level. Leafarea expansion was increased, but the mean area not affected. | [218] | |
| Mn3O4 | Cucumber (Cucumis sativus L.) | Foliar | 1–5 mg plant−1 | Significantly enhanced plant development, chlorophyll content, photosynthetic responses, and plant biomass. Increased endogenous antioxidative defense mechanisms. | [219] |
| nUrea modified with hydroxyapatite | Almond (Prunus dulcis L.) | Soil irrigation | 25–100% | Applied NPs enhanced seed germination rate, plant height, perimeter, elongation of secondary and primary roots/plant, and the number of secondary roots, increasing seed moisture status. | [220] |
5. Conclusions and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pouratashi, M.; Iravani, H. Farmers’ knowledge of integrated pest management and learning style preferences: Implications for information delivery. Int. J. Pest Manag. 2012, 58, 347–353. [Google Scholar] [CrossRef]
- Mittal, D.; Kaur, G.; Singh, P.; Yadav, K.; Ali, S.A. Nanoparticle-Based Sustainable Agriculture and Food Science: Recent Advances and Future Outlook. Front. Nanotechnol. 2020, 2, 10. [Google Scholar] [CrossRef]
- Hussein, M.M.; Abou-Baker, N.H. The contribution of nano-zinc to alleviate salinity stress on cotton plants. R. Soc. Open Sci. 2018, 5, 171809. [Google Scholar] [CrossRef] [Green Version]
- Robroek, B.J.M.; Jassey, V.E.J.; Beltman, B.; Hefting, M. Diverse fen plant communities enhance carbon-related multifunctionality, but do not mitigate negative effects of drought. R. Soc. Open Sci. 2017, 4, 170449. [Google Scholar] [CrossRef] [Green Version]
- Bindraban, P.S.; Dimkpa, C.O.; Angle, S.; Rabbinge, R. Unlocking the multiple public good services from balanced fertilizers. Food Secur. 2018, 10, 273–285. [Google Scholar] [CrossRef]
- Mandal, D. Nanofertilizer and its application in horticulture. J. Appl. Hortic. 2021, 23, 70–77. [Google Scholar] [CrossRef]
- Fellet, G.; Pilotto, L.; Marchiol, L.; Braidot, E. Tools for Nano-Enabled Agriculture: Fertilizers Based on Calcium Phosphate, Silicon, and Chitosan Nanostructures. Agronomy 2021, 11, 1239. [Google Scholar] [CrossRef]
- Elmer, W.H.; White, J.C. The use of metallic oxide nanoparticles to enhance growth of tomatoes and eggplants in disease infested soil or soilless medium. Environ. Sci. Nano 2016, 3, 1072–1079. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, X.; Zhao, J.; Liu, X.; Feng, W.; White, J.C.; Xing, B. Xylem- and Phloem-Based Transport of CuO Nanoparticles in Maize (Zea mays L.). Environ. Sci. Technol. 2012, 46, 4434–4441. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Adisa, I.O.; Pullagurala, V.L.R.; Peralta-Videa, J.R.; Dimkpa, C.O.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Recent advances in nano-enabled fertilizers and pesticides: A critical review of mechanisms of action. Environ. Sci. Nano 2019, 6, 2002–2030. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Feizi, M.; Kumari, A.; Khan, M.; Mandzhieva, S.; Sushkova, S.; El-Ramady, H.; Verma, K.K.; Singh, A.; et al. Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth. Biology 2021, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Raliya, R.; Saharan, V.; Dimkpa, C.; Biswas, P. Nanofertilizer for Precision and Sustainable Agriculture: Current State and Future Perspectives. J. Agric. Food Chem. 2017, 66, 6487–6503. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, D.; Wilson, A. World population agriculture and malnutrition. World Watch 2004, 17, 22–25. [Google Scholar]
- Renner, A.; Cadillo-Benalcazar, J.J.; Benini, L.; Giampietro, M. Environmental pressure of the European agricultural system: Anticipating the biophysical consequences of internalization. Ecosyst. Serv. 2020, 46, 101195. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.M.; Gilbertson, L.M. Rational Ligand Design to Improve Agrochemical Delivery Efficiency and Advance Agriculture Sustainability. ACS Sustain. Chem. Eng. 2018, 6, 13599–13610. [Google Scholar] [CrossRef]
- F2F. Farm to Fork Strategy. 2020. Available online: https://ec.europa.eu/food/farm2fork_en (accessed on 3 August 2021).
- OECD-FAO. Agricultural Outlook 2020–2029; OECD Publishing: Paris, France; FAO: Rome, Italy, 2020. [Google Scholar]
- Faizan, M.; Rajput, V.; Al-Khuraif, A.; Arshad, M.; Minkina, T.; Sushkova, S.; Yu, F. Effect of Foliar Fertigation of Chitosan Nanoparticles on Cadmium Accumulation and Toxicity in Solanum lycopersicum. Biology 2021, 10, 666. [Google Scholar] [CrossRef]
- Gogos, A.; Knauer, K.; Bucheli, T.D. Nanomaterials in Plant Protection and Fertilization: Current State, Foreseen Applications, and Research Priorities. J. Agric. Food Chem. 2012, 60, 9781–9792. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Singh, A.; Minkina, T.; Rawat, S.; Mandzhieva, S.; Sushkova, S.; Shuvaeva, V.; Nazarenko, O.; Rajput, P.; Komariah; et al. Nano-Enabled Products: Challenges and Opportunities for Sustainable Agriculture. Plants 2021, 10, 2727. [Google Scholar] [CrossRef]
- Singh, H.; Sharma, A.; Bhardwaj, S.K.; Arya, S.K.; Bhardwaj, N.; Khatri, M. Recent advances in the applications of nano-agrochemicals for sustainable agricultural development. Environ. Sci. Process. Impacts 2021, 23, 213–239. [Google Scholar] [CrossRef] [PubMed]
- Lowry, G.V.; Avellan, A.; Gilbertson, L.M. Opportunities and challenges for nanotechnology in the agri-tech revolution. Nat. Nanotechnol. 2019, 14, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Bandala, E.R.; Berli, M. Engineered nanomaterials (ENMs) and their role at the nexus of Food, Energy, and Water. Mater. Sci. Energy Technol. 2019, 2, 29–40. [Google Scholar] [CrossRef]
- Fraceto, L.F.; Grillo, R.; de Medeiros, G.A.; Scognamiglio, V.; Rea, G.; Bartolucci, C. Nanotechnology in agriculture: Which innovation potential does it have? Front. Environ. Sci. 2016, 4, 20. [Google Scholar] [CrossRef]
- Saharan, V.; Kumaraswamy, R.V.; Choudhary, R.C.; Kumari, S.; Pal, A.; Raliya, R.; Biswas, P. Cu-Chitosan Nanoparticle Mediated Sustainable Approach to Enhance Seedling Growth in Maize by Mobilizing Reserved Food. J. Agric. Food Chem. 2016, 64, 6148–6155. [Google Scholar] [CrossRef]
- Wiesner, M.R.; Lowry, G.V.; Alvarez, P.; Dionysiou, D.; Biswas, P. Assessing the Risks of Manufactured Nanomaterials. Environ. Sci. Technol. 2006, 40, 4336–4345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raliya, R.; Biswas, P. Environmentally benign bio-inspired synthesis of Au nanoparticles, their self-assembly and agglomeration. RSC Adv. 2015, 5, 42081–42087. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Bindraban, P.S.; Fugice, J.; Agyin-Birikorang, S.; Singh, U.; Hellums, D. Composite micronutrient nanoparticles and salts decrease drought stress in soybean. Agron. Sustain. Dev. 2017, 37, 5. [Google Scholar] [CrossRef] [Green Version]
- Subbaiah, L.V.; Prasad, T.N.V.K.V.; Krishna, T.G.; Sudhakar, P.; Reddy, B.R.; Pradeep, T. Novel Effects of Nanoparticulate Delivery of Zinc on Growth, Productivity, and Zinc Biofortification in Maize (Zea mays L.). J. Agric. Food Chem. 2016, 64, 3778–3788. [Google Scholar] [CrossRef]
- Li, S.X.; Wang, Z.H.; Miao, Y.F.; Li, S.Q. Soil organic nitrogen and its contribution to crop production. J. Integr. Agric. 2014, 13, 2061–2080. [Google Scholar] [CrossRef]
- Dubey, A.; Mailapalli, D.R. Nanofertilisers, nanopesticides, nanosensors of pest and nanotoxicity in agriculture. In Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer: Cham, Switzerland, 2016; Volume 19, pp. 307–330. [Google Scholar] [CrossRef]
- Naderi, M.R.; Shahraki, A.D. Nanofertilizers and their roles in sustainable agriculture. Intl. J. Agr. Crop. Sci. 2013, 5, 2229–2232. [Google Scholar]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef]
- Chhipa, H. Nanofertilizers and nanopesticides for agriculture. Environ. Chem. Lett. 2017, 15, 15–22. [Google Scholar] [CrossRef]
- Jyothi, T.; Hebsur, N. Effect of nanofertilizers on growth and yield of selected cereals—A review. Agric. Rev. 2017, 38, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Marchiol, L.; Mattiello, A.; Pošćić, F.; Fellet, G.; Zavalloni, C.; Carlino, E.; Musetti, R. Changes in physiological and agronomical parameters of barley (Hordeum vulgare) exposed to cerium and titanium dioxide nanoparticles. Int. J. Environ. Res. Public Health 2016, 13, 332. [Google Scholar] [CrossRef] [Green Version]
- Peralta-Videa, J.R. Are nanomaterials a real solution for sustainable agriculture? Acta Sci. Agric. 2018, 2, 16–17. [Google Scholar]
- Chugh, G.; Siddique, K.; Solaiman, Z. Nanobiotechnology for Agriculture: Smart Technology for Combating Nutrient Deficiencies with Nanotoxicity Challenges. Sustainability 2021, 13, 1781. [Google Scholar] [CrossRef]
- Kah, M.; Tufenkji, N.; White, J.C. Nano-enabled strategies to enhance crop nutrition and protection. Nat. Nanotechnol. 2019, 14, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A critical evaluation of nanopesticides and nanofertilizers against their conventional analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Pullagurala, V.L.R.; Adisa, I.O.; Rawat, S.; Kim, B.; Barrios, A.C.; Medina-Velo, I.A.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Finding the conditions for the beneficial use of ZnO nanoparticles towards plants-A review. Environ. Pollut. 2018, 241, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Rawat, M.; Nayan, R.; Negi, B.; Zaidi, M.; Arora, S. Physio-biochemical basis of iron-sulfide nanoparticle induced growth and seed yield enhancement in B. juncea. Plant Physiol. Biochem. 2017, 118, 274–284. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munne-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef]
- Sharifi, M.; Faryabi, K.; Talaei, A.J.; Shekha, M.S.; Ale-Ebrahim, M.; Salihi, A.; Nanakali, N.M.Q.; Aziz, F.M.; Rasti, B.; Hasan, A.; et al. Antioxidant properties of gold nanozyme: A review. J. Mol. Liq. 2020, 297, 112004. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon dioxide mitigates the adverse effects of salt stress on Cucurbita pepo L. Environ. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef] [PubMed]
- Mingyu, S.; Chao, L.; Chunxiang, Q.; Lei, Z.; Liang, C.; Hao, H.; Xiaoqing, L.; Xiao, W.; Fashui, H. Nano-Anatase Relieves the Inhibition of Electron Transport Caused by Linolenic Acid in Chloroplasts of Spinach. Biol. Trace Elem. Res. 2008, 122, 73–81. [Google Scholar] [CrossRef]
- Khalkhal, K.; Lajayer, B.A.; Ghorbanpour, M. An overview on the effect of soil physicochemical properties on the immobilization of biogenic nanoparticles. In Biogenic Nano-Particles and Their Use in Agro-Ecosystems; Springer Science and Business Media LLC: Singapore, 2020; pp. 133–160. [Google Scholar]
- Li, M.; Wang, P.; Dang, F.; Zhou, D.-M. The transformation and fate of silver nanoparticles in paddy soil: Effects of soil organic matter and redox conditions. Environ. Sci. Nano 2017, 4, 919–928. [Google Scholar] [CrossRef]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Claassen, N.; Syring, K.M.; Jungk, A. Verification of a mathematical model by simulating potassium uptake from soil. Plant Soil 1986, 95, 209–220. [Google Scholar] [CrossRef]
- Thornley, J.H.; Johnson, I.R. Plant and Crop Modeling—A Mathematical Approach to Plant and Crop Physiology; Clarendon Press: Oxford, UK, 1990; p. 660. [Google Scholar]
- Minchin, P.E.H.; Thorpe, M.R.; Farrar, J.F. A Simple Mechanistic Model of Phloem Transport which Explains Sink Priority. J. Exp. Bot. 1993, 44, 947–955. [Google Scholar] [CrossRef]
- Payvandi, S.; Daly, K.R.; Zygalakis, K.; Roose, T. Mathematical Modelling of the Phloem: The Importance of Diffusion on Sugar Transport at Osmotic Equilibrium. Bull. Math. Biol. 2014, 76, 2834–2865. [Google Scholar] [CrossRef]
- Pitambara; Archana; Shukla, Y.M. Nanofertilizers: A Recent Approach in Crop Production. Nanotechnol. Agric. Crop Prod. Prot. 2019, 25–58. [Google Scholar] [CrossRef]
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of Nanoparticles with Edible Plants and Their Possible Implications in the Food Chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Corredor, E.; Testillano, P.S.; Coronado, M.-J.; Gonzalez-Melendi, P.; Fernández-Pacheco, R.; Marquina, C.; Ibarra, M.R.; de la Fuente, J.M.; Rubiales, D.; Perez-de-Laque, A. Nanoparticlepenetration and transport in living pumpkin plants: In Situ subcellular identification. BMC Plant Biol. 2009, 9, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juárez-Maldonado, A.; Ortega-Ortíz, H.; Morales-Díaz, A.B.; González-Morales, S.; Morelos-Moreno, Á.; Sandoval-Rangel, A.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Nanoparticles and Nanomaterials as Plant Biostimulants. Int. J. Mol. Sci. 2019, 20, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotze, E.; Phenrat, T.; Lowry, G.V. Nanoparticle Aggregation: Challenges to Understanding Transport and Reactivity in the Environment. J. Environ. Qual. 2010, 39, 1909–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, M.; Pagay, V.; Stroock, A.D. Impact of Electroviscosity on the Hydraulic Conductance of the Bordered Pit Membrane: A Theoretical Investigation. Plant Physiol. 2013, 163, 999–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Himmel, M.E.; Ding, S.-Y. Visualizing chemical functionality in plant cell walls. Biotechnol. Biofuels 2017, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meychik, N.; Nikolaeva, J.; Yermakov, I. Ion Exchange Properties of the Root Cell Walls Isolated from the Halophyte Plants (Suaedaaltissima L.) Grown Under Conditions of Different Salinity. Plant Soil 2005, 277, 163. [Google Scholar] [CrossRef]
- Zhu, Z.-J.; Wang, H.; Yan, B.; Zheng, H.; Jiang, Y.; Miranda, O.R.; Rotello, V.M.; Xing, B.; Vachet, R.W. Effect of Surface Charge on the Uptake and Distribution of Gold Nanoparticles in Four Plant Species. Environ. Sci. Technol. 2012, 46, 12391–12398. [Google Scholar] [CrossRef]
- Liu, M.; Feng, S.; Ma, Y.; Xie, C.; He, X.; Ding, Y.; Zhang, J.; Luo, W.; Zheng, L.; Chen, D.; et al. Influence of Surface Charge on the Phytotoxicity, Transformation, and Translocation of CeO2 Nanoparticles in Cucumber Plants. ACS Appl. Mater. Interfaces 2019, 11, 16905–16913. [Google Scholar] [CrossRef] [PubMed]
- Pirvulescua, A.; Salaa, F.; Boldea, M. Variation of chlorophyll content in sunflower under the influence of magnetic nanofluids. In Proceedings of the International Conference of Numerical Analysis and Applied Mathematics 2014 (ICNAAM-2014), Rhodes, Greece, 22–28 September 2014. [Google Scholar]
- Morteza, E.; Moaveni, P.; Farahani, H.A.; Kiyani, M. Study of photosynthetic pigments changes of maize (Zea mays L.) under nano TiO2 spraying at various growth stages. SpringerPlus 2013, 2, 247. [Google Scholar] [CrossRef] [Green Version]
- Janmohammadi, M.; Amanzadeh, T.; Sabaghnia, N.; Dashti, S. Impact of foliar application of nano micronutrient fertilizers and titanium dioxide nanoparticles on the growth and yield components of barley under supplemental irrigation. Acta Agric. Slov. 2016, 107, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Šebesta, M.; Kolenčík, M.; Sunil, B.R.; Illa, R.; Mosnáček, J.; Ingle, A.P.; Urík, M. Field Application of ZnO and TiO2 Nanoparticles on Agricultural Plants. Agronomy 2021, 11, 2281. [Google Scholar] [CrossRef]
- Gao, F.; Hong, F.; Liu, C.; Zheng, L.; Su, M.; Wu, X.; Yang, F.; Wu, C.; Yang, P. Mechanism of Nano-anatase TiO2 on Promoting Photosynthetic Carbon Reaction of Spinach. Biol. Trace Element Res. 2006, 111, 239–254. [Google Scholar] [CrossRef]
- Rezaei, M.; Abbasi, H. Foliar application of nanochelate and non-nanochelate of zinc on plant resistance physiological processes in cotton (Gossipiumhirsutum L.). Iran. J. Plant Physiol. 2014, 4, 1137–1144. [Google Scholar]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; GolezaniGhassemi, M. Changes in antioxidant enzymes activity and plant performance by salinity stress and zinc application in soybean (Glycin max L.). Plant Omics J. 2012, 5, 60–67. [Google Scholar]
- Tarafdar, J.C.; Raliya, R.; Mahawar, H.; Rathore, I. Development of Zinc Nanofertilizer to Enhance Crop Production in Pearl Millet (Pennisetumamericanum). Agric. Res. 2014, 3, 257–262. [Google Scholar] [CrossRef]
- Vafa, Z.N.; Sirousmehr, A.R.; Ghanbari, A.; Khammari, E.; Falahi, N. Effect of nano-zinc and humic acid in quantitative and qualitative characteristics of savory (Saturejahortensis L.). Int. J. Bio Sci. 2015, 6, 124–136. [Google Scholar]
- Benzon, H.R.L.; Rubenecia, M.R.U.; Ultra, V.U.; Lee, S.C. Nano-fertilizer affects the growth, development, and chemical properties of rice. Int. J. Agron. Agric. Res. 2015, 7, 105–117. [Google Scholar]
- Gao, J.; Xu, G.; Qian, H.; Liu, P.; Zhao, P.; Hu, Y. Effects of nanoTiO2 on photosynthetic characteristics of Ulmus elongate seedlings. Environ. Pollut. 2013, 176, 63–70. [Google Scholar] [CrossRef]
- Younis, A.; Khattab, H.; Emam, M. Impacts of silicon and silicon nanoparticles on leaf ultrastructure and TaPIP1 and TaNIP2 gene expressions in heat stressed wheat seedlings. Biol. Plant. 2020, 64, 343–352. [Google Scholar] [CrossRef]
- Kondal, R.; Kalia, A.; Krejcar, O.; Kuca, K.; Sharma, S.P.; Luthra, K.; Dheri, G.S.; Vikal, Y.; Taggar, M.S.; Abd-Elsalam, K.A.; et al. Chitosan-Urea Nanocomposite for Improved Fertilizer Applications: The Effect on the Soil Enzymatic Activities and Microflora Dynamics in N Cycle of Potatoes (Solanum tuberosum L.). Polymers 2021, 13, 2887. [Google Scholar] [CrossRef]
- Saharan, V.; Sharma, G.; Yadav, M.; Choudhary, M.K.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P. Synthesis and In Vitro antifungal efficacy of Cu–chitosan nanoparticles against pathogenic fungi of tomato. Int. J. Biol. Macromol. 2015, 75, 346–353. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Shome, S.; Tewari, S.; Bhattacharya, M.K.; Panda, S.K. Effect of Zn nano-particles on growth responses of rice. In Nanotechnology: Novel Perspectives and Prospects; Singh, B., Kaushik, A., Mehta, S.K., Tripathi, S.K., Eds.; McGraw Hill Education: New Delhi, India, 2015; pp. 508–512. [Google Scholar]
- Torabian, S.; Zahedi, M.; Khoshgoftarmanesh, A. Effect of foliar spray of zinc oxide on some antioxidant enzymes activity of sunflower under salt stress. J. Agric. Sci. Technol. 2016, 18, 1013–1025. [Google Scholar]
- Babaei, K.; Sharifi, R.S.; Pirzad, A.; Khalilzadeh, R. Effects of bio fertilizer and nano Zn-Fe oxide on physiological traits, antioxidant enzymes activity and yield of wheat (Triticum aestivum L.) under salinity stress. J. Plant Interact. 2017, 12, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Farnia, A.; Omidi, M.M.; Farnia, A. Effect of nano-zinc chelate and nano-biofertilizer on yield and yield components of maize (Zea Mays L.) under water stress condition. Ind. J. Nat. Sci. 2015, 5, 4614–4646. [Google Scholar]
- Huang, S.; Wang, L.; Liu, L.; Hou, Y.; Li, L. Nanotechnology in agriculture, livestock, and aquaculture in China. A review. Agron. Sustain. Dev. 2015, 35, 369–400. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Fan, X.; Li, X.; Zhang, Z.; Sun, L.; Fu, Z.; Lavoie, M.; Pan, X.; Qian, H. Distinct physiological and molecular responses in Arabidopsis thaliana exposed to aluminum oxide nanoparticles and ionic aluminum. Environ. Pollut. 2017, 228, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Khodakovskaya, M.V.; de Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon Nanotubes Induce Growth Enhancement of Tobacco Cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef]
- Acharya, P.; Jayaprakasha, G.K.; Crosby, K.M.; Jifon, J.L.; Patil, B.S. Nanoparticle-Mediated Seed Priming Improves Germination, Growth, Yield, and Quality of Watermelons (Citrullus lanatus) at multi-locations in Texas. Sci. Rep. 2020, 10, 5037. [Google Scholar] [CrossRef] [Green Version]
- Anand, K.V.; Anugraga, A.; Kannan, M.; Singaravelu, G.; Govindaraju, K. Bio-engineered magnesium oxide nanoparticles as nano-priming agent for enhancing seed germination and seedling vigour of green gram (Vigna radiata L.). Mater. Lett. 2020, 8, 127792. [Google Scholar] [CrossRef]
- Bhat, J.A.; Rajora, N.; Raturi, G.; Sharma, S.; Dhiman, P.; Sanand, S.; Shivaraj, S.M.; Sonah, H.; Deshmukh, R. Silicon nanoparticles (SiNPs) in sustainable agriculture: Major emphasis on the practicality, efficacy and concerns. Nanoscale Adv. 2021, 3, 4019. [Google Scholar] [CrossRef]
- Rafi, M.M.; Epstein, E.; Falk, R.H. Silicon deprivation causes physical abnormalities in wheat (Triticum aestivum L). J. Plant Physiol. 1997, 151, 497–501. [Google Scholar] [CrossRef]
- Larue, C.; Castillo-Michel, H.; Sobanska, S.; Cécillon, L.; Bureau, S.; Barthès, V.; Ouerdane, L.; Carrière, M.; Sarret, G. Foliar exposure of the crop Lactuca sativa to silver nanoparticles: Evidence for internalization and changes in Ag speciation. J. Hazard. Mater. 2014, 264, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Boykov, I.N.; Shuford, E.; Zhang, B. Nanoparticle titanium dioxide affects the growth and microRNA expression of switchgrass (Panicum virgatum). Genomics 2019, 111, 450–456. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, Y.; Li, H.; Lu, J.; Zhao, H.; Liu, M.; Nechitaylo, G.S.; Glushchenko, N.N. New insights into the cellular responses to iron nanoparticles in Capsicum annuum. Sci. Rep. 2018, 8, 3228. [Google Scholar] [CrossRef]
- Rajput, V.; Minkina, T.; Fedorenko, A.; Sushkova, S.; Mandzhieva, S.; Lysenko, V.; Duplii, N.; Fedorenko, G.; Dvadnenko, K.; Ghazaryan, K. Toxicity of copper oxide nanoparticles on spring barley (Hordeum sativum distichum). Sci. Total Environ. 2018, 645, 1103–1113. [Google Scholar] [CrossRef]
- Raja, K.; Sowmya, R.; Sudhagar, R.; Moorthy, P.S.; Govindaraju, K.; Subramanian, K. Biogenic ZnO and Cu nanoparticles to improve seed germination quality in blackgram (Vigna mungo). Mater. Lett. 2019, 235, 164–167. [Google Scholar] [CrossRef]
- Rahman, M.; Chakraborty, A.; Mazumdar, S.; Nandi, N.; Bhuiyan, M.; Alauddin, S.; Khan, I.; Hossain, M.J. Effects of poly(vinylpyrrolidone) protected platinum nanoparticles on seed germination and growth performance of Pisum sativum. Nano-Struct. Nano-Objects 2020, 21, 100408. [Google Scholar] [CrossRef]
- Li, R.; He, J.; Xie, H.; Wang, W.; Bose, S.K.; Sun, Y.; Hu, J.; Yin, H. Effects of chitosan nanoparticles on seed germination and seedling growth of wheat (Triticum aestivum L.). Int. J. Biol. Macromol. 2019, 126, 91–100. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.M.M.; Hasaneen, M.N.A.; Aya, M.O. Foliar application of nano chitosan NPK fertilizer improves the yield of wheat plants grown on two different soils. Egypt J. Exp. Biol. 2018, 14, 63–72. [Google Scholar]
- Jaberzadeh, A.; Moaveni, P.; Moghadam, H.R.T.; Zahedi, H. Influence of Bulk and Nanoparticles Titanium Foliar Application on some Agronomic Traits, Seed Gluten and Starch Contents of Wheat Subjected to Water Deficit Stress. Not. Bot. HortiAgrobot. Cluj-Napoca 2013, 41, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Ghafariyan, M.H.; Malakouti, M.J.; Dadpour, M.R.; Stroeve, P.; Mahmoudi, M. Effects of Magnetite Nanoparticles on Soybean Chlorophyll. Environ. Sci. Technol. 2013, 47, 10645–10652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quary, F.X.; Leenhardt, F.; Remesy, C. Genetic variability and stability of grain Mg, Zn and Fe concentration in bread wheat. Eur. J. Agron. 2006, 25, 177–185. [Google Scholar]
- Mekkdad, A.A.A. Response of peanut nitrogen fertilizer levels and foliar zinc spraying rates in newly reclaimed sandy soils. J. Plant Prod. Mansoura Univ. 2017, 8, 153–159. [Google Scholar] [CrossRef]
- El-Metwally, I.M.; Doaa, M.R.; Abo-Basha, A.E.A.M.; Abd El-Aziz, M. Response of peanut plants to different foliar applications of nano-iron, manganese and zinc under sandy soil conditions. Middle East J. Appl. Sci. 2018, 8, 474–482. [Google Scholar]
- Mehrangiz, J.M.; Sirous, B.; Ebrahim, A. Study the effect of foliar application of nano chelate molybdenum fertilizer on the yield and yield components of peanut. Egypt. Acad. J. Biol. Sci. H. Bot. 2014, 6, 37–40. [Google Scholar]
- Anonymous. Annual Report; ICAR-CICR: Nagapur, Maharashtra, India, 2016; p. 32. [Google Scholar]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Mehmood, S.S.; Tabassum, J.; Batool, R. Targeting plant hormones to develop abiotic stress resistance in wheat. In Wheat Production in Changing Environments; Springer: Singapore, 2019; p. 557. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Hussain, H.I.; Yi, Z.; Rookes, J.E.; Kong, L.; Cahill, D.M. Delivery of Abscisic Acid to Plants Using Glutathione Responsive Mesoporous Silica Nanoparticles. J. Nanosci. Nanotechnol. 2018, 18, 1615–1625. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Benková, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vankova, R.; Landa, P.; Podlipna, R.; Dobrev, P.I.; Přerostová, S.; Langhansova, L.; Gaudinova, A.; Motkova, K.; Knirsch, V.; Vanek, T. ZnO nanoparticle effects on hormonal pools in Arabidopsis thaliana. Sci. Total Environ. 2017, 593, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Salachna, P.; Mizielińska, M.; Płoszaj-Witkowska, B.; Jaszczak, A. Zinc Oxide Nanoparticles Enhanced Biomass and Zinc Content and Induced Changes in Biological Properties of Red Perilla frutescens. Materials 2021, 14, 6182. [Google Scholar] [CrossRef]
- El-Tohamy, W.A.; El-Greadly, N.H.M. Physiological responses, growth, yield and quality of snap bean in response to foliar application of yeast, vitamin E and zinc under sandy soil conditions. Aust. J. Basic Appl. Sci. 2007, 1, 249–299. [Google Scholar]
- Li, M.; Wang, S.; Tian, X.; Zhao, J.; Li, H.; Guo, C.; Chen, Y.; Zhao, A. Zn distribution and bioavailability in whole grain and grain fractions of winter wheat as affected by applications of soil N and foliar Zn combined with N or P. J. Cereal Sci. 2015, 61, 26–32. [Google Scholar] [CrossRef]
- Burman, U.; Saini, M.; Kumar, P. Effect of zinc oxide nanoparticles on growth and antioxidant system of chickpea seedlings. Toxicol. Environ. Chem. 2013, 95, 605–612. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic bio-fortification. Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C. ZnO nanoparticle biosynthesis and its effect on phosphorusmobilizing enzyme secretion and gum contents in clusterbean (Cyamopsistetragonoloba L.). Agric. Res. 2013, 20, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995. [Google Scholar]
- Wang, P.; Lombi, E.; Zhao, F.-J.; Kopittke, P. Nanotechnology: A New Opportunity in Plant Sciences. Trends Plant Sci. 2016, 21, 699–712. [Google Scholar] [CrossRef]
- Zahra, Z.; Arshad, M.; Rafique, R.; Mahmood, A.; Habib, A.; Qazi, I.A.; Khan, S.A. Metallic Nanoparticle (TiO2 and Fe3O4) Application Modifies Rhizosphere Phosphorus Availability and Uptake by Lactuca sativa. J. Agric. Food Chem. 2015, 63, 6876–6882. [Google Scholar] [CrossRef]
- Hong, F.; Yang, F.; Liu, C.; Gao, Q.; Wan, Z.; Gu, F.; Wu, C.; Ma, Z.; Zhou, J.; Yang, P. Influences of nano-TiO2 on the chloroplast aging of spinach under light. Biol. Trace Elem. Res. 2005, 104, 249–260. [Google Scholar] [CrossRef]
- Hong, F.; Zhou, J.; Liu, C.; Yang, F.; Wu, C.; Zheng, L.; Yang, P. Effect of nano-TiO2 on photochemical reaction of chloroplasts of spinach. Biol. Trace Elem. Res. 2005, 105, 269–279. [Google Scholar] [CrossRef]
- Linglan, M.; Chao, L.; Chunxiang, Q.; Sitao, Y.; Jie, L.; Fengqing, G.; Fashui, H. Rubisco Activase mRNA Expression in Spinach: Modulation by Nanoanatase Treatment. Biol. Trace Elem. Res. 2008, 122, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Gao, Y.; Wu, H.; Hou, W.; Zhang, C.; Ma, H. Physiological effect of anatase TiO2 nanoparticles on Lemna minor. Environ. Toxicol. Chem. 2012, 31, 2147–2152. [Google Scholar] [CrossRef] [PubMed]
- Feizi, H.; Rezvani Moghaddam, P.; Shahtahmassebi, N.; Fotovat, A. Impact of bulk and nanosized titanium dioxide (TiO2) on wheat seed germination and seedling growth. Biol. Trace Elem. Res. 2012, 146, 101–106. [Google Scholar] [CrossRef]
- Wang, W.-N.; Tarafdar, J.C.; Biswas, P. Nanoparticle synthesis and delivery by an aerosol route for watermelon plant foliar uptake. J. Nanoparticle Res. 2013, 15, 1417. [Google Scholar] [CrossRef]
- Raliya, R.; Biswas, P.; Tarafdar, J. TiO2 nanoparticle biosynthesis and its physiological effect on mung bean (Vigna radiata L.). Biotechnol. Rep. 2015, 5, 22–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarafdar, A.; Raliya, R.; Wang, W.-N.; Biswas, P.; Tarafdar, J.C. Green Synthesis of TiO2 Nanoparticle Using Aspergillus tubingensis. Adv. Sci. Eng. Med. 2013, 5, 943–949. [Google Scholar] [CrossRef]
- Burachevskaya, M.; Minkina, T.; Mandzhieva, S.; Bauer, T.; Nevidomskaya, D.; Shuvaeva, V.; Sushkova, S.; Kizilkaya, R.; Gülser, C.; Rajput, V. Transformation of copper oxide and copper oxide nanoparticles in the soil and their accumulation by Hordeum sativum. Environ. Geochem. Health 2021, 43, 1655–1672. [Google Scholar] [CrossRef]
- Gao, F.; Liu, C.; Qu, C.; Zheng, L.; Yang, F.; Su, M.; Hong, F. Was improvement of spinach growth by nano-TiO2 treatment related to the changes of rubisco activase? BioMetals 2008, 21, 211–217. [Google Scholar] [CrossRef]
- Tripathy, B.C.; Oelmüller, R. Reactive oxygen species generation and signaling in plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, L.L.A.; Zhang, H.; Huang, M.; Wu, H.; Xing, B.; Wang, Z.; Ji, R. Nano-biotechonlgy in agriculture: Use of nanomaterials to promote plant growth and stress tolerance. J. Agric. Food Chem. 2020, 68, 1935–1947. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Du, W.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L.; White, J.C.; Keller, A.A.; Guo, H.; Ji, R.; Zhao, L. Metabolomics Reveals How Cucumber (Cucumis sativus) Reprograms Metabolites to Cope with Silver Ions and Silver Nanoparticle-Induced Oxidative Stress. Environ. Sci. Technol. 2018, 52, 8016–8026. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Tomar, R.S.; Kumar, M. Exploring nanobiotechnology to mitigate abiotic stress in crop plants. J. Pharma Sci. Res. 2016, 8, 974. [Google Scholar]
- Wakeel, A.; Xu, M.; Gan, Y. Chromium-Induced Reactive Oxygen Species Accumulation by Altering the Enzymatic Antioxidant System and Associated Cytotoxic, Genotoxic, Ultrastructural, and Photosynthetic Changes in Plants. Int. J. Mol. Sci. 2020, 21, 728. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ma, X.; Zhang, W.; Pei, H.; Chen, Y. The impact of cerium oxide nanoparticles on tomato (Solanum lycopersicum L.) and its implications for food safety. Metallomics 2012, 4, 1105–1112. [Google Scholar] [CrossRef]
- Van Nhan, L.; Ma, C.; Rui, Y.; Liu, S.; Li, X.; Xing, B.; Liu, L. Phytotoxic Mechanism of Nanoparticles: Destruction of Chloroplasts and Vascular Bundles and Alteration of Nutrient Absorption. Sci. Rep. 2015, 5, 11618. [Google Scholar] [CrossRef] [Green Version]
- Pulido-Reyes, G.; Palomares, I.M.R.; Das, S.; Sakthivel, T.S.; Leganes, F.; Rosal, R.; Seal, S.; Fernández-Piñas, F. Untangling the biological effects of cerium oxide nanoparticles: The role of surface valence states. Sci. Rep. 2015, 5, 15613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.N.; AlSolami, M.A.; Basahi, R.A.; Siddiqui, M.; Al-Huqail, A.A.; Abbas, Z.K.; Siddiqui, Z.; Ali, H.M.; Khan, F. Nitric oxide is involved in nano-titanium dioxide-induced activation of antioxidant defense system and accumulation of osmolytes under water-deficit stress in Viciafaba L. Ecotoxicol. Environ. Saf. 2020, 190, 110152. [Google Scholar] [CrossRef]
- Ahmad, B.; Shabbir, A.; Jaleel, H.; Khan, M.M.A.; Sadiq, Y. Efficacy of titanium dioxide nanoparticles in modulating photosynthesis, peltate glandular trichomes and essential oil production and quality in Mentha piperita L. Curr. Plant Biol. 2018, 13, 6–15. [Google Scholar] [CrossRef]
- Yin, J.-J.; Liu, J.; Ehrenshaft, M.; Roberts, J.E.; Fu, P.P.; Mason, R.P.; Zhao, B. Phototoxicity of nano titanium dioxides in HaCaT keratinocytes—Generation of reactive oxygen species and cell damage. Toxicol. Appl. Pharmacol. 2012, 263, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.P.; Xia, Q.; Hwang, H.-M.; Ray, P.C.; Yu, H. Mechanisms of nanotoxicity: Generation of reactive oxygen species. J. Food Drug Anal. 2014, 22, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Fedorenko, A.G.; Minkina, T.M.; Chernikova, N.P.; Fedorenko, G.M.; Mandzhieva, S.S.; Rajput, V.D.; Burachevskaya, M.V.; Chaplygin, V.A.; Bauer, T.V.; Sushkova, S.N.; et al. The toxic effect of CuO of different dispersion degrees on the structure and ultrastructure of spring barley cells (Hordeum sativum distichum). Environ. Geochem. Health 2021, 43, 1673–1687. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Madler, L. Toxic potential of materials at the nano level. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Hondroulis, E.; Liu, W.; Li, C.-Z. Biosensing Approaches for Rapid Genotoxicity and Cytotoxicity Assays upon Nanomaterial Exposure. Small 2013, 9, 1821–1830. [Google Scholar] [CrossRef]
- Vara, D.; Pula, G. Reactive oxygen species: Physiological roles in the regulation of vascular cells. Curr. Mol. Med. 2014, 14, 1103–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, T.; Kovochich, M.; Liong, M.; Mädler, L.; Gilbert, B.; Shi, H.; Yeh, J.I.; Zink, J.I.; Nel, A.E. Comparison of the Mechanism of Toxicity of Zinc Oxide and Cerium Oxide Nanoparticles Based on Dissolution and Oxidative Stress Properties. ACS Nano 2008, 2, 2121–2134. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Saadony, M.T.; Saad, A.M.; Najjar, A.A.; Alzahrani, S.O.; Alkhatib, F.M.; Selem, E.; Desoky, S.M.; Fouda, S.S.; El-Tahan, A.M.; Hassan, M.A.A. The use of biological selenium nanoparticles in controlling Triticum aestivum L. crown root and rot diseases induced by Fusarium species and improve yield under drought and heat stress. Saudi J. Biol. Sci. 2021, 28, 4461–4471. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicumesculentum seeds Mill.). Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaine, S.J.; Alvarez, P.J.J.; Batley, G.E.; Fernandes, T.F.; Handy, R.D.; Lyon, D.Y.; Mahendra, S.; McLaughlin, M.J.; Lead, J.R. Nanomaterials in the environment: Behavior, fate, bioavailability, and effects. Environ. Toxicol. Chem. 2008, 27, 1825–1851. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Shen, C.; Ji, Q.; An, H.; Wang, J.; Liu, Q.; Zhang, Z. Food storage material silver nanoparticles interfere with DNA replication fidelity and bind with DNA. Nanotechnology 2009, 20, 085102. [Google Scholar] [CrossRef]
- Karimi, E.; Fard, E.M. Nanomaterial effects on soil microorganisms. In Nanoscience and Plant–Soil Systems. Soil Biology 48; Ghorbanpour, M., Manika, K., Varma, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 137–200. [Google Scholar]
- Yang, Y.; Wang, J.; Xiu, Z.; Alvarez, P.J.J. Impacts of silver nanoparticles on cellular and transcriptional activity of nitrogen-cycling bacteria. Environ. Toxicol. Chem. 2013, 32, 1488–1494. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, M.; Schirmer, K. Interference of silver nanoparticles with essential metal homeostasis in a novel enterohepatic fish in vitro system. Environ. Sci. Nano 2019, 6, 1777–1790. [Google Scholar] [CrossRef]
- Weber, N.C.; Koron, D.; Jakopič, J.; Veberič, R.; Hudina, M.; Česnik, H.B. Influence of Nitrogen, Calcium and Nano-Fertilizer on Strawberry (Fragaria × ananassa Duch.) Fruit Inner and Outer Quality. Agronomy 2021, 11, 997. [Google Scholar] [CrossRef]
- Mirzajani, F.; Askari, H.; Hamzelou, S.; Schober, Y.; Römpp, A.; Ghassempour, A.; Spengler, B. Proteomics study of silver nanoparticles toxicity on Bacillus thuringiensis. Ecotoxicol. Environ. Saf. 2014, 100, 122–130. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-Fertilization as an Emerging Fertilization Technique: Why Can Modern Agriculture Benefit from Its Use? Plants 2021, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.A.; Hassan, M.M.; Ibrahim, E.A.; Abou-Baker, N.H.; Shaaban, E.A. Carbon nanotubes impact on date palm in vitro cultures. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 127, 525–534. [Google Scholar] [CrossRef]
- El-Kereti, M.A.; El-Feky, S.A.; Khater, M.S.; Osman, Y.A.; El-Sherbini, E.S.A. ZnOnanofertilizer and He Ne laser irradiation for promoting growth and yield of sweet basil plant. Recent Pat. Food Nutr. Agric. 2013, 5, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Rico, C.; Barrios, A.C.; Tan, W.; Rubenecia, R.; Lee, S.C.; Varela-Ramirez, A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Physiological and biochemical response of soil-grown barley (Hordeum vulgare L.) to cerium oxide nanoparticles. Environ. Sci. Pollut. Res. 2015, 22, 10551–10558. [Google Scholar] [CrossRef]
- Rico, C.; Lee, S.C.; Rubenecia, R.; Mukherjee, A.; Hong, J.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Cerium Oxide Nanoparticles Impact Yield and Modify Nutritional Parameters in Wheat (Triticum aestivum L.). J. Agric. Food Chem. 2014, 62, 9669–9675. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Gardea-Torresdey, J.L.; Ji, R.; Yin, Y.; Zhu, J.; Peralta-Videa, J.R.; Guo, H. Physiological and Biochemical Changes Imposed by CeO2 Nanoparticles on Wheat: A Life Cycle Field Study. Environ. Sci. Technol. 2015, 49, 11884–11893. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Peralta-Videa, J.R.; Rico, C.M.; Hernandez-Viezcas, J.A.; Sun, Y.; Niu, G.; Servin, A.; Nunez, J.E.; Duarte-Gardea, M.; Gardea-Torresdey, J.L. CeO2 and ZnO Nanoparticles Change the Nutritional Qualities of Cucumber (Cucumis sativus). J. Agric. Food Chem. 2014, 62, 2752–2759. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.I.; Rico, C.M.; Hernandez-Viezcas, J.A.; Nunez, J.E.; Barrios, A.C.; Tafoya, A.; Flores-Marges, J.P.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Toxicity Assessment of Cerium Oxide Nanoparticles in Cilantro (Coriandrum sativum L.) Plants Grown in Organic Soil. J. Agric. Food Chem. 2013, 61, 6224–6230. [Google Scholar] [CrossRef] [PubMed]
- López-Vargas, E.R.; Ortega-Ortíz, H.; Cadenas-Pliego, G.; Romenus, K.D.A.; de la Fuente, M.C.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Foliar Application of Copper Nanoparticles Increases the Fruit Quality and the Content of Bioactive Compounds in Tomatoes. Appl. Sci. 2018, 8, 1020. [Google Scholar] [CrossRef] [Green Version]
- Hernández, H.H.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Hernández-Fuentes, A.D.; Juárez-Maldonado, A. Cu Nanoparticles in chitosan-PVA hydrogels as promoters of growth, productivity and fruit quality in tomato. Emir. J. Food Agric. 2017, 29, 573–580. [Google Scholar]
- Zhao, L.; Huang, Y.; Hu, J.; Zhou, H.; Adeleye, A.S.; Keller, A.A. 1H NMR and GC-MS Based Metabolomics Reveal Defense and Detoxification Mechanism of Cucumber Plant under Nano-Cu Stress. Environ. Sci. Technol. 2016, 50, 2000–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Hu, J.; Huang, Y.; Wang, H.; Adeleye, A.; Ortiz, C.; Keller, A.A. 1H NMR and GC–MS based metabolomics reveal nano-Cu altered cucumber (Cucumis sativus) fruit nutritional supply. Plant Physiol. Biochem. 2017, 110, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, H.; González-Morales, S.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Juárez-Maldonado, A. Effects of Chitosan–PVA and Cu Nanoparticles on the Growth and Antioxidant Capacity of Tomato under Saline Stress. Molecules 2018, 23, 178. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, B.; Shahid, M.; Khan, M.S.; Musarrat, J. Chromosomal aberrations, cell suppression and oxidative stress generation induced by metal oxide nanoparticles in onion (Allium cepa) bulb. Metallomics 2018, 10, 1315–1327. [Google Scholar] [CrossRef]
- Apodaca, S.A.; Tan, W.; Dominguez, O.E.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Physiological and biochemical effects of nanoparticulate copper, bulk copper, copper chloride, and kinetin in kidney bean (Phaseolus vulgaris) plants. Sci. Total Environ. 2017, 599, 2085–2094. [Google Scholar] [CrossRef]
- Juarez-Maldonado, A.; Ortega-Ortíz, H.; Pérez-Labrada, F.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Cu Nanoparticles absorbed on chitosan hydrogels positively alter morphological, production, and quality characteristics of tomato. J. Appl. Bot. Food Qual. 2016, 89, 183–189. [Google Scholar] [CrossRef]
- Hoang, S.A.; Nguyen, L.Q.; Nguyen, N.H.; Tran, C.Q.; Nguyen, D.V.; Le, N.T.; Ha, C.V.; Vu, Q.N.; Phan, C.M. Metal nanoparticles as effective promotors for Maize production. Sci. Rep. 2019, 9, 13925. [Google Scholar] [CrossRef] [Green Version]
- Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Silica Nanoparticles for Increased Silica Availability in Maize (Zea mays L) Seeds Under Hydroponic Conditions. Curr. Nanosci. 2012, 8, 902–908. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, N.; Liang, X.; Bai, X.; Zheng, L.; Zhao, J.; Li, Y.-F.; Zhang, Z.; Gao, Y. Silica nanoparticles alleviate mercury toxicity via immobilization and inactivation of Hg(ii) in soybean (Glycine max). Environ. Sci. Nano 2020, 7, 1807–1817. [Google Scholar] [CrossRef]
- Ashour, H.A.; Abdel Wahab, M.; Mahmoud, A.W.M. Response of Jatropha integerrima Plants Irrigated with Different Levels of Saline Water to Nano Silicon and Gypsum. J. Agric. Stud. 2017, 5, 136–160. [Google Scholar] [CrossRef] [Green Version]
- Almutairi, Z.M. Effect of nano-silicon application on the expression of salt tolerance genes in germinating tomato (Solanum lycopersicum L.) seedlings under salt stress. Plant Omics 2016, 9, 106. [Google Scholar]
- Ashkavand, P.; Zarafshar, M.; Tabari, M.; Mirzaie, J.; Nikpour, A.; Bordbar, S.K.; Struve, D.; Striker, G.G. Application of SiO2 nanoparticles as pretreatment alleviates the impact of drought on the physiological performance of Prunus mahaleb (Rosaceae). Bol. Soc. Argent. Botánica 2018, 53, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Qados, A.M.S.A.; Moftah, A.E. Influence of Silicon and Nano-Silicon on Germination, Growth and Yield of Faba Bean (Viciafaba L.) Under Salt Stress Conditions. Am. J. Exp. Agric. 2015, 5, 509. [Google Scholar] [CrossRef]
- Yassen, A.E.; Abdallah, M.; Gaballah, M.; Zaghloul, S. Role of silicon dioxide nano fertilizer in mitigating salt stress on growth, yield and chemical composition of cucumber (Cucumis sativus L.). Int. J. Agric. Res. 2017, 12, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, R.; Esna-Ashari, M. The effect of micro-and nanoparticles of silicon on concentration of macro-and microelements and silicon content of strawberry plant in soilless culture conditions. J. Sci. Tech. Greenh. Cult. 2017, 8, 57–71. [Google Scholar]
- Elsheery, N.I.; Sunoj, V.; Wen, Y.; Zhu, J.; Muralidharan, G.; Cao, K. Foliar application of nanoparticles mitigates the chilling effect on photosynthesis and photoprotection in sugarcane. Plant Physiol. Biochem. 2020, 149, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanpour, M.; Mohammadi, H.; Kariman, K. Nanosilicon-based recovery of barley (Hordeum vulgare) plants subjected to drought stress. Environ. Sci. Nano 2020, 7, 443–461. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Attia, E.A.; Elhawat, N. Combined foliar and soil application of silica nanoparticles enhances the growth, flowering period and flower characteristics of marigold (Tageteserecta L.). Sci. Hortic. 2021, 282, 110015. [Google Scholar] [CrossRef]
- Schaller, J.; Cramer, A.; Carminati, A.; Zarebanadkouki, M. Biogenic amorphous silica as main driver for plant available water in soils. Sci. Rep. 2020, 10, 2424. [Google Scholar] [CrossRef] [Green Version]
- Sheykhbaglou, R.; Sedghi, M.; Shishevan, M.T.; Sharifi, R.S. Effects of Nano-Iron Oxide Particles on Agronomic Traits of Soybean. Not. Sci. Biol. 2010, 2, 112–113. [Google Scholar] [CrossRef] [Green Version]
- Rui, M.; Ma, C.; Hao, J.; Guo, Y.; Rui, X.; Tang, X.; Zhu, S. Iron oxide nanoparticles as a potential iron fertilizer for peanut (Arachishypogaea). Front. Plant Sci. 2016, 7, 815. [Google Scholar] [CrossRef] [Green Version]
- Shankramma, K.; Yallappa, S.; Shivanna, M.B.; Manjanna, J. Fe2O3 magnetic nanoparticles to enhance S. lycopersicum (tomato) plant growth and their biomineralization. Appl. Nanosci. 2016, 6, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Servin, A.D.; Morales, M.I.; Castillo-Michel, H.; Hernandez-Viezcas, J.A.; Munoz, B.; Zhao, L.; Nunez, J.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Synchrotron Verification of TiO2 Accumulation in Cucumber Fruit: A Possible Pathway of TiO2 Nanoparticle Transfer from Soil into the Food Chain. Environ. Sci. Technol. 2013, 47, 11592–11598. [Google Scholar] [CrossRef] [PubMed]
- Zahra, Z.; Waseem, N.; Zahra, R.; Lee, H.; Badshah, M.A.; Mehmood, A.; Choi, H.-K.; Arshad, M. Growth and Metabolic Responses of Rice (Oryza sativa L.) Cultivated in Phosphorus-Deficient Soil Amended with TiO2 Nanoparticles. J. Agric. Food Chem. 2017, 65, 5598–5606. [Google Scholar] [CrossRef]
- Raliya, R.; Nair, R.; Chavalmane, S.; Wang, W.-N.; Biswas, P. Mechanistic evaluation of translocation and physiological impact of titanium dioxide and zinc oxide nanoparticles on the tomato (Solanum lycopersicum L.) plant. Metallomics 2015, 7, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Sharma, N.C.; Fleischmann, P.; Burbage, J.; Venkatachalam, P.; Sahi, S.V. Nanotitania Exposure Causes Alterations in Physiological, Nutritional and Stress Responses in Tomato (Solanum lycopersicum). Front. Plant Sci. 2017, 8, 633. [Google Scholar] [CrossRef] [Green Version]
- Hong, F.; Yang, P.; Gao, F.-Q.; Liu, C.; Zheng, L.; Yang, F.; Zhou, J. Effect of nano-anatase TiO2 on spectral characterization of photosystem particles from spinach. Chem. Res. China Univ. 2005, 21, 196–200. [Google Scholar]
- Qi, M.; Liu, Y.; Li, T. Nano-TiO2 improve the photosynthesis of tomato leaves under mild heat stress. Biol. Trace Elem. Res. 2013, 156, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Dağhan, H.; Gülmezoğlu, N.; Köleli, N.; Karakaya, B. Impact of titanium dioxide nanoparticles (TiO2-NPs) on growth and mineral nutrient uptake of wheat (Triticum vulgare L.). Biotech Stud. 2020, 29, 69–76. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.; Borthakur, A.; Srivastava, P.; Srivastava, N.; Tiwary, D.; Mishra, P.K. Effect of nanoscale TiO2-activated carbon composite on Solanum lycopersicum (L.) and Vigna radiata (L.) seeds germination. Energ. Ecol. Environ. 2016, 1, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Konate, A.; Wang, Y.; He, X.; Adeel, M.; Zhang, P.; Ma, Y.; Ding, Y.; Zhang, J.; Yang, J.; Kizito, S.; et al. Comparative effects of nano and bulk-Fe3O4 on the growth of cucumber (Cucumis sativus). Ecotoxicol. Environ. Saf. 2018, 165, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Tombuloglu, H.; Slimani, Y.; Tombuloglu, G.; Almessiere, M.; Baykal, A. Uptake and translocation of magnetite (Fe3O4) nanoparticles and its impact on photosynthetic genes in barley (Hordeum vulgare L.). Chemosphere 2019, 226, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, Z.M. Influence of Silver Nano-particles on the Salt Resistance of Tomato (Solanum lycopersicum) during Germination. Int. J. Agric. Biol. 2016, 18, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Younes, N.A.; Nassef, D.M. Effect of silver nanoparticles on salt tolerancy of tomato transplants (Solanum lycopersicom L. Mill.). Assiut J. Agric. Sci. 2015, 46, 76–85. [Google Scholar]
- Ma, C.; Liu, H.; Chen, G.; Zhao, Q.; Guo, H.; Minocha, R.; Long, S.; Tang, Y.; Saad, E.M.; de la Torre Roche, R.; et al. Dual roles of glutathione in silver nanoparticle detoxification and enhancement of nitrogen assimilation in soybean (Glycine max (L.) Merrill). Environ. Sci. Nano 2020, 7, 1954–1966. [Google Scholar] [CrossRef]
- Raddy, R.; Salimath, M.; Geetha, K.; Shankar, A. ZnO Nanoparticle Improves Maize Growth, Yield and Seed Zinc under High Soil pH Condition. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1593–1601. [Google Scholar] [CrossRef]
- Bahri, S.; Bhatia, S.S.; Moitra, S.; Sharma, N.; Bhatt, R.; Borthakur, N.S.; Agarwal, R.; Jain, D. Influence of silver nanoparticles on seedlings of Vigna radiata L. R. Wilczek. DU J. Undergrad. Res. Innov. 2016, 2, 142–148. [Google Scholar]
- Alharby, H.F.; Metwali, E.M.; Fuller, M.; Aldhebiani, A.Y. Impact of application of zinc oxide nanoparticles on callus induction, plant regeneration, element content and antioxidant enzyme activity in tomato (Solanum lycopersicum Mill.) under salt stress. Arch. Biol. Sci. 2016, 68, 723–735. [Google Scholar] [CrossRef] [Green Version]
- Prasad, T.N.V.K.V.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Reddy, K.R.; Sreeprasad, T.S.; Sajanlal, P.R.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Tavallali, V.; Rowshan, V.; Bahmanzadegan, A. Variations in sweet basil in response to Green synthesized Zinc-Amino nano complexes. J. Clean. Prod. 2018, 196, 452–459. [Google Scholar] [CrossRef]
- Rajiv, P.; Vanathi, P. Effect of Parthenium based vermicompost and zinc oxide nanoparticles on growth and yield of Arachis hypogaea L. in zinc deficient soil. Biocatal. Agric. Biotechnol. 2018, 13, 251–257. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; White, J.C.; Elmer, W.H.; Gardea-Torresdey, J. Nanoparticle and Ionic Zn Promote Nutrient Loading of Sorghum Grain under Low NPK Fertilization. J. Agric. Food Chem. 2017, 65, 8552–8559. [Google Scholar] [CrossRef]
- Deshpande, P.; Dapkekar, A.; Oak, M.D.; Paknikar, K.M.; Rajwade, J.M. Zinc complexed chitosan/TPP nanoparticles: A promising micronutrient nanocarrier suited for foliar application. Carbohydr. Polym. 2017, 165, 394–401. [Google Scholar] [CrossRef]
- Behboudi, F.; Tahmasebi Sarvestani, Z.; Kassaee, M.Z.; Modares Sanavi, S.A.M.; Sorooshzadeh, A.; Ahmadi, S.B. Evaluation of Chitosan Nanoparticles Effects on Yield and Yield Components of Barley (Hordeum vulgare L.) under Late Season Drought Stress. J. Water Environ. Nanotechnol. 2018, 3, 22–39. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.; Hasaneen, M.N.A.; Omer, A.M. Nano chitosan-NPK fertilizer enhances the growth and productivity of wheat plants grown in sandy soil. Span. J. Agric. Res. 2016, 14, e0902. [Google Scholar] [CrossRef]
- Prasad, A.; Astete, C.E.; Bodoki, A.E.; Windham, M.; Bodoki, E.; Sabliov, C.M. Zein Nanoparticles Uptake and Translocation in Hydroponically Grown Sugar Cane Plants. J. Agric. Food Chem. 2017, 66, 6544–6551. [Google Scholar] [CrossRef]
- Kumar, V.; Guleria, P.; Kumar, V.; Yadav, S.K. Gold nanoparticle exposure induces growth and yield enhancement in Arabidopsis thaliana. Sci. Total Environ. 2013, 461, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Sharma, P.; Kumar, S.; Nayan, R.; Khanna, P.K.; Zaidi, M.G.H. Gold-nanoparticle induced enhancement in growth and seed yield of Brassica juncea. Plant Growth Regul. 2012, 66, 303–310. [Google Scholar] [CrossRef]
- Lu, L.; Huang, M.; Huang, Y.; Corvini, P.F.-X.; Ji, R.; Zhao, L. Mn3O4 nanozymes boost endogenous antioxidant metabolites in cucumber (Cucumis sativus) plant and enhance resistance to salinity stress. Environ. Sci. Nano 2020, 7, 1692–1703. [Google Scholar] [CrossRef]
- Badran, A.; Savin, I. Effect of Nano-Fertilizer on Seed Germination and First Stages of Bitter Almond Seedlings’ Growth Under Saline Conditions. BioNanoScience 2018, 8, 742–751. [Google Scholar] [CrossRef]
- Laurent, S.; Forge, D.; Port, M.; Roch, A.; Robic, C.; Vander Elst, L.; Muller, R.N. Magnetic Iron Oxide Nanoparticles: Synthesis, Stabilization, Vectorization, Physicochemical Characterizations, and Biological Applications. Chem. Rev. 2008, 108, 2064–2110. [Google Scholar] [CrossRef]
- Li, J.; Hu, J.; Ma, C.; Wang, Y.; Wu, C.; Huang, J.; Xing, B. Uptake, translocation and physiological effects of magnetic iron oxide (γ-Fe2O3) nanoparticles in corn (Zea mays L.). Chemosphere 2016, 159, 326–334. [Google Scholar] [CrossRef]
- Zhu, H.; Han, J.; Xiao, J.Q.; Jin, Y. Uptake, translocation, and accumulation of manufactured iron oxide nanoparticles by pumpkin plants. J. Environ. Monit. 2008, 10, 713–717. [Google Scholar] [CrossRef]
- Li, J.; Chang, P.R.; Huang, J.; Wang, Y.; Yuan, H.; Ren, H. Physiological Effects of Magnetic Iron Oxide Nanoparticles Towards Watermelon. J. Nanosci. Nanotechnol. 2013, 13, 5561–5567. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Cui, X.; He, S.; Dong, G.; Chen, M.; Wang, J.; Lin, X. The role of metal nanoparticles in influencing arbuscular mycorrhizal fungi effects on plant growth. Environ. Sci. Technol. 2013, 47, 9496–9504. [Google Scholar] [CrossRef] [PubMed]
- Alidoust, D.; Isoda, A. Effect of γFe2O3 nanoparticles on photosynthetic characteristic of soybean (Glycine max (L.) Merr.): Foliar spray versus soil amendment. Acta Physiol. Plant. 2013, 35, 3365–3375. [Google Scholar] [CrossRef]
- Alidoust, D.; Isoda, A. Phytotoxicity assessment of γ-Fe2O3 nanoparticles on root elongation and growth of rice plant. Environ. Earth Sci. 2014, 71, 5173–5182. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Keeping, M.; Meyer, J.H. Silicon-augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-Ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon-Mediated Tolerance to Salt Stress. Silicon in Agriculture; Springer: Dordrecht, The Netherlands, 2015; pp. 123–142. [Google Scholar] [CrossRef]
- Babajani, A.; Iranbakhsh, A.; Ardebili, Z.O.; Eslami, B. Differential growth, nutrition, physiology, and gene expression in Melissa officinalis mediated by zinc oxide and elemental selenium nanoparticles. Environ. Sci. Pollut. Res. 2019, 26, 24430–24444. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P. Nutrient Cycling and Nutrient Use Efficiency. Am. Nat. 1982, 119, 553–572. [Google Scholar] [CrossRef]
- Neu, S.; Schaller, J.; Dudel, E.G. Silicon availability modifies nutrient use efficiency and content, C:N:P stoichiometry, and productivity of winter wheat (Triticum aestivum L.). Sci. Rep. 2017, 7, 40829. [Google Scholar] [CrossRef] [PubMed]
- Carey, J.C.; Fulweiler, R.W. Human appropriation of biogenic silicon—The increasing role of agriculture. Funct. Ecol. 2016, 30, 1331–1339. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, S.C. The uptake and transport of silicon by perennial ryegrass and wheat. Plant Soil 1987, 97, 429–437. [Google Scholar] [CrossRef]
- Schaller, J.; Faucherre, S.; Joss, H.; Obst, M.; Goeckede, M.; Planer-Friedrich, B.; Peiffer, S.; Gilfedder, B.; Elberling, B. Silicon increases the phosphorus availability of Arctic soils. Sci. Rep. 2019, 9, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, J.N.; Tiwari, R.N.; Kim, K.S. Zero-dimensional, one-dimensional, two-dimensional and three-dimensional nanostructured materials for advanced electrochemical energy devices. Prog. Mater. Sci. 2012, 57, 724–803. [Google Scholar] [CrossRef]
- Leon-Silva, S.; Arrieta-Cortes, R.; Fernández-Luqueño, F.; López-Valdez, F. Design and production of nanofertilizers. In Agricultural Nanobiotechnology; Springer Science and Business Media LLC: Cham, Switzerland, 2018; pp. 17–31. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verma, K.K.; Song, X.-P.; Joshi, A.; Tian, D.-D.; Rajput, V.D.; Singh, M.; Arora, J.; Minkina, T.; Li, Y.-R. Recent Trends in Nano-Fertilizers for Sustainable Agriculture under Climate Change for Global Food Security. Nanomaterials 2022, 12, 173. https://doi.org/10.3390/nano12010173
Verma KK, Song X-P, Joshi A, Tian D-D, Rajput VD, Singh M, Arora J, Minkina T, Li Y-R. Recent Trends in Nano-Fertilizers for Sustainable Agriculture under Climate Change for Global Food Security. Nanomaterials. 2022; 12(1):173. https://doi.org/10.3390/nano12010173
Chicago/Turabian StyleVerma, Krishan K., Xiu-Peng Song, Abhishek Joshi, Dan-Dan Tian, Vishnu D. Rajput, Munna Singh, Jaya Arora, Tatiana Minkina, and Yang-Rui Li. 2022. "Recent Trends in Nano-Fertilizers for Sustainable Agriculture under Climate Change for Global Food Security" Nanomaterials 12, no. 1: 173. https://doi.org/10.3390/nano12010173