
New Finding of Eggs and Leptocephalus for Muraenesox cinereus off Jeju Island, Korea and Its Relation to Environmental Variables

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Morphological Identification and Eggs Type Divide
2.3. Genomic DNA Extraction, Polymerase Chain Reaction (PCR), and Sequencing
3. Results
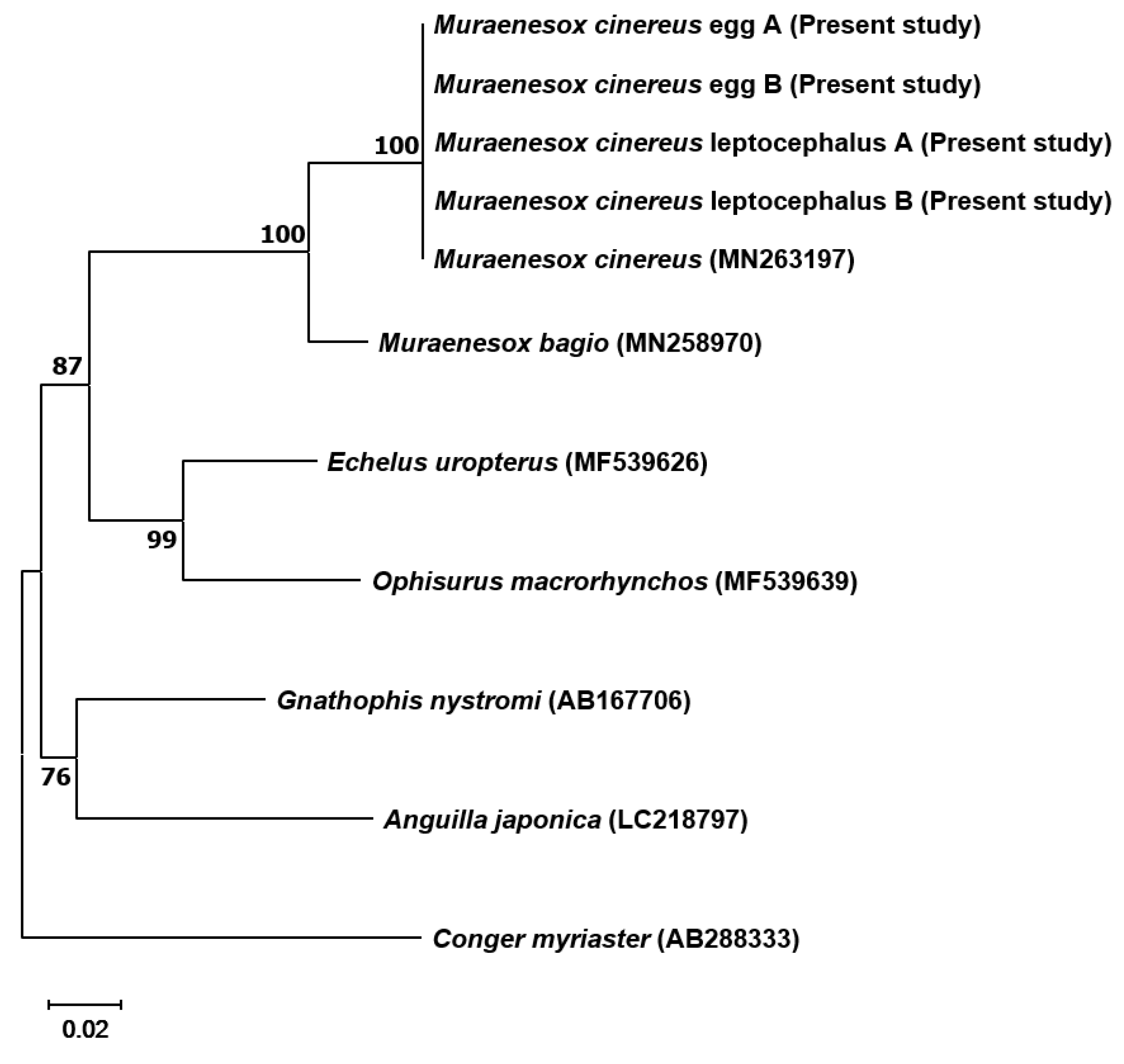
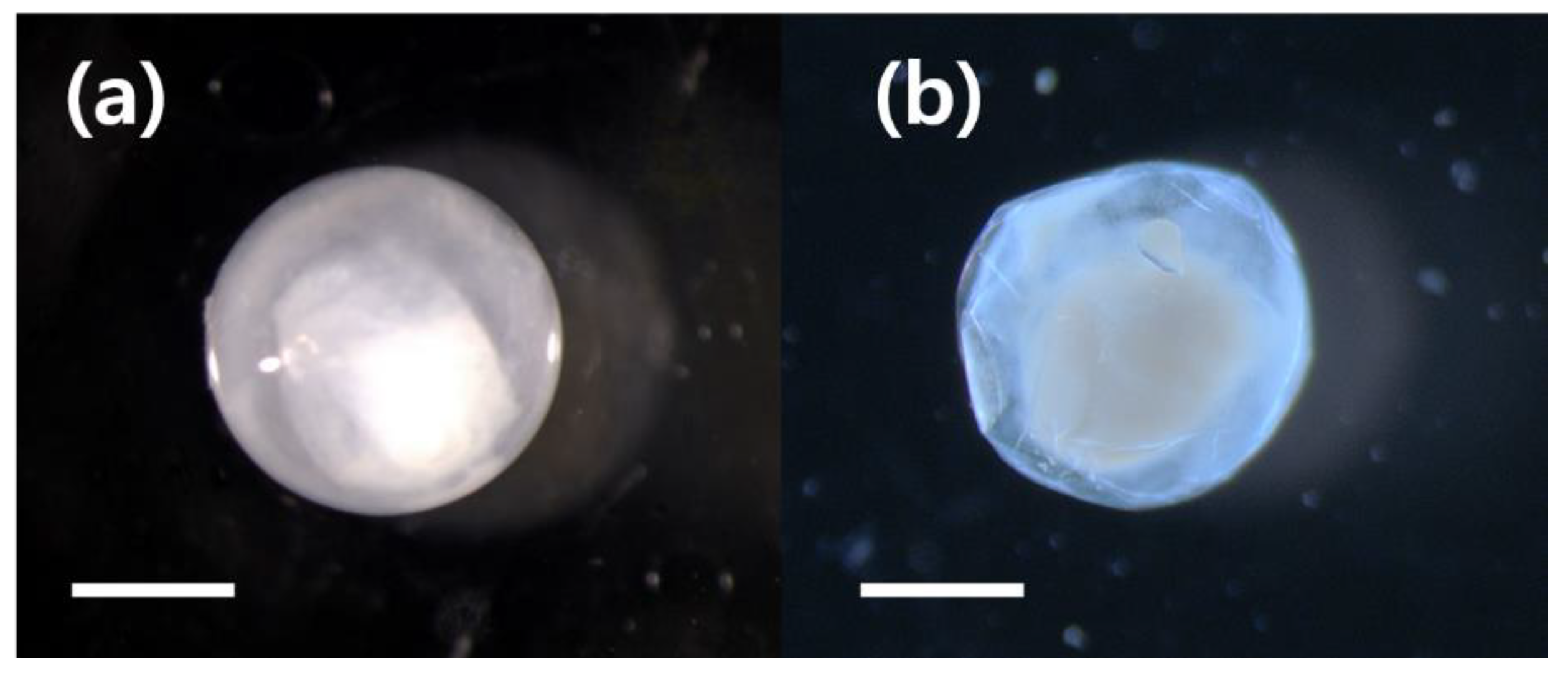
3.1. Molecular Identification Morphological Characteristics of M. cinereus Eggs and Preletocephalus
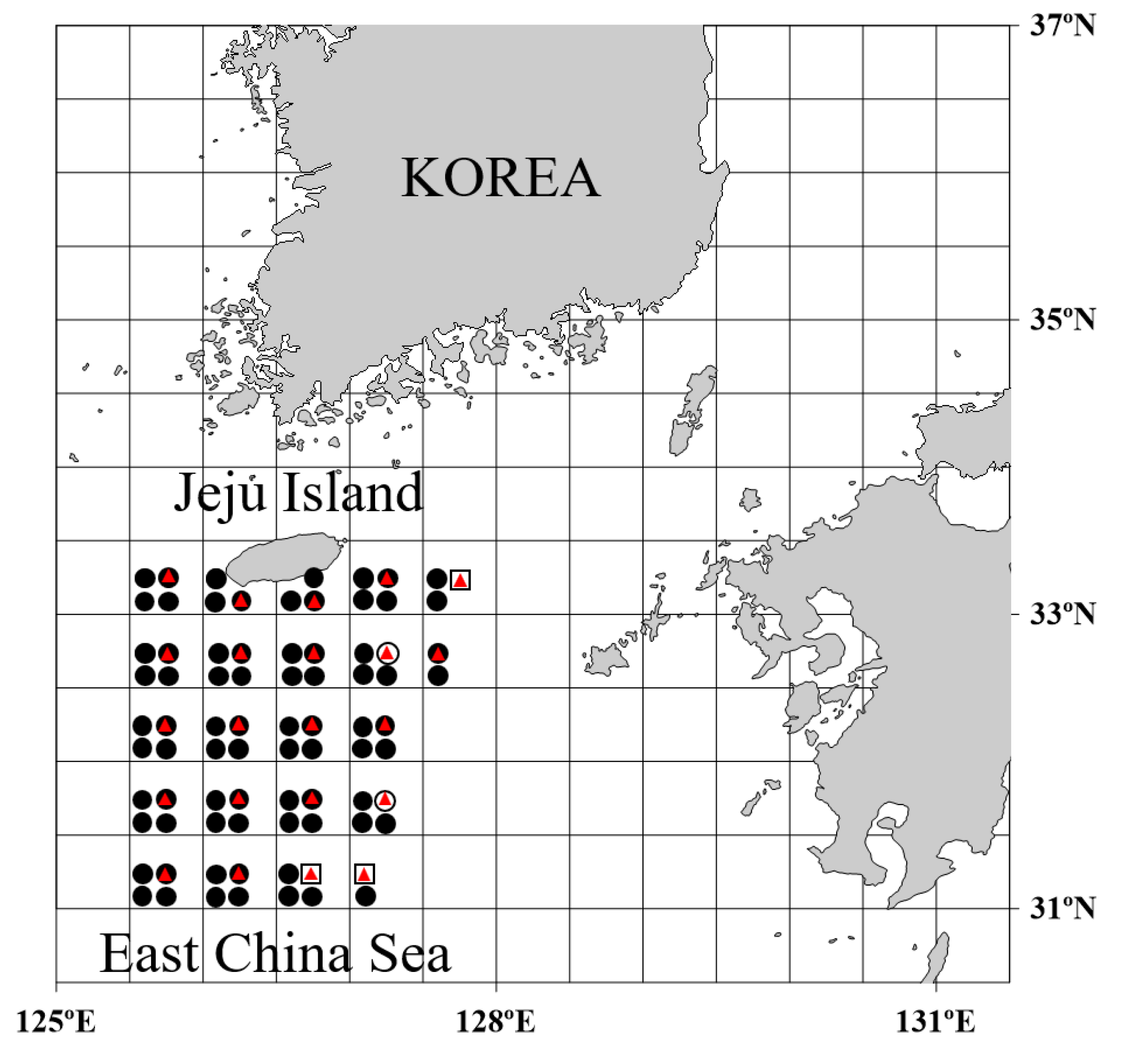
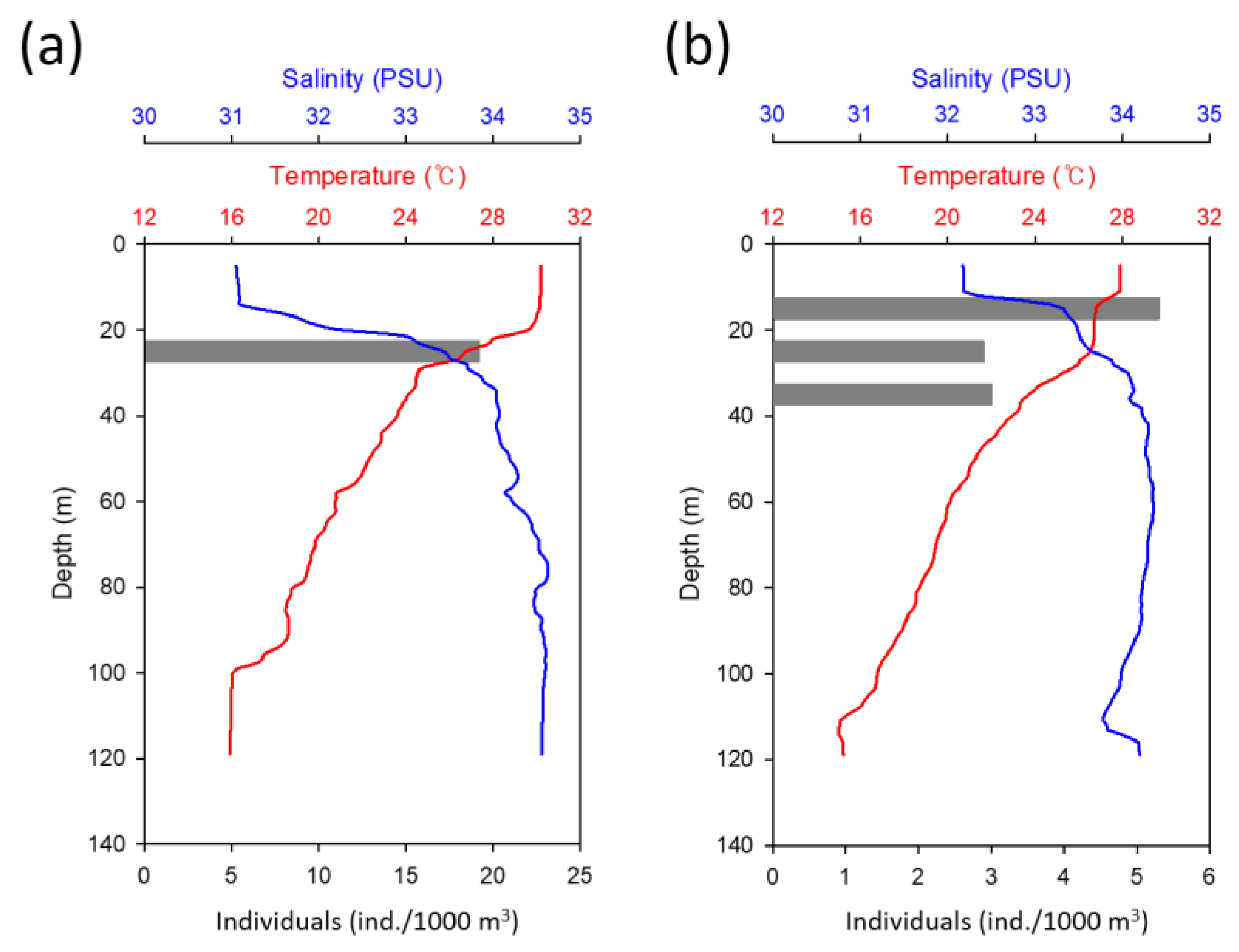
3.2. Spawning Area Characteristics of Muraenesox cinereus and Collection of Preleptocephalus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Russel, B.C.; Houston, W. Offshore fishes of the Arafura Sea. Beagle 1989, 6, 69–84. [Google Scholar] [CrossRef]
- Eschmeyer’s Catalog of Fishes. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 13 December 2022).
- An, Y.S.; Park, J.M.; Kim, H.J.; Haeck, G.W. Feeding habitat of daggertooth pike conger Muraenesox cinereus in the coastal water off Goseong, Korea. Korean J. Fish. Aquat. Sci. 2012, 45, 76–81. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. Fishbase. Available online: http://www.fishbase.org (accessed on 2 January 2023).
- NIFS (National Institute of Fisheries Science). Ecology and Fishing Ground of Fisheries Resources in the Korean Waters; Ye-Moon-Publishing: Busan, Korea, 2021; pp. 19–30. [Google Scholar]
- Ji, H.S.; Kim, J.K.; Oh, T.Y.; Choi, K.H.; Choi, J.H.; Seo, Y.I.; Lee, D.W. Larval distribution pattern of Muraenesox cinereus (Anguilliformes: Muraenesocidae) leptocephali in waters adjacent to Korea. Ocean Sci. J. 2015, 50, 537–545. [Google Scholar] [CrossRef]
- Kim, M.J.; Kang, Y.J.; Park, C.S. A study on the stock management of sharp-toothed eel, Muraenesox cinereus (FORSKAL) in Korean waters II. Age and growth. J. Korean Soc. Fish. Res. 1998, 1, 11–17. [Google Scholar]
- Watari, S.; Murata, M.; Hinoshita, Y.; Mishiro, K.; Oda, S.; Ishitani, M. Re-examination of age and growth of daggertooth pike conger Muraenesox cinereus in the western Seto Inland Sea. Jpn. Fish. Sci. 2013, 79, 367–373. [Google Scholar] [CrossRef]
- Kor, E.H.; Kwon, D.H.; Kim, Y.H. Age and growth of daggertooth pike conger Muraenesox cinereus in the South Sea of Korea. Korean J. Fish. Aquat. Sci. 2019, 52, 650–655. [Google Scholar] [CrossRef]
- Cha, H.K.; Seo, Y.I.; Oh, T.Y.; Kim, H.Y.; Lee, S.G.; Choi, M.S. Reproductive ecology of the sharp toothed eel in the southern Korean waters. Korean Soc. Fish. Technol. 2012, 48, 217–226. [Google Scholar] [CrossRef]
- Tang, Y.; Wu, C. Feeding habit of conger pike Muraenesox cinereus in Zhejiang coastal waters. J. Zhejiang Ocean Univ. 1992, 2, 98–101. [Google Scholar]
- Okazaki, T.; Ueta, Y.; Hamano, T. Distribution and migration of daggertooth pike-conger Muraenesox cinereus in the eastern Seto Inland Sea, Japan estimated by mark and recapture experiments. Nippon. Suisan Gakkaishi 2012, 78, 913–921. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Mototani, T.; Murayama, F.; Sakamoto, T. Basic reproductive biology of daggertooth pike conger, Muraenesox cinereus: A possible model for oogenesis in Anguilliformes. Zool. Lett. 2015, 1, 25. [Google Scholar] [CrossRef] [Green Version]
- Kor, E.H.; Kwon, D.H.; Jang, C.S. Basic reproductive biology of Muraenesox cinereus in Korean waters. J. Korean Soc. Fish. Technol. 2018, 54, 353–359. [Google Scholar]
- Zhou, Y.D.; Xu, H.X. Biomass estimates of Muraenesox cinereus are calculated by LCA in the East China Sea. J. Zhejiang Ocean Univ. 2007, 4, 399–403. [Google Scholar]
- Watari, S.; Murata, M.; Bada, T.; Oda, S.; Ishitani, M.; Mishiro, K.; Uchida, Y. Fisheries resource management of the daggertooth pike conger, Muraenesox cinereus, using existing limited datasets in western Seo Inland Sea, Japan. Fish. Manag. Ecol. 2014, 21, 470–479. [Google Scholar] [CrossRef]
- Yamada, U.; Tokimura, M.; Horikawa, H.; Nakabo, T. Fisheries and Fisheries of the East China and Yellow Seas; Tokai University Press: Tokyo, Japan, 2007; pp. 165–170. [Google Scholar]
- Tsukamoto, K. Discovery of the spawning area for Japanese eel. Nature 1992, 356, 789–791. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Chow, S.; Otake, T.; Kurogi, H.; Mochioka, N.; Miller, M.J.; Aoyama, J.; Kimura, S.; Watanabe, S.; Yoshinaga, T.; et al. Oceanic spawning ecology of freshwater eels in the western North Pacific. Nat. Commun. 2011, 2, 179. [Google Scholar] [CrossRef] [Green Version]
- Kurogi, H.; Mochioka, N.; Okazaki, M.; Takahashi, M.; Miller, M.J.; Tsukamoto, K.; Ambe, D.; Katayama, S.; Chow, S. Discovery of a spawning area of the common Japanese conger Conger myriaster along the Kyushu-Palau ridge in the western north Pacific. Fish. Sci. 2012, 78, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Umezawa, A.; Otake, T.; Hirokawa, J.; Tsukamoto, K.; Okiyama, M. Development of the eggs and larvae of the pike eel, Muraenesox cinereus. Jpn. J. Ichthyol. 1991, 38, 35–39. [Google Scholar] [CrossRef]
- Lin, H.Y.; Chiu, M.Y.; Shih, Y.M.; Chen, I.S.; Lee, M.A.; Shao, K.T. Species composition and assemblages of ichthyoplankton during summer in the East China Sea. Cont. Shelf Res. 2016, 126, 64–78. [Google Scholar] [CrossRef]
- NIFS (National Institute of Fisheries Science). Fisheries Resources Investigation Techniques; National Institute of Fisheries Science: Busan, Korea, 2021; p. 170. [Google Scholar]
- Ji, H.S.; Yoo, H.J.; Kim, J.K.; Kim, D.N.; Kim, S.T.; Kim, J.N.; Kim, H.J.; Moon, S.Y.; Shin, D.H.; Oh, T.Y.; et al. Fish eggs, Larvae and Juveniles of Korea; Hangeul Graphics: Busan, Korea, 2020; p. 442. [Google Scholar]
- Lira, N.L.; Tonello, S.; Lui, R.L.; Traldi, J.B.; Brandão, H.; Oliveira, C.; Blanco, D.R. Identifying fish eggs and larvae: From classic methodologies to DNA metabarcoding. Mol. Biol. Rep. 2023, 50, 1713–1726. [Google Scholar] [CrossRef]
- Jang, S.H.; Kim, J.K. First report on the occurrence of eggs of the small yellow croaker Larimichthys polyactis from Chilsan-do Island, Jeollanam-do, Korea. Korean J. Fish. Aquat. Sci. 2020, 53, 650–655. [Google Scholar] [CrossRef]
- Baek, J.I.; Ji, H.S.; Yu, H.J.; Hwang, K.S.; Kim, D.N. Distribution of eggs and larvae in coastal waters of Korea. Korean J. Fish. Aquat. Sci. 2021, 54, 467–479. [Google Scholar] [CrossRef]
- Tabeta, O.; Mochioka, N. Ophihthidae. In An Atlas of the Early Stage Fishes in Japan; Okiyama, M., Ed.; Tokai Univ Press: Tokyo, Japan, 1998; pp. 58–62. [Google Scholar] [CrossRef]
- Palumbi, S.R. Nucleic acids II: The polymerase chain reaction. In Molecular Systematics; Hilli, D.M., Moritz, C., Mable, B.K., Eds.; Sinauer Associates: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-frendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Horie, N.; Utoh, T.; Yamada, Y.; Okamura, A.; Zhang, H.; Mikawa, N.; Akazawa, A.; Tanaka, S.; Oka, H. Development of embryos and larvae in the common Japanese conger Conger myriaster. Fish. Sci. 2002, 68, 972–983. [Google Scholar] [CrossRef]
- Lee, S.J.; Go, Y.B.; Kim, B.J. Seasonal variation of species composition and distribution of fish eggs and larvae in the western part of Jeju Island, Korea. Korean J. Ichthyol. 2006, 18, 129–140. [Google Scholar]
- Lee, S.J.; Kim, J.B.; Han, S.H. Distribution of mackerel, Scomber japonicus eggs and larvae in the coast of Jeju Island, Korea in spring. J. Korean Soc. Fish. Technol. 2016, 5, 121–129. [Google Scholar] [CrossRef]
- Aoyama, J.; Watanabe, S.; Miller, M.J.; Mochioka, N.; Otake, T.; Yoshinaga, T.; Tsukamoto, K. Spawning sites of the Japanese eel in relation to oceanographic structure and the west Mariana ridge. PLoS ONE 2014, 9, e88759. [Google Scholar] [CrossRef]
- Onda, H.; Miller, J.M.; Takeshige, A.; Miyake, Y.; Kuroki, M.; Aoyama, J.; Kimura, S. Vertical distribution and assemblage structure of leptocephali in the North Equatorial Current region of the western Pacific. Mar. Ecol. Prog. Ser. 2017, 575, 119–136. [Google Scholar] [CrossRef]
- Miller, M.; Nakamura, Y.; Shibuno, T.; Tsukamoto, K. Leptocephali collected in light traps near coral reef habitats of Ishigaki Island in the southern Ryukyu Island chain. Coastal Mar. Sci. 2010, 34, 41–54. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Date | Latitude | Longitude | Sampling Gears | TL (mm) | Stage | Depth |
|---|---|---|---|---|---|---|
| 19 August 2022 | 32°75′ | 127°25′ | MOCNESS | 2.0 | Egg | 20–30 m |
| 17 August 2020 | 31°75′ | 127°25′ | Bongo net | 2.2 | Egg | None |
| 17 August 2020 | 31°75′ | 127°25′ | Bongo net | 2.1 | Egg | None |
| 15 July 2021 | 33°25′ | 127°75′ | MOCNESS | 11.5 | preleptocephalus | 10–20 m |
| 15 July 2021 | 33°25′ | 127°75′ | MOCNESS | 13.5 | preleptocephalus | 10–20 m |
| 15 July 2021 | 33°25′ | 127°75′ | MOCNESS | 20.9 | leptocephalus | 20–30 m |
| 15 July 2021 | 33°25′ | 127°75′ | MOCNESS | 23.5 | leptocephalus | 30–40 m |
| 15 July 2021 | 31°25′ | 127°08′34′′ | MOCNESS | 34.4 | leptocephalus | 20–30 m |
| 16 July 2021 | 31°25′ | 126°75′ | MOCNESS | 30.2 | leptocephalus | 10–20 m |
| 16 July 2021 | 31°25′ | 126°75′ | MOCNESS | 31.5 | leptocephalus | 10–20 m |
| 16 July 2021 | 31°25′ | 126°75′ | MOCNESS | 22.1 | leptocephalus | 20–30 m |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Muraenesox cinereus egg A (1) | ||||||||||
| Muraenesox cinereus egg B (2) | 0.000 | |||||||||
| Muraenesox cinereus leptocephalus A (3) | 0.000 | 0.000 | ||||||||
| Muraenesox cinereus leptocephalus B (4) | 0.000 | 0.000 | 0.000 | |||||||
| Muraenesox cinereus (5) | 0.000 | 0.000 | 0.000 | 0.000 | ||||||
| Muraenesox bagio (6) | 0.047 | 0.047 | 0.047 | 0.047 | 0.047 | |||||
| Echelus uropterus (7) | 0.151 | 0.151 | 0.151 | 0.151 | 0.151 | 0.137 | ||||
| Ophisurus macrorhynchos (8) | 0.172 | 0.172 | 0.172 | 0.172 | 0.172 | 0.156 | 0.086 | |||
| Gnathophis nystromi (9) | 0.166 | 0.166 | 0.166 | 0.166 | 0.166 | 0.157 | 0.141 | 0.157 | ||
| Conger myriaster (10) | 0.223 | 0.223 | 0.223 | 0.223 | 0.223 | 0207 | 0.191 | 0.212 | 0.178 | |
| Anguilla japonica (11) | 0.198 | 0.198 | 0.198 | 0.198 | 0.198 | 0.183 | 0.174 | 0.166 | 0.133 | 0.207 |
| Development | Preleptocephalus | Leptocephalus |
|---|---|---|
| The number of specimens | 2 | 6 |
| Total length, TL (mm) | 11.5–13.5 | 20.9–34.4 |
| In % of total length | ||
| Head length | 13.3–14.7 (13.8 ± 0.5) | 8.4–11.7 (9.8 ± 1.1) |
| Preanal length | 75.8–80.0 (77.9 ± 2.1) | 76.9–88.5 (81.2 ± 4.4) |
| Body depth | 11.8–13.2 (12.5 ± 0.7) | 9.0–11.1 (9.7 ± 0.7) |
| In % of Head length | ||
| Eye diameters | 22.8–23.6 (23.2 ± 0.4) | 19.1–23.8 (20.7 ± 1.6) |
| Snout length | 42.4–43.7 (43.0 ± 0.6) | 42.0–45.7 (43.4 ± 1.2) |
| Upper jaw length | 66.5–66.7 (66.6 ± 0.1) | 56.2–62.5 (58.5 ±2.3) |
| Counts | ||
| Total myomeres | >150 | 145–153 |
| Preanal myomeres | 95–104 | 93–98 |
| Number of nostril | 2 | 2 |
| Dentition formula |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, H.-S.; Yu, H.-J.; Lee, S.-J.; Kang, S.; Kim, J.-K. New Finding of Eggs and Leptocephalus for Muraenesox cinereus off Jeju Island, Korea and Its Relation to Environmental Variables. J. Mar. Sci. Eng. 2023, 11, 655. https://doi.org/10.3390/jmse11030655
Ji H-S, Yu H-J, Lee S-J, Kang S, Kim J-K. New Finding of Eggs and Leptocephalus for Muraenesox cinereus off Jeju Island, Korea and Its Relation to Environmental Variables. Journal of Marine Science and Engineering. 2023; 11(3):655. https://doi.org/10.3390/jmse11030655
Chicago/Turabian StyleJi, Hwan-Sung, Hyo-Jae Yu, Seung-Jong Lee, Sukyung Kang, and Jin-Koo Kim. 2023. "New Finding of Eggs and Leptocephalus for Muraenesox cinereus off Jeju Island, Korea and Its Relation to Environmental Variables" Journal of Marine Science and Engineering 11, no. 3: 655. https://doi.org/10.3390/jmse11030655