

Progress on the Impact of Persistent Pollutants on Marine Turtles: A Review

Department of Earth Sciences, Environment and Resources, University of Naples Federico II, Via Vicinale Cupa Cintia 21, 80126 Naples, Italy
J. Mar. Sci. Eng. 2023, 11(2), 266; https://doi.org/10.3390/jmse11020266
Submission received: 28 December 2022
/
Revised: 17 January 2023
/
Accepted: 18 January 2023
/
Published: 24 January 2023
(This article belongs to the Special Issue The 10th Anniversary of JMSE - Review Collection)
Abstract
:The review examines the most recent advances on the effect of persistent pollutants on sea turtles. Research on this topic is still limited, especially that related to toxicity, since they are protected species and in vivo toxicity studies are difficult, with most studies carried out on deceased individuals. Besides toxicology, the review considers direct links to pollution sources, reproduction impact, health effects and biomarkers of pollution exposure. Little attempts have been made so far to gather data that would provide insight into the causes of the observed health trends. Considering this, studies correlating PPs accumulation with health parameters were also discussed. The review synthesizes the recent progress of the research on these topics and indicates the main urgent need of investigation to limit threats from anthropic pressure.
1. Introduction
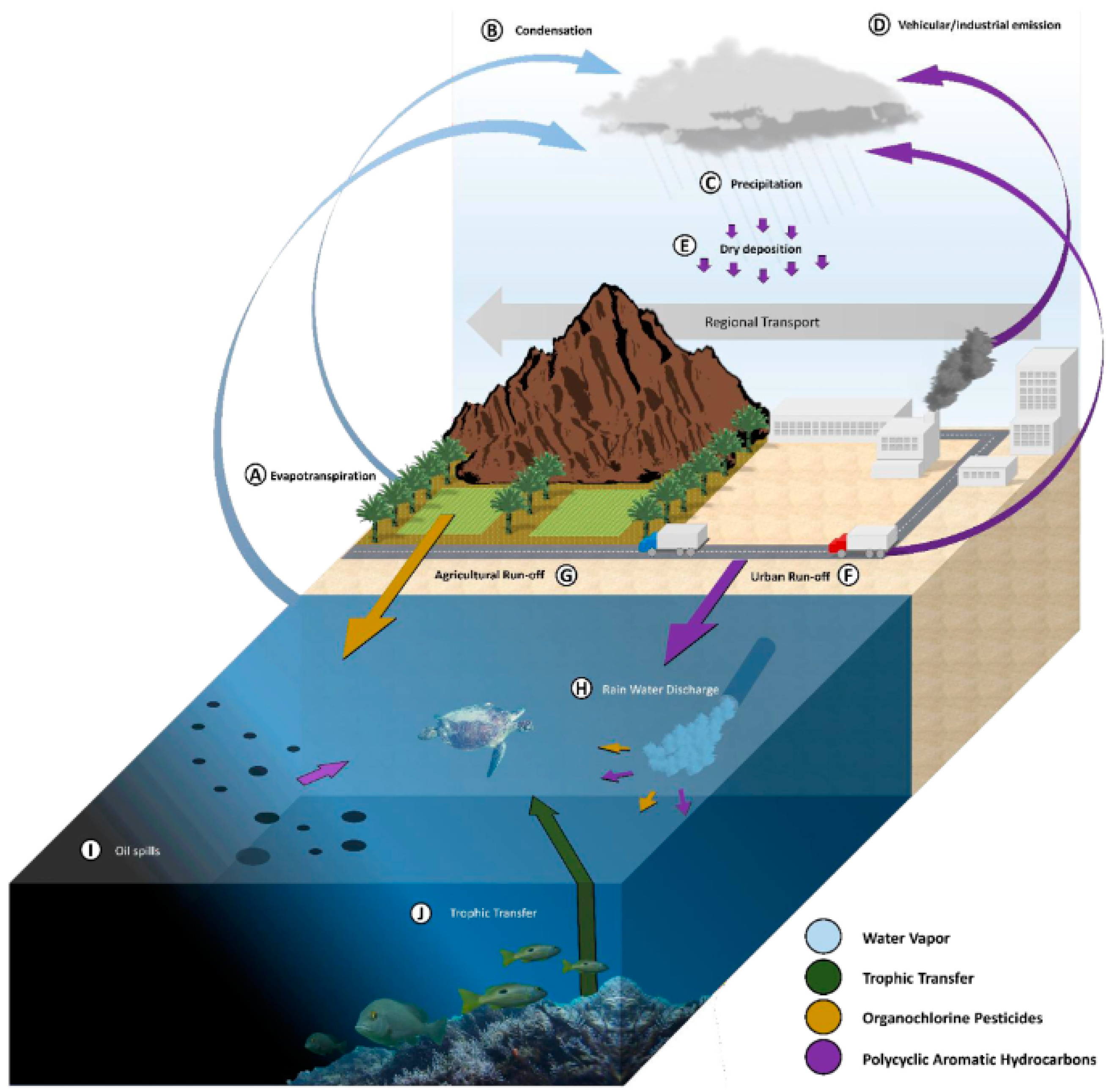
Persistent pollutants, PPs, can be termed as “forever chemicals”, including inorganic and organic compounds such as heavy metals, HMs, polycyclic aromatic hydrocarbons, PAHs, polychlorobiphenyls, PCBs, organochlorine pesticides, and OCPs, which are resistant to environmental degradation through chemical, biological, and photolytic processes. One of the major targets of PPs are oceans where they impact marine biota depending on sources and forms as sound, thermal, photic, plastics, chemical, effluent, and others, Figure 1. Wildlife species, and especially sea turtles as a long-lived species, accumulating high concentrations of chemicals throughout their lifetime, traveling long migratory routes able to travel long distances, may be exposed and accumulate large amounts of PPs from different areas [1,2,3,4]. Exposure routes include water cycle, atmospheric deposition, trophic transfer, plastic ingestion, marine sediments, urban and agricultural run-off, vehicular and industrial emissions, vessel operation, tanker accidents, exploration production, and oil spills [5,6,7,8] (Figure 2). This can be the case for OCPs, which can enter the marine environment through inputs from air and water through agricultural run-off and storm-water discharge (Figure 1). Many PPs are highly hydrophobic and are resistant to degradation and metabolism. This has elicited significant bioaccumulation and magnification [8]. Sea turtles diving into deep and cold water and returning to the surface for breathing, rest, or rewarm are highly exposed to pollution from both the seabed and surface [9]. They are considered endangered species from PPs exposure due to their delayed maturity and longevity and high-level position in the food web. Due to these features, they are considered potential bioindicators of environmental contamination by PPs [10,11,12]. They are distributed from coasts to open ocean [13], migrate covering long distances, and have a complex life cycle [13,14], including terrestrial habitats where females spawn, neritic areas as a refuge for hatchlings’ growth, and the development of juveniles until they reach adulthood and the open ocean for large migrations. They then begin their long back journey to born places where they reproduce, about 25 years after birth, depending on the species, initiating a new cycle [15]. Because of this complex life cycle and longevity, up to 75 years [16], they can bioaccumulate contaminants and be important biomonitoring organisms. This complex life cycle also means that they feed off different organisms: at the juvenile life stage, they live in oceanic areas and behave as opportunistic predators and feed on planktonic biota [2], whereas once they become adults they live in deeper waters, feeding on benthic organisms (Snape et al., 2020) in the form of crustaceous, crabs, and fish [17], and hence bioaccumulate consistent amount of lipophilic pollutants. Their long lifespan and migratory behavior put them at risk of exposure to environmental contaminants that may accumulate over time leading to adverse health outcomes [18,19,20,21,22]. Some of them may weigh up to 900 kg, live at least 45 years grazing in all world’s oceans with wide habitat range from the Indian Ocean to the eastern Pacific, periodically come ashore to either bask or nest, making long journeys up to 10,000 miles per year [23,24] showed as green and loggerhead turtles tend to keep similar migratory routes, foraging areas, and wintering sites, between and within years and after successive breeding migrations and explained the phenomena as due to food resource limitations, territorial defense, or vicinity to wintering sites. This was observed for loggerheads migrations in the Mediterranean Sea [24,25,26,27,28], with individuals foraging in certain specific areas of Mediterraean and then returning to the nesting areas. Uçar et al. [28] reported that loggerheads journey covered between the area between the Island of Ventotene (southwestern Italy) in the Tyrrhenian Sea and the Eastern coast of Turkey in the Eastern Mediterranean Sea.
Due to these features, they can build up many chemicals in tissues, having serious consequences on the conservation of these reptiles, leading over time to immuno-suppression, often associated with fibro-papillomatosis disease, disruption of endocrine function, respiratory inflammation, gastrointestinal ulceration, organ damage, and reproductive failure [2].
There are seven species of sea turtles: leatherback (Dermochelys coriacea), kemps’ ridley (Lepidochelys Kempii), olive ridley (Lepidochelys olivacea), hawksbill (Eretmochelys imbricata), green (Chelonia mydas), loggerhead (Caretta caretta), and flatback (Natator depressus) [29] Figure 3. The International Union for Conservation of Nature’s Red List of Threatened Species [12] includes six of the seven sea turtle species in the ‘threatened’ category, and the deleterious effect of marine pollutants is included among the top 20 research topics for sea turtle conservation [30]. The list classifies as vulnerable (L. olivacea), endangered (C. caretta, C. mydas), and critically endangered (D. coriacea, L. kempii, E. imbricata) (Figure 2). Caretta caretta is the most widespread chelonian in the Mediterranean Sea, using the Italian coasts for nesting and feeding ground [11], and is considered a “flagship species” for biodiversity conservation because it is sensitive to human-induced environmental pollution [31,32,33]. Since turtles are endangered taxa, knowing the level of exposure to PPs provides a framework for assessing the health and improving the conservation of the species. Their presence on the IUCN red list makes the study of the impact of pollution vital.
Figure 1.
Main threats for sea turtles. Source [34].
Figure 1.
Main threats for sea turtles. Source [34].

Figure 2.
POPs exposure paths for marine turtles. Adapted from Yaghmour et al. [7]. With the permission of Elsevier.
Figure 2.
POPs exposure paths for marine turtles. Adapted from Yaghmour et al. [7]. With the permission of Elsevier.

Scientific study of these reptiles began in earnest in the 1950s. Since then, interest in this field has grown, and all six continents (excluding Antarctica) now have active sea turtle research programs [35]. PPs can elicit developmental, reproductive, general health, and immune system effects of sea turtles, even at low concentrations [36] that have long life span, low metabolic rate, and highly efficient conversion of prey into biomass [36]. Studies on the exposure, accumulation, and toxicity of PPs are interesting, since turtles occupy a high trophic niche and can be used as biological indicators of long-term bioaccumulation in the environment. Some studies, in fact [37], report on turtles’ sensitivity to chemical exposures in the environment, and this makes sea turtles early indicators of environmental health issues. Many species such as green sea turtles show sensitivity to different pollutant exposure by assaying immunological and physiological effects [37]. This allows the assessment of the general contaminant status of the environment in which they forage. Research on these animals remains difficult and many issues are understood as direct links to pollution sources, especially non-point sources, toxicology, links between pollutants, and reproduction impact [38]. The amount of data available on PPs contamination of sea turtles and the effect on their health is still relatively limited [32], and many studies have been performed with deceased individuals, significantly limiting the ability to determine if concentrations influenced health state. In the last decade, collection and analysis of blood samples from living turtles enabled to investigate their health more in-depth [35]. Biomarkers can help to address the health status of these threatened reptiles [39].
Figure 3.
Sea turtle species and endangered risk. Source [40].
Figure 3.
Sea turtle species and endangered risk. Source [40].

To date, investigation of the effects of pollutants on sea turtle health has been sparse, leaving a critical research area to be evaluated in their conservation [41]. Little attempts have been made so far to gather data that would provide insight into the causes of the observed health trends. In light of this, studies correlating PPs accumulation with health parameters are clearly needed. The review summarizes the reading of ninety papers from the discovery platform of peer-reviewed literature of science over the last twenty years. The selection was made by keywords such as sea turtles, persistent pollutants, heavy metals, and persistent organic pollutants and toxicity. The review does not collect and summarize huge amounts of data, whereas it has the scope to indicate few and important very recent gaps of the research on these amazing sea water reptiles at risk of extinction. Thus, in light of this, the main review aims were to identify current gaps of the research on the kind of accumulation organs and tissues, biological and environmental PPs effects, relationships between heath state impairment, and causes and toxicity to PPs threshold.
2. Persistent Pollutants
2.1. Heavy Metals
Heavy metals, HMs, are elements with a specific density above 5 g/cm3 and atomic weight greater than 40.04 g/mol [42]. Being non-biodegradable, they can be bioaccumulated in tissues or excreted [43]. Both metals and metalloids form covalent bonds with functional groups of organic molecules, creating lipophilic complexes capable of crossing cell membranes, producing toxic compounds such as tetraalkyl lead or methylated forms of Hg and As. They can also bind to nonmetallic components of cells, as sulphydryl moieties of proteins, causing toxic effects [44]. Moreover, metals can be distinguished in essential for biological function that are toxic at specific thresholds, as aluminum Al, molybdenum Mo, cobalt Co, and nonessential as barium Ba, beryllium Be, and lead Pb, with higher toxicity due to their competition with essential metals [44].
2.2. Factors Driving HMs Accumulation
The bioaccumulation of HMs in sea turtles is governed by biological and environmental factors [45,46].
2.2.1. Biological Factors
Biological factors include body size, age, and sex. Literature displays studies on the relationship between HMs levels in storage organs and animal size through the measurement of the standard carapace length, SCL [46,47,48,49]. Others also consider HMs concentrations in red blood cells, RBCs [50,51,52,53]. Often, body size appeared well-correlated with age. In adults, higher than average levels of metals are often reported: Pb in bone, Cd in kidney, As in kidney, Cd, Hg, Pb, and Zn in muscle, As, Hg, Pb, and Se in central nervous system tissue, may be indicative of chronic exposure [46]. In other cases, no correlation was found, as was the case for the study carried out along the Adriatic Sea coast, where Franzellitti et al. [54] reported a negative correlation (p < 0.05) between SCL and Zn in liver tissue, while Cd, Cu, Fe, Mn, and Ni showed widely variable values with no apparent correlation with SCL. Maffucci et al. [55] reported that Cd, Cu, Hg, and Zn in liver, kidney, and pectoral muscle tissues of loggerhead turtles from the southern Tyrrhenian coast of Italy were not influenced by the size of the specimen, except for Se in the liver, which was negatively correlated with the CCL. Aymak et al. [53] reported that Cu, Zn, Cd, Cr, Mn, and Pb did not show correlation with the CCL, and a highly negative linear relationship (p < 0.05) was found only between the concentration of As in muscle tissue and CCL. While body sizes appeared to be often well-correlated with metal levels, sex has not been shown to be significantly correlated. Maffucci et al. [55] reported that sex had no significant influence on Cd, Cu, Hg, and Zn concentrations in liver, kidney, and pectoral muscle tissues of loggerheads from southern Tyrrhenian coast of Italy.
One fundamental question on HMs accumulation regards the type of target tissue or organ. Table 1 summarizes reports concentrations of HMs in liver and muscles and patterns of abundancy order over the last decades. Most abundant set scaled as Zn > As > Cu > Pb > Mn> Cd> Cr in both liver and muscle, with an evident dominance of Zn and Cu in most studies. García-Fernández et al. [49] assayed metals in liver, kidney, pectoral muscle, brain, and bone of loggerheads from the southwestern Mediterranean coast and found positive correlations between hepatic and renal Cd and hepatic and renal Zn. In other studies, Jerez et al. [46] reported statistically significant positive correlation between Zn in liver and kidney tissue and a statistically significant negative correlation between Se in muscle and liver tissue in dead loggerhead from the Western Mediterranean shores of Murcia (Spain). Saeki et al. [47] reported an inverse correlation with As in muscle for green turtles. Gardner et al. [48] found a positive correlation of Cd, Cu, and Ni in kidney with SCL and Ni with the SCL in both kidney and liver tissue for loggerhead turtles from northwestern Mexico. Aymak et al. [53] reported how Zn showed the highest levels in muscle, 91.09 ± 33.01, range: 59.29–135.78 μg g/g dry wt and liver 83.21 ± 52.17, range: 47.92–186.95 μg/g tissues. Authors also reported that the mean concentrations of Cd, Cu, Pb, Mn, and Cr had higher levels in the liver tissue than in the muscle tissue, but the highest Zn and As levels were found in the muscle tissue, as previously reported in studies by Andreani et al. [56] for Zn in Italy Adriatic and Mediterranean coasts of Italy, Maffucci et al. [55] for Zn in western Italy, Storelli et al. [57] for As in eastern Italy (South Adriatic Sea), Saeki et al. [47] for As from the North Pacific, Jerez et al. [46] for Zn and As in eastern Spain, Caurant et al. [58] for Zn in western France (Atlantic coast of France), Sakai et al. [59] for Zn in Japan, and Sakai et al. [60] for Zn in Japan. Regarding the difference in metal loading among the different tissues in general, heavy metal concentrations in liver and kidney are higher than in the muscle tissue of loggerhead and green turtles [36,59,60]. Arsenic is an exception, since it is present mainly in muscle tissue [47,57,61]. It has been found that As distribution varies with species and growth [47]. Juveniles are carnivorous regardless of the species, whereas when they are adult, species differ from each other in prey items, for instance, green turtles are apparently algicolous, loggerhead turtles tend to be carnivorous, and hawksbill turtles prefer sponges [47]. An interesting study was carried out by Saeki et al. [47] on As accumulation by liver, kidney, and muscle of green, loggerhead, and hawksbill turtles. Authors found how arsenic scaled as muscle > kidney > liver and that As levels were higher in hawksbill, feeding mainly on sponges instead of the two other turtles primarily eating algae and mollusks, normally loading higher amount of As. Arsenic also significantly decreased with standard carapace length as an indicator of growth and the main arsenic compounds was found to be arsenobetaine, detected in the tissues of all the turtles with accumulation differences related to prey items. A similar path was observed by Aymak et al. [53] with As concentration in the order: muscle > liver. This might be due to a specific metabolic mechanism in muscles, as outlined by Saeki et al. [47]. In the same way, Zn was high in loggerheads muscle tissue from the Mediterranean Sea, the Atlantic, and the Pacific Oceans [32]. Storelli and Marcotrigiano [61] reported that As was mostly present in organic form, 90.2–99.7% in muscle, and 81.4–94.2% in liver, and the inorganic As levels in liver were three times higher than in the muscle of loggerheads.
Few studies documented HMs concentrations in the nesting environment and nest contents of sea turtles as eggshells, yolk, embryonic livers of hatchling specimens [62,63,64].
If liver, muscles, and red blood cells are the most widely studied target of accumulation, other targets such as salt glands, SG, have been recently individuated. SG is a gland placed near turtle eyes that allow them to drink sea water and excrete salt in high concentrations [22]. This represents an important adaptation system that plays an important physiological function in turtles regarding osmotic, electrolyte, mineral, and hydration balance [22]. Little research has been conducted on contaminant accumulation and excretion in this organ [22] and mainly for inorganic pollutants. Perrault et al. [22] hypothesized that SG might be a potential surrogate for evaluating inorganic contaminants levels in blood in sea turtles. Outside of the normal osmoregulatory function, the gland can store and excrete inorganic contaminants [60]. Perrault et al. [22] studied the accumulation of As, Cr, Cd, Pb, Hg, Se, in RBCs and salt gland secretions, SGSs, of nesting leatherbacks and collected salt glands from different life stage classes of dead stranded leatherbacks from the western Atlantic Ocean and reported that SG, Cd, and Hg concentrations increased with CCL, suggesting this organ as a potential target for accumulation.
Leatherbacks ingests excessive amounts of salts due to their diet of gelatinous zooplankton, and SG glands serve to maintain osmotic and ionic homeostasis. Authors did not find correlations between the inorganic contaminants in RBCs and SGS, as well as significantly higher concentrations in RBCs due to the affinity to the heme group. SG was shown to be a potential target for inorganics with positive correlation of SGS contaminant concentrations with increasing CCL. The authors indicated different physiological mechanisms, causing the distribution of metals in blood and SGSs.
2.2.2. Environmental Factors
Typically, environmental factors are type of forages, travelled routes, or a combination of these, and the environment from which the turtle is captured. There are authors [54,58,65] retaining the diet as the main source of exposure with differences among species. This is important when turtles move through water of more industrialized regions and metal burden in tissues become dependent by feeding habits, life cycles, and chronic exposure in these areas. For instance, loggerheads, being at the top of their food web, are exposed to high levels of HMs resulting from bio-amplification processes across the trophic chain [54]. Aymak et al. [53] hypothesized that the concentrations of metals in loggerheads tissues depend mainly on their feeding habits in different areas. Tomas et al. [66] reported three different foraging habitats of loggerheads as they grow: the pelagic, the neritic, and the benthic-demersal phases. Juvenile green individuals accumulate higher metal levels, but as they grow, and there is a decrease in contaminant intake by dilution effect [48]. Aymak [53] related feeding habits and chronic exposure with As, Cd, Cr, Cu, Mn, Pb, and Zn in the muscle and liver of loggerheads from Kazanlı beach, Mersin, Turkey. Authors highlighted the dominant presence of Zn ˃ As ˃ Cu in both tissues (Table 1). Muscle accumulated higher levels of As and Zn than liver tissue, whereas the others were dominated in the liver, but with significant differences only for Pb. The authors documented a negative correlation between turtle size and muscle As levels due to chronic exposure and hypothesized a relation between As and the health state. If, as it seems, sea turtles are highly sensitive to metal exposure, some metals may be used as markers to individuate the feeding areas and indicators of the turtle–environment interaction [54].
Some species as green turtles forage very close to shores and hence are capable of accumulating high levels of elements. Villa et al. [67] used a clinical reference interval, RI, method to set up exposure baseline limits for essential and non-essential elements and assayed blood from healthy subadults foraging in a remote and offshore part of the Great Barrier Reef in Australia. This baseline was compared with metal levels in coastal populations, foraging in areas dominated by agricultural, urban, and military activities. Authors found clear habitat-specific differences in blood metal profiles, which were reported respective to the offshore blood baseline. In particular, Co blood concentrations were 4–25 times above RI, levels known to cause acute effects in many vertebrates. The higher levels of Co, Sb, and Mn in onshore individuals significantly correlated with markers of acute inflammation and liver dysfunction.
Barraza et al. [6] highlighted how the kind of urbanized habitat influences metal concentrations in cute and red blood cells of green sea turtles. Cd and Se showed location-specific differences, supporting the hypothesis that coastal cities can increase trace metal exposure to local green turtles. In the same species, Jan et al. [68] evidenced how the concentrations of Mn, Zn, Se, As, Cd, and Pb in eggshells were significantly correlated with those in coral sand sediments. In eggshells, metals were rated as follows (μg/g): Sr (41.3) > Zn (20.3) > Cu (12.8) > Fe (4.92) > Al (4.37) > Se (2.44) > Mn (0.91) > Cr (0.81) > Ba (0.44) > Pb (0.14) > As (0.08) > Cd (0.02), with Cu above the toxic reference value for bird eggs. Identifying spatial variability of metal exposure may improve the understanding of habitat utilization by turtles in highly urbanized estuaries. This is what provided evidence to Komoroske et al. [37], who estimated the levels of metals in organisms and sediments feeding green sea turtles in San Diego Bay and found Ag, Cd, Cu, Mn, Se, and Zn with the highest bioaccumulation levels.
2.3. Organic Pollutants
Among organic contaminants, PCBs, PAHs, and OCPs are those more frequently studied in the literature [1,2,3,11,17,21,31,33,36,69,70,71,72,73,74,75,76,77]. They were classified as persistent organic pollutants, POPs, under the Stockholm Convention, due to their high persistence in the environment [78].
PCBs are chemically stable aromatic chlorinated hydrocarbons that were commercialized in 1930 and have banned since the early 1970s [79]. They soon found wide usage for their specific and unique features such as chemical inertness, low electrical conductance, resistance to fire, and thermal breakdown [80]. Even though more than 50 years have passed since their prohibition, PCBs are commonly detected in the environment, being volatile and easily transported over long distances and lipophilic [3,81]. In biota they bioaccumulate in fatty foods and animal adipose tissue [80,82], and are often detected in wildlife of terrestrial and marine habitats [82], biomagnifying in animals of higher trophic levels. They encompass 209 congeners based on their chlorination and can be divided in dioxin-like PCBs, DL-PCBs, having planar molecular structure, and non-dioxin-like PCB congeners, NDL-PCB, with a non-planar molecular structure [83].
Similar to PCBs, PAHs are mutagenic, carcinogenic, and teratogenic ubiquitous pollutants POPs that can be detected quite often in marine and coastal environments [84,85]. With respect to PCBs, PAHs are frequently detected in marine benthic invertebrates as mussels and clams [86], but the evidence in higher trophic level predators is limited [22,87], where they are often metabolized, and this reduces the biomagnification risk [88]. Meador et al. [89] produced one of the first report on the PAHs presence in sea turtles; Camacho et al. [90] reported that PAHs represent one of many major environmental issues for sea turtles in the Mediterranean Sea.
The other important class of POPs are OCPs, which are synthetic pesticides that have been widely used all over the world for managing pests and disease vectors in agricultural and public health applications. Similarly to PCBs, these compounds have been banned in developed countries, but their abuse usage is rising in practice across the continents [2,72].
Bioaccumulation of POPs
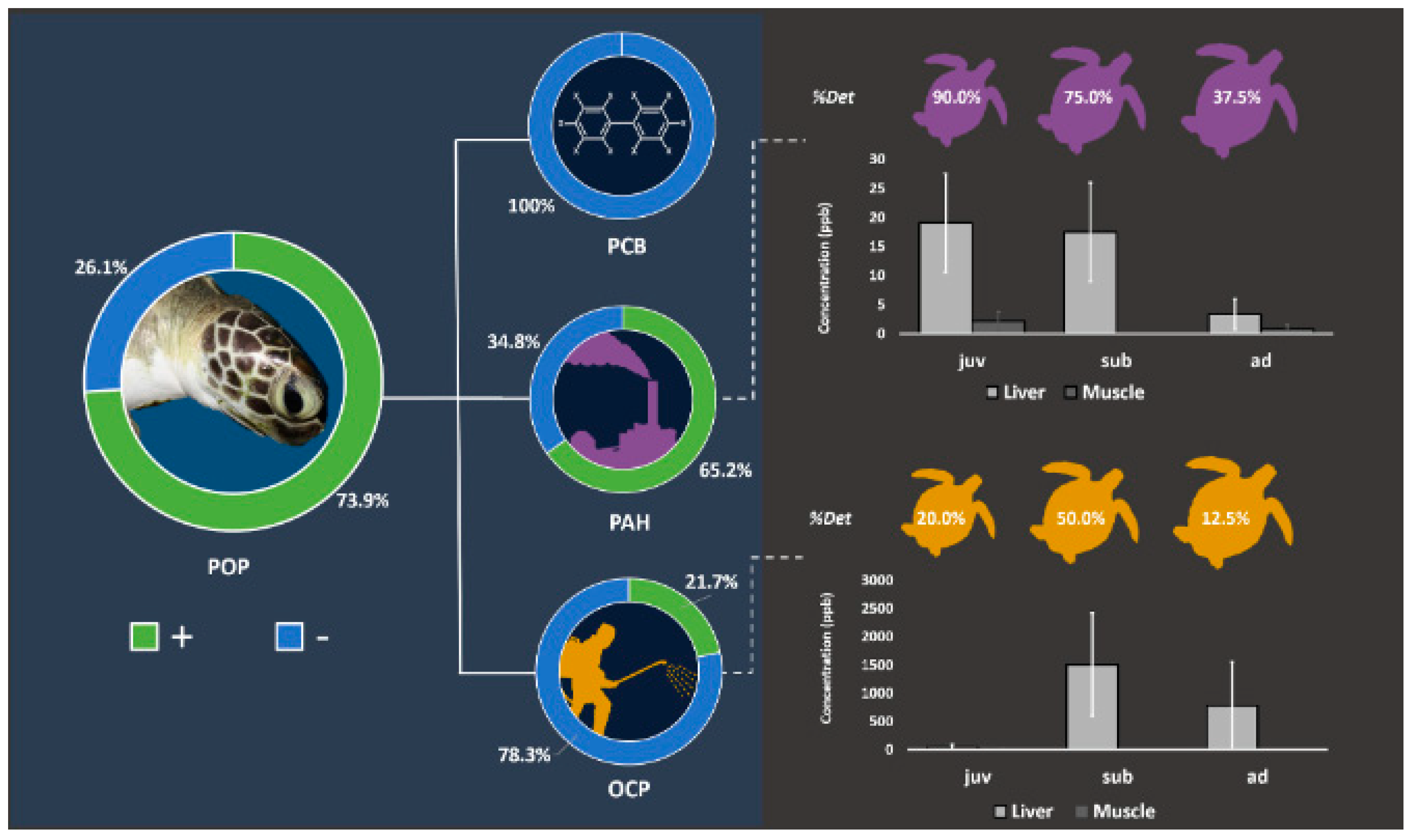
Two mayor key factors that drive accumulation are time and space. Finlayson et al. [91], using a species-specific in vitro bioassay with primary cells, revealed that the levels of organic pollutants extracted by green turtles’ blood significantly matched the spatial contaminant distribution and increased over exposure time. However, the approach does not discriminate pollutants, since it is based on the QuEChERS (quick, easy, cheap, effective, rugged, and safe) method validated for turtle blood by Dogruer et al. [92], which is a non-targeted extraction for the POPS, pesticides, pharmaceuticals, polycyclic aromatic hydrocarbons (PAHs), and flame-retardants assay. Table 2 reports on the levels of POPs in liver, muscle, fat, eggs, salt glands, and blood of green and loggerhead turtles. It seems that among POPs, and tissues such as muscles, fat, and blood, OCPs accumulated at higher rates and up to 563 ng/g [7] Figure 4.
3. Bioindicators of Pollution
Even though sea turtles can be considered good bioindicators for marine pollution, little knowledge exists on sea turtles’ health effects [14]. Within PPs, HMs and POPs build complex and combined effects [14]. In the case of HMs, it is well-known that common animal powerful mechanisms of defence are embedded in antioxidant, detoxification, and transport mechanisms, implying the production of glutaredoxin (GRx) [97], ferritin (FER) [98], and metallothioneins (MTs) [99,100]. These enzymes sequester metals and protect against oxidative stress by scavenging free radicals [101]. MTs are activated by metal regulatory transcription factor 1 (MTF1), which also regulates other proteins/genes such as ferritin, selenoproteins, thioredoxin reductase, and glutamate-cysteine ligase [102], and can also be used to assess other points of oxidative stress. Under this view, Morão et al. [14] assayed metal and metalloid contamination in the blood of green sea turtles and identified signs of contamination by analysing molecular stress responses. The authors identified effective biomarkers associated with metal stress and revealed how turtles adapt to metal oxidative stress and damage through the increased expression of antioxidants and other protectors.
4. PPs Toxicity
One of the major issues that need to be investigated and enlarged is sea turtle PPs toxicity. Toxicity varies with water quality and among species [43]. Developmental stage, dietary factors, physiology, and biochemical functions all influence the degree to which PPs are toxic. Concentration, duration of exposure, and chemical speciation of the element also play important roles in toxicity. Water quality influences toxicity through pH, hardness, and salinity interactions.
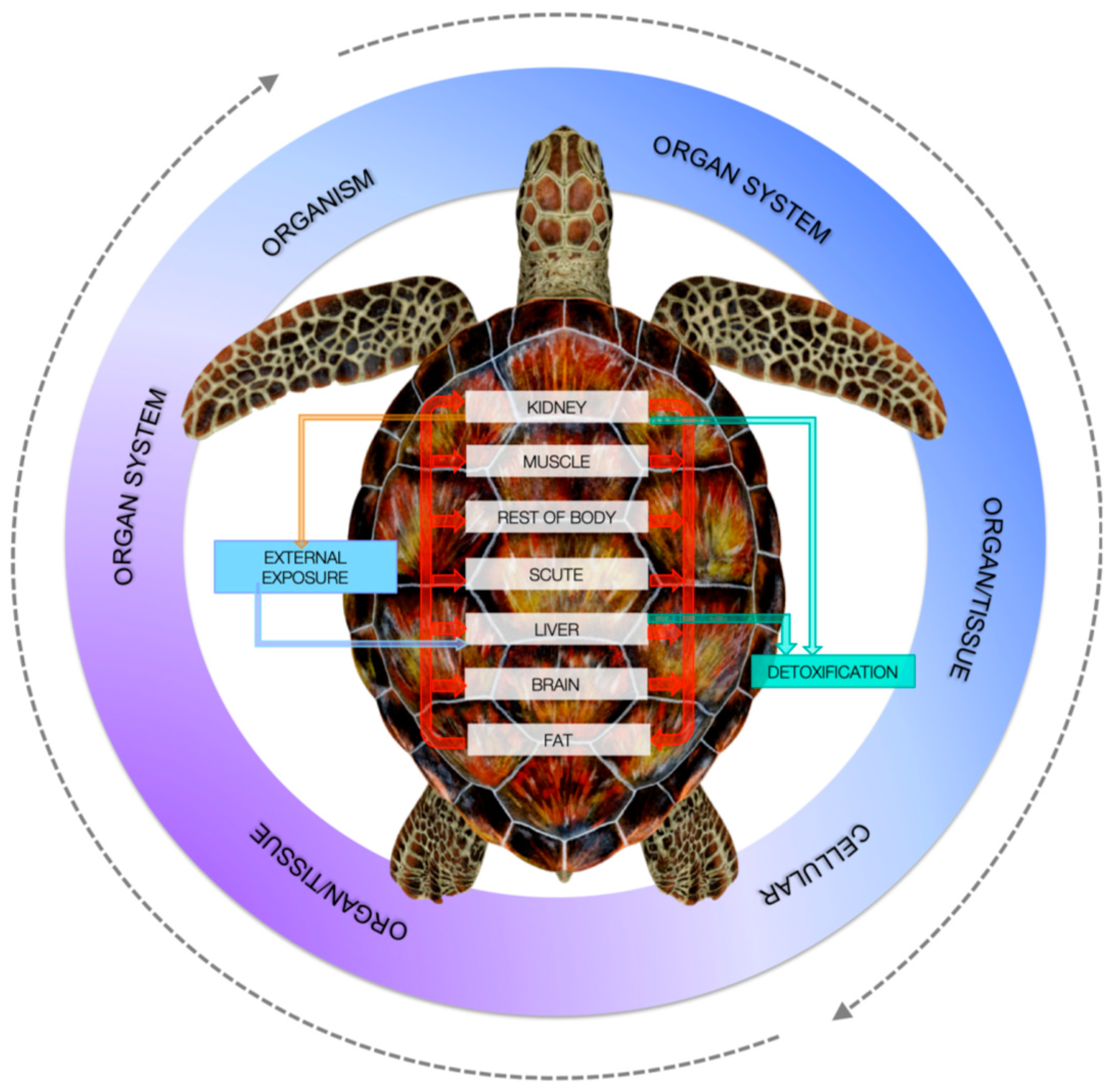
In the case of metals, one of the most threatening elements is chromium, which can be highly toxic for turtles besides humans. Chromium is considered as one of the main marine global metal contaminants that can have toxic impacts across species [41]. Speer et al. [41] studied the cytotoxicity and genotoxicity of particulate Cr(VI) in humans and leatherbacks lung fibroblasts and found that particulate Cr(VI) induced cytotoxicity in leatherback cells comparable to human cells. Kaska et al. [63,103] considered that HMs in turtle tissues are likely to affect the health of loggerhead turtles in the Mediterranean. The reason why PPs pollutant health effects need to be investigated more deeply lies in the difficulty to set up exposure threshold limits, which are largely lacking [92]. Dogruer et al. [92] studied the effects of Cd in green sea turtles to test a mechanistic model named physiologically-based kinetic (PBK) to test contaminant risk assessment. The model was built on species, chemical lifetime bioaccumulation, in vitro and in vivo toxicity threshold assay, and chemical risk quantification (Figure 5). Seven targets were considered as kidney, muscle, scute, liver, brain fat, and the rest of body. As has been stated above, in the rest of body it can be included—for example, salt glands might possess outstanding enrichment power of PPs as PAHs [96]. Dogruer et al. [92] revealed that Cd kidney toxicity was age-dependent and varied with physiology and feeding ecology, and set up a steady-state exposure threshold of 0.1 μg/g dry weight Cd in forages. Thus, even with some uncertainty, threshold limits can be set up. PCBs may affect the immune system of sea turtles, causing anaemia, hepatic damage, and immunomodulation [22,33,77].
Within POPs, PAHs may have wide mechanisms of toxicity, including modulation in target gene expression and DNA methylation [22]. Chronic or high-level exposure to PAHs or crude oil may cause serious health and reproductive problems [104], with toxicity varying between species and largely depending on the route of exposure, sex, and life stage of the organism. Hall [105] evidenced that in some cases enzymatic activities, as microsomal enzymes, are capable of metabolizing PAHs into metabolites that are even more toxic than the original compound [106,107]. Thus, for sea turtle conservation purposes, it is essential to determine how they respond to PAHs exposure in different tissues and organs.
5. Toxicity Assessment
Most of the toxicity assessment research has been conducted on tissue samples, including the liver, kidney, and muscle. Occasionally, studies included analysis of the stomach, lung, adipose tissue, pancreas, and spleen. All tissue samples were obtained from deceased turtles. Many studies used tissue only from turtles where time of death was determined to be less than 24 h. Loggerhead turtle is one of the most studied species along the Mediterranean coast [46,49,54,55,59,60,70], the Atlantic coasts of France [60], the Baja California Peninsula in northwestern Mexico [48], Japanese waters [59,60], the North Pacific [60], and south-eastern Queensland, Australia [108]. These studies, in addition to reporting PPs concentrations toxicity effects, provide limited evidence on cause–effect paths due to the objective difficulty to test toxicity on living sea turtle individuals and ethical and logistical constraints [109]. Kaska et al. [103], in a monitoring assay of the Mediterranean Turkish coast, documented the possible cause of mortalities of stranded sea turtles, and the statistical linkages of PPs in the tissues of dead stranded loggerhead and green sea turtles. To overcome these constraints, a feasible cost-effective, reproducible way could be represented by in vitro assays as an alternative to live animal toxicity testing. The main issue of this approach is if the cell culture represents the toxicity in that species. Finlayson et al. [109] studied the cytotoxic response of primary skin fibroblasts green and loggerhead turtles in response to contaminants. The authors found that validated cell cultures may be useful in toxicity testing with limited differences in species chemical sensitivity, facilitating rapid assessment of chemical risk. Different in vitro assays have been drawn for assessing the impacts of contaminants in sea turtles. One regards the extraction of organic contaminants from blood and the evaluation of the biological effects on rat and human cells (Jin et al. [110] and Dogruer et al. [92]), and the main limitation of the method is that it is not species-specific. One of the main problems when using single cell cultures is if they adequately represent the toxicity in that species. Keller et al. [111] overcame this problem by examining the effects of organochlorine contaminants on loggerhead turtles using peripheral blood leukocytes (PBL) in several individuals. In the same way, Webb et al. [112] used primary skin fibroblast cultures from loggerhead turtles for testing the effects of perfluorooctanoic acid and benzo[a]pyrene, evaluating cell viability and cytochrome P4501A expression. The validation of primary cell cultures in toxicity bioassays lies in the establishment of the variation in the response of cells from individuals with different ages and belonging to various populations. Since sea turtle are a threatened species, for this reason, direct exposure experiments would be difficult to conduct with collection and sacrifice restrictions. Laboratory experiments are limited by legal, ethical, and logistical constraints [38]. Cell-based, or in vitro, studies present good opportunities to study toxicology in threatened or endangered species [109]. Few studies exist using in vitro or molecular methods, such as estrogenic receptor expression, cell culture exposures, and DNA expression studies. Previous studies in sea turtles have assessed estrogenic endocrine disruption by assaying DNA expression of estrogenic receptors within different tissues [113]. While there are some limitations to in vitro toxicity testing, including the absence of feedback processes, low cell density, the use of a single cell type, and artificial culture conditions [114], in vitro bioassays represent a more ethical and sensitive approach [115]. Normally, reproductive effects include maternal transfer, developmental abnormalities, altered sexual morphology, sex reversal, and sex steroid alterations. Another possible route to assess toxicity is to compare toxicological data with those on humans, for the identification of the exposure to a specific chemical and mechanistic insights. The output of the comparison can then be used in risk assessment, conservation efforts, and regulations to protect humans, wildlife, and the environment. Very few studies have been performed on the comparison of toxicity between sea turtle cells and human cells following PPs exposure. This is the case for chromium which, in the form of Cr (VI), is one of the most common marine metal contaminants [116]. Cell culture studies showed its cytotoxicity and genomic instability in human lung cells [117] with the metal capable to induces DNA double strand breaks and inhibition of breaks repair [118]. The mechanisms leading to these endpoints remain unknown. Young et al. [119] compared the effects of Cr between hawksbill sea turtle (Eretmochelys imbricata) skin cells and human skin cells and showed a similar response in the two cell types. Speer et al. [41] assessed the cytotoxic and genotoxic effects of particulate Cr in leatherback fibroblasts after acute and prolonged exposures and compared these results with data in human lung fibroblasts. For sea turtles, it has been found that in the case of POPs within the mother and the developing embryo, the less lipophilic compounds are absorbed more readily at first and, later on, those that are more lipophilic are absorbed [38]. The more lipophilic compounds get absorbed towards the end of egg and embryo development. This is important when testing patterns of lower lipophilic congener PCBs, which can have higher toxic equivalency factors than the more lipophilic compounds [120].
6. Health Parameters
Monitoring health indicators in sea turtles is relatively new, with preliminary studies of baseline parameters being initiated fewer than 20 years ago [50,51,121,122]. Health parameters in sea turtles have been monitored in several species, but the ability to control for external influences has been limited. This makes it extremely difficult to determine what factors are affecting the health of these organisms. In fact, little research has been done regarding health parameters to potential causes.
Investigations regarding baseline health parameters have been conducted in several species of sea turtle, including C. mydas [123], D. coriacea [124], and C. caretta [125]. The first of these studies was conducted on two populations of healthy juvenile C. mydas turtles, with and without fibropapillomatosis, Figure 6 [123]. Individuals were categorized into age/size classes and assigned a fibropapilloma severity score, and 25 biochemistry analytes were examined in blood. The authors speculated that the difference in enzymatic values between the two healthy groups was the result of a more efficient stress response. Fibropapillomatosis was known to be associated with chronic stress and immunosuppression [121].
Another study was conducted on leatherback turtles [124], assaying baseline hematology, plasma biochemistry, and plasma protein values of clinically healthy nesting D. coriacea. Samples were also tested for organochlorine contaminants and polychlorinated biphenyls, arsenic, and mercury. Most of the chemicals were found below detectible limits, possibly due to the dietary preference of leatherbacks for jellyfish, which occupy a low trophic level and subsequently do not bioaccumulate in high levels, and did not cause any significant alteration of the enzymatic plasma parameters and cause health effects.
Day et al. [51] conducted a study on C. caretta from the eastern coast of the United States to look at the health implications of blood mercury concentrations. It was found that total blood mercury concentrations were positively related to measures of hematocrit and CPK, with Hg levels altering physiological parameters.
7. Conclusions
The review outlines some recent progress on PPs effect on sea turtles. In the short term it will be very demanding to study PPs concentrations toxicity effects for different storing organs, providing evidence on cause–effect paths. In the long term, it will be important to set up methods defining exposure threshold toxicity limits for different PPs, and to have deeper knowledge on the impact of PPs on reproduction via direct and indirect pathways of exposure and clinical health parameters. It will be important to have knowledge of health parameters and their relationships with different causes in several species and under different habitats. It will be important to determine what factors affect the health of these organisms and to design new methods for assessing biomarkers of endocrine disruption, examining how PPs affect reproduction and sex ratio. A better understanding of the effects of PPs on the health of these endangered species will facilitate more effective monitoring and protection in the future, enabling us to conserve these fascinating creatures more effectively.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barraza, A.D.; Komoroske, L.M.; Allen, C.D.; Eguchi, T.; Gossett, R.; Holland, E.; Lawson, D.D.; LeRoux, R.A.; Lorenzi, V.; Seminoff, J.A.; et al. Persistent organic pollutants in green sea turtles (Chelonia mydas) inhabiting two urbanized Southern California habitats. Mar. Pollut. Bull. 2020, 153, 110979. [Google Scholar] [CrossRef] [PubMed]
- Esposito, M.; Canzanella, S.; Iaccarino, D.; Bruno, T.; Esposito, E.; Di Nocera, F.; Arienzo, M.; Ferrara, L.; Gallo, P. Levels of non-dioxin-like PCBs (NDL-PCBs) in liver of loggerhead turtles (Caretta caretta) from the Tyrrhenian Sea (Southern Italy). Chemosphere 2022, 308, 136393. [Google Scholar] [CrossRef] [PubMed]
- Nava Montes, A.D.; Espinosa Reyes, G.; Flores Ramírez, R.; Ramírez Romero, P. Persistent organic pollutants in kemp’s ridley Sea turtle Lepidochelys kempii in playa rancho nuevo sanctuary, Tamaulipas, Mexico. Sci. Total Environ. 2020, 739, 140176. [Google Scholar] [CrossRef] [PubMed]
- Canzanella, S.; Danese, A.; Mandato, M.; Lucifora, G.; Riverso, C.; Federico, G.; Gallo, P.; Esposito, M. Concentrations of trace elements in tissues of loggerhead turtles (Caretta caretta) from the Tyrrhenian and the Ionian coastlines (Calabria, Italy). Environ. Sci. Pollut. Res. Int. 2021, 28, 26545–26557. [Google Scholar] [CrossRef] [PubMed]
- Arienzo, M.; Donadio, C.; Mangoni, O.; Bolinesi, F.; Stanislao, C.; Trifuoggi, M.; Toscanesi, M.; Di Natale, G.; Ferrara, L. Characterization and source apportionment of polycyclic aromatic hydrocarbons (PAHs) in the sediments of gulf of Pozzuoli (Campania, Italy). Mar. Pollut. Bull. 2017, 124, 480–487. [Google Scholar] [CrossRef]
- Arienzo, M.; Toscanesi, M.; Trifuoggi, M.; Ferrara, L.; Stanislao, C.; Donadio, C.; Villari, G.; De Vico, G.; Carella, F. Contaminants bioaccumulation and pathological assessment in Mytilus galloprovincialis in coastal waters facing the brownfield site of Bagnoli, Italy. Mar. Pollut. Bull. 2019, 140, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Yaghmour, F.; Samara, F.; Alam, I. Analysis of polychlorinated biphenyls, polycyclic aromatic hydrocarbons and organochlorine pesticides in the tissues of green sea turtles, Chelonia mydas, (Linnaeus, 1758) from the eastern coast of the United Arab Emirates. Mar. Poll. Bull. 2020, 160, 111574. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.A.; Lutz, P.L. Marine turtles as sentinels of ecosystem health: Is fibropapillomatosis an indicator? EcoHealth 2004, 1, 275–283. [Google Scholar] [CrossRef]
- Pasanisi, E.; Cortés-Gómez, A.A.; Pérez-López, M.; Soler, F.; Hernández-Moreno, D.; Guerranti, C.; Martellini, T.; Fuentes-Mascorro, G.; Romero, D.; Cincinelli, A. Levels of perfluorinated acids (PFCAs) in different tissues of Lepidochelys olivacea sea turtles from the Escobilla beach (Oaxaca, Mexico). Sci. Tot. Environ. 2016, 572, 1059–1065. [Google Scholar] [CrossRef]
- Lutcavage, M.E.; Plotkin, P.; Witherington, B.; Lutz, P.L. Human impacts on sea turtle survival. In The Biology of Sea Turtles; Lutz, P.L., Musick, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 387–409. [Google Scholar]
- Perugini, M.; Giammarino, A.; Olivieri, V.; Guccione, S.; Lai, O.R.; Amorena, M. Polychlorinated biphenyls and organochlorine pesticide levels in tissues of Caretta caretta from the Adriatic Sea. Dis. Aquat. Org. 2006, 71, 155–161. [Google Scholar] [CrossRef]
- IUCN. IUCN Red List of Threatened Species. Version 2021. 2021. Available online: https://www.iucnredlist.org/species/4615/11037468 (accessed on 24 December 2022).
- Pike, D.A. Climate influences the global distribution of sea turtle nesting. Global Ecol. Biogeogr. 2013, 22, 555–566. [Google Scholar] [CrossRef]
- Morão, F.C.; Lemos, M.F.L.; Félix, R.; Vieira, S.; Barata, C.; Novais, S.C. Stress response markers in the blood of S˜ao Tom´e green sea turtles (Chelonia mydas) and their relation with accumulated metal levels. Environ. Poll. 2022, 293, 118490. [Google Scholar] [CrossRef] [PubMed]
- Luschi, P.; Lutjeharms, J.R.E.; Lambardi, P.; Mencacci, R.; Hughes, G.R.; Hays, G.C. A review of migratory behaviour of sea turtles off southeastern Africa. S. Afr. J. Sci. 2006, 102, 51–58. [Google Scholar]
- Mayne, B.; Tucker, A.D.; Berry, O.; Jarman, S. Lifespan estimation in marine turtles using genomic promoter CpG density. PLoS ONE 2020, 15, e0236888. [Google Scholar] [CrossRef] [PubMed]
- Cammilleri, G.; Calvaruso, E.; Pantano, L.; Cascio, G.L.; Randisi, B.; Macaluso, A.; Vazzana, M.; Caracappa, G.; Giangrosso, G.; Vella, A.; et al. Survey on the presence of non-dioxin-like PCBs (NDL-PCBs) in loggerhead turtles (Caretta caretta) stranded in south Mediterranean coasts (Sicily, Southern Italy). Environ. Toxicol. Chem. 2017, 36, 2997–3002. [Google Scholar] [CrossRef] [PubMed]
- Stewart, K.R.; Keller, J.M.; Templeton, J.R.; Kucklick, J.R.; Johnson, C. Monitoring persistent organic pollutants in leatherback turtles (Dermochelys Coriacea) confirms maternal transfer. Mar. Pollut. Bull. 2011, 62, 1396–1409. [Google Scholar] [CrossRef]
- Perrault, J.R. Assessment of mercury and selenium concentrations in tissues of stranded leatherback sea turtles (Dermochelys coriacea). J. Herp. Med. Surg. 2012, 22, 76–85. [Google Scholar] [CrossRef]
- Perrault, J.R. Mercury and selenium ingestion rates of Atlantic leatherback sea turtles (Dermochelys coriacea): A cause for concern in this species? Mar. Environ. Res. 2014, 99, 160–169. [Google Scholar] [CrossRef]
- Perrault, J.R.; Lehner, A.F.; Buchweitz, J.P.; Page-Karjian, A. Evidence of accumulation and elimination of inorganic contaminants from the lachrymal salt glands of leatherback sea turtles (Dermochelys coriacea). Chemosphere 2019, 217, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Cocci, P.; Mosconi, G.; Bracchetti, L.; Nalocca, J.M.; Frapiccini, E.; Marini, M.; Caprioli, G.; Sagratini, G.; Palermo, F.A. Investigating the potential impact of polycyclic aromatic hydrocarbons (PAHs) and polychlorinated biphenyls (PCBs) on gene biomarker expression and global DNA methylation in loggerhead sea turtles (Caretta caretta) from the Adriatic Sea. Sci. Total Environ. 2018, 619–620, 49–57. [Google Scholar] [CrossRef]
- Dodge, K.L.; Galuardi, B.; Miller, T.J.; Lutcavage, M.E. Leatherback turtle movements, dive behavior, and habitat characteristics in ecoregions of the Northwest Atlantic Ocean. PLoS ONE 2014, 9, e91726. [Google Scholar] [CrossRef] [PubMed]
- Broderick, A.C.; Coyne, M.S.; Fuller, W.J.; Glen, F.G.; Godley, B.J. Fidelity and over-wintering of sea turtles. Proc. R. Soc. B 2007, 274, 1533–1538. [Google Scholar] [CrossRef] [PubMed]
- Bentivegna, F. Intra-Mediterranean migrations of loggerhead sea turtles (Caretta caretta) monitored by satellite telemetry. Mar. Biol. 2002, 141, 795–800. [Google Scholar]
- Camiñas, J.A. Sea turtles of the Mediterranean Sea: Population dynamics, sources of mortality and relative importance of fisheries impacts. FAO Fish. Rep. 2004, 738, 27–84. [Google Scholar]
- Maffucci, F.; Kooistra, W.H.C.F.; Bentivegna, F. Natal origin of loggerhead turtles, Caretta caretta, in the neritic habitat off the Italian coasts, Central Mediterranean. Biol. Cons. 2006, 127, 183–189. [Google Scholar] [CrossRef]
- Uçar, A.H.; Maffucci, F.; Ergene, S.; Ergene, M.; Katılmış, Y.; Başkale, E.; Kaska, Y.; Hochscheid, S. A Stranded Loggerhead Turtle Tracked by Satellite in Mersin Bay, Eastern Mediterranean Sea, Turkey. Mar. Turt. Newsl. 2018, 155, 12–14. [Google Scholar]
- Lutz, P.L. Salt, water, and pH balance in sea turtles. In The Biology of Sea Turtles; Lutz, P.L., Musick, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 1997; Volume I, pp. 343–361. [Google Scholar]
- Hamann, M.; Godfrey, M.H.; Seminoff, J.A.; Arthur, K.; Barata, P.C.R.; Bjorndal, K.A.; Bolten, A.B.; Broderick, A.C.; Campbell, L.M.; Carreras, C.; et al. Global research priorities for sea turtles: Informing management and conservation in the 21st century. Endang. Species Res. 2010, 11, 245–269. [Google Scholar] [CrossRef] [Green Version]
- Corsolini, S.; Aurigi, S.; Focardi, S. Presence of polychlorobiphenyls (PCBs) and coplanar congeners in the tissues of the Mediterranean Loggerhead Turtle Caretta caretta. Mar. Pollut. Bull. 2000, 40, 952–960. [Google Scholar] [CrossRef]
- D’llio, S.; Mattei, D.; Blasi, M.F.; Alimonti, A.; Bogialli, S. The occurrence of chemical elements and POPs in loggerhead turtles (Caretta caretta): An overview. Mar. Poll. Bull. 2011, 62, 1606–1615. [Google Scholar] [CrossRef]
- Camacho, M.; Boada, L.D.; Orós, J.; López, P.; Zumbado, M.; Almeida- González, M.; Luzardo, O.P. Monitoring organic and inorganic pollutants in juvenile live sea turtles: Results from a study of Chelonia mydas and Eretmochelys imbricata in Cape Verde. Sci. Total Environ. 2014, 481, 303–310. [Google Scholar] [CrossRef]
- Sea Turtle Posters Infographics. Available online: https://rollingharbour.com/marine-life-2/sea-turtles/sea-turtle-info-posters/ (accessed on 24 December 2022).
- Register, A.L. Effects of Heavy Metal Pollution on the Loggerhead Sea Turtle Loma Linda University Electronic Theses, Dissertations & Projects 2011. Master’s Thesis. Available online: http://scholarsrepository.llu.edu/etd (accessed on 24 December 2022).
- Perrault, J.R.; Stacy, N.I.; Lehner, A.F.; Poor, S.K.; Buchweitz, J.P.; Walsh, C.J. Toxic elements and associations with hematology, plasma biochemistry, and protein electrophoresis in nesting loggerhead sea turtles (Caretta caretta) from Casey Key, Florida. Environ. Poll. 2017, 231, 1398–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komoroske, L.M.; Lewison, R.L.; Seminoff, J.A.; Deustchman, D.D.; Deheyn, D.D. Trace metals in an urbanized estuarine sea turtle food web in San Diego Bay, CA. Sci. Total Environ. 2012, 417–418, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Barraza, A.D.; Finlayson, K.A.; Leusch, F.D.L.; van de Merwe, J.P. Systematic review of reptile reproductive toxicology to inform future research directions on endangered or threatened species, such as sea turtles. Environ. Poll. 2021, 286, 117470. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Góomez, A.A.; Morcillo, P.; Guardiola, F.A.; Espinosa, C.; Esteban, M.A.; Cuesta, A.; Girondot, M.; Romero, D. Molecular oxidative stress markers in olive ridley turtles (Lepidochelys olivacea) and their relation to metal concentrations in wild populations. Environ. Pollut. 2018, 233, 156–167. [Google Scholar] [CrossRef]
- Do You Know the 7 Sea Turtle Species? Available online: https://twitter.com/manatipr/status/1272943922348732416 (accessed on 24 December 2022).
- Speer, R.M.; Wisea, S.S.; Croom-Pereza, T.J.; Aboueissab, A.M.; Brasa, M.M.; Barandiaran, M.; Bermudezd, E.; Wise Sr, J.P.A. comparison of particulate hexavalent chromium cytotoxicity and genotoxicity in human and leatherback sea turtle lung cells from a one environmental health perspective. Toxicol. Appl. Pharm. 2019, 376, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Engwa, A.G.; Ferdinand, P.U.; Nwalo, F.N.; Unachukwu, M.N. Mechanism and health effects of heavy metal toxicity in humans. In Poisoning in the Modern World; Karcioglu, O., Arslan, B., Eds.; IntechOpen: London, UK, 2019; pp. 1–23. [Google Scholar]
- Rand, G.M. Fundamentals of Aquatic Toxicology: Effects, Environmental Fate, and Risk Assessment; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Walker, C.H.; Hopkin, S.P.; Sibly, R.M.; Peakall, D.B. Principles of Ecotoxicology; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Godley, B.J.; Thompson, D.R.; Furness, R.W. Do heavy metal concentrations pose a threat to marine turtles from the Mediterranean Sea? Mar. Poll. Bull. 1999, 38, 497–502. [Google Scholar] [CrossRef]
- Jerez, S.; Motas, M.; Cánovas, R.Á.; Talavera, J.; Almela, R.M.; Del Río, A.B. Accumulation and tissue distribution of heavy metals and essential elements in loggerhead turtles (Caretta caretta) from Spanish Mediterranean coastline of Murcia. Chemosphere 2010, 78, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Saeki, K.; Sakakibara, H.; Sakai, H.; Kunito, T.; Shinsuke, T. Arsenic accumulation in three species of sea turtles. Biometals 2000, 13, 241–250. [Google Scholar] [CrossRef]
- Gardner, S.C.; Fitzgerald, S.L.; Vargas, B.A.; Rodríguez, L.M. Heavy metal accumulation in four species of sea turtles from the Baja California peninsula, Mexico. Biometals 2006, 19, 91–99. [Google Scholar] [CrossRef]
- García-Fernández, A.J.; Gómez-Ramírez, P.; Martínez-López, E.; Hernández-García, A.; María-Mojica, P.; Romero, D.; Bellido, J.J. Heavy metals in tissues from loggerhead turtles (Caretta caretta) from the southwestern Mediterranean (Spain). Ecotox. Environ. Saf. 2009, 72, 557–563. [Google Scholar] [CrossRef]
- Day, R.D.; Christopher, S.J.; Becker, P.R.; Whitaker, D.W. Monitoring Mercury in the Loggerhead Sea Turtle, Caretta caretta. Environ. Sci. Technol. 2005, 39, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Day, R.D.; Segars, A.L.; Arendt, M.D.; Lee, A.M.; Peden-Adams, M.M. Relationship of Blood Mercury Levels to Health Parameters in the Loggerhead Sea Turtle (Caretta caretta). Environ. Health Persp. 2007, 115, 1421–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, R.D.; Keller, J.M.; Harms, C.A.; Segars, A.L.; Cluse, W.M.; Godfrey, M.H.; Lee, A.M.; Peden-Adams, M.M.; Thorvalson, K.; Dodd, M.; et al. Comparison of Mercury Burdens in Chronically Debilitated and Healthy Loggerhead Sea Turtles (Caretta caretta). J. Wildl. Dis. 2010, 46, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Aymak, C.; Uçar, A.H.; Ergene, S. Distribution of heavy metals in tissues of stranded Loggerhead Turtles (Caretta caretta) on Kazanlı Beach, Turkey North. J. Zool. 2021, 17, 82–91. [Google Scholar]
- Franzellitti, S.; Locatelli, C.; Gerosa, G.; Vallini, C.; Fabbri, E. Heavy metals in tissues of loggerhead turtles (Caretta caretta) from the northwestern Adriatic Sea. Comparative Biochemistry and Physiology C. Toxic. Pharm. 2004, 138, 187–194. [Google Scholar]
- Maffucci, F.; Caurant, F.; Bustamante, P.; Bentivegna, F. Trace element (Cd, Cu, Hg, Se, Zn) accumulation and tissue distribution in loggerhead turtles (Caretta caretta) from the Western Mediterranean Sea (southern Italy). Chemosphere 2005, 58, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Andreani, G.; Santoro, M.; Cottignoli, S.; Fabbri, M.; Carpenè, E.; Isani, G. Metal distribution and metallothionein in loggerhead (Caretta caretta) and green (Chelonia mydas) sea turtles. Sci. Tot. Environ. 2008, 390, 287–294. [Google Scholar] [CrossRef]
- Storelli, M.M.; Ceci, E.; Marcotrigiano, G.O. Distribution of heavy metal residues in some tissues of Caretta caretta (Linnaeus) specimen beached along the Adriatic Sea (Italy). Bull. Environ. Cont. Tox. 1998, 60, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Caurant, F.; Bustamante, P.; Bordes, M.; Miramand, P. Bioaccumulation of cadmium, copper and zinc in some tissues of three species of marine turtles stranded along the French Atlantic coasts. Mar. Poll. Bull. 1999, 38, 1085–1091. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Ichihashi, H.; Suganuma, H.; Tatsukawa, R. Heavy metal monitoring in sea turtles using eggs. Mar. Poll. Bull. 1995, 30, 347–353. [Google Scholar] [CrossRef]
- Sakai, H.; Saeki, K.; Ichihashi, H.; Suganuma, H.; Tanabe, S.; Tatsukawa, R. Species-specific distribution of heavy metals in tissues and organs of loggerhead turtle (Caretta caretta) and green turtle (Chelonia mydas) from Japanese coastal waters. Mar. Poll. Bull. 2000, 40, 701–709. [Google Scholar] [CrossRef]
- Storelli, M.M.; Marcotrigiano, G.O. Total organic and inorganic as from marine turtles (Caretta caretta) beached along the Italian coast (South Adriatic Sea). Bull. Environ. Contam. Toxicol. 2000, 65, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Kaska, Y.; Furness, R.W. Heavy metals in marine turtle eggs and hatchlings in the Mediterranean. Zool. Midd. East 2001, 24, 127–132. [Google Scholar] [CrossRef]
- Çelik, A.; Kaska, Y.; Bağ, H.; Aureggi, M.; Semiz, G.; Kartal, A.A.; Elçi, L. Heavy metal monitoring around the nesting environment of green sea turtles in Turkey. Water Air Soil Poll. 2006, 169, 67–79. [Google Scholar] [CrossRef]
- Yalçın Özdilek, Ş.; Özdilek, H.G.; Sangün, M.K. Change in physical and chemical composition of green turtle (Chelonia mydas) eggshells during embryonic development. Chelonian Conserv. Biol. 2011, 10, 265–270. [Google Scholar] [CrossRef]
- McKenzie, C.; Godley, B.J.; Furness, R.W.; Wells, D.E. Concentrations and patterns of organochlorine contaminants in marine turtles from Mediterranean and Atlantic waters. Mar. Environ. Res. 1999, 47, 117–135. [Google Scholar] [CrossRef]
- Tomas, J.; Aznar, F.J.; Raga, J.A. Feeding ecology of the loggerhead turtle Caretta caretta in the western Mediterranean. J. Zool. 2001, 255, 525–532. [Google Scholar] [CrossRef]
- Villa, C.A.; Flint, M.; Bell, I.; Hof, C.; Limpus, C.J.; Gaus, C. Trace element reference intervals in the blood of healthy green sea turtles to evaluate exposure of coastal populations. Environ. Poll. 2017, 220, 1465–1476. [Google Scholar] [CrossRef] [Green Version]
- Jian, L.; Guo, R.; Zheng, X.; Shi, H.; Wang, J. Trace elements in green turtle eggshells and coral sand sediments from the Xisha Islands, South China Sea. Mar. Poll. Bull. 2001, 164, 112036. [Google Scholar] [CrossRef]
- Keller, J.M.; Kucklick, J.R.; Stamper, M.A.; Harms, C.A.; McClellan-Green, P.D. Associations between organochlorine contaminant concentrations and clinical health parameters in loggerhead sea turtles from North Carolina, USA. Environ. Health Perspect. 2004, 112, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Storelli, M.M.; Storelli, A.; D’Addabbo, R.; Marano, C.; Bruno, R.; Marcotrigiano, G.O. Trace elements in loggerhead turtles (Caretta caretta) from the eastern Mediterranean Sea: Overview and evaluation. Environ. Poll. 2005, 135, 163–170. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.; Taniguchi, S.; Becker, J.H.; Werneck, M.R.; Montone, R.C. Occurrence of organochlorines in the green sea turtle (Chelonia mydas) on the northern coast of the state of Säo Paulo, Brazil. Mar. Pollut. Bull. 2016, 112, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Esposito, M.; Perugini, M.; Lambiase, S.; Conte, A.; Baldi, L.; Amorena, M. Seasonal trend of PAHs concentrations in farmed mussels from the coastal areas of the Naples, Italy. Bull. Environ. Contam. Toxicol. 2017, 99, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Clukey, K.E.; Lepczyk, C.A.; Balazs, G.H.; Work, T.M.; Li, Q.X.; Bachman, M.J.; Lynch, J.M. Persistent organic pollutants in fat of three species of Pacific pelagic longline caught sea turtles: Accumulation in relation to ingested plastic marine debris. Sci. Total Environ. 2018, 610–611, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Orós, J.; González-Díaz, O.M.; Monagas, P. High levels of polychlorinated biphenyls in tissues of Atlantic turtles stranded in the Canary Islands, Spain. Chemosphere 2009, 74, 473–478. [Google Scholar] [CrossRef]
- Yaghmour, F. Are oil spills a key mortality factor for marine turtles from the eastern coast of the United Arab Emirates? Mar. Poll. Bull. 2019, 149, 110624. [Google Scholar] [CrossRef]
- Lambiase, S.; Serpe, F.P.; Pilia, M.; Fiorito, F.; Iaccarino, D.; Gallo, P.; Esposito, M. Polychlorinated organic pollutants (PCDD/Fs and DL-PCBs) in loggerhead (Caretta caretta) and green (Chelonia mydas) turtles from Central-Southern Tyrrhenian Sea. Chemosphere 2021, 263, 128226. [Google Scholar] [CrossRef] [PubMed]
- Filippos, L.S.; Taniguchi, S.; Baldassin, P.; Pires, T.; Montone, R.C. Persistent organic pollutants in plasma and stable isotopes in red blood cells of Caretta caretta, Chelonia mydas and Lepidochelys olivacea sea turtles that nest in Brazil. Mar. Pollut. Bull. 2021, 167, 112283. [Google Scholar] [CrossRef] [PubMed]
- Stockholm Convention. Stockholm Convention on Persistent Organic Pollutants, as amended in 2009. Châtelaine (Geneva): Secretariat of the Stockholm Convention. Available online: http://chm.pops.int/TheConvention/Overview/TextoftheConvention/tabid/2232/Default.aspx (accessed on 3 January 2016).
- UNEP. Guidelines for the Identification of PCBs and Materials Containing PCBs. First Issue. August 1999. Châtelaine (Geneva): United Nations Environment Programme, UNEP Chemicals, Switzerland. Available online: http://chm.pops.int/Implementation/PCBs/DocumentsPublications/tabid/665/Default.aspx (accessed on 7 July 2016).
- World Health Organization, Geneva. WHO FOOD ADDITIVES SERIES: 71-S1 Prepared by the Eightieth Meeting of the Joint FAO/WHO Expert Committee on Food Additives (JECFA) Safety Evaluation of Certain Food Additives and Contaminants Supplement 1: Non-Dioxin-Like Polychlorinated Biphenyls. 2016. Available online: https://apps.who.int/iris/handle/10665/246225 (accessed on 24 December 2022).
- Habibullah-Al-Mamun, M.; Ahmed, M.K.; Islam, M.S.; Hossain, A.; Tokumura, M.; Masunaga, S. Polychlorinated biphenyls (PCBs) in commonly consumed seafood from the coastal area of Bangladesh: Occurrence, distribution, and human health implications. Environ. Sci. Pollut. Res. Int. 2019, 26, 1355–1369. [Google Scholar] [CrossRef]
- Lazar, B.; Maslov, L.; Herceg Romaníc, S.; Gríâcan, R.; Krauthacker, B.; Holcer, D.; Tvrtkovic, N. Accumulation of organochlorine contaminants in loggerhead sea turtles, Caretta caretta, from the eastern Adriatic Sea. Chemosphere 2011, 82, 121–129. [Google Scholar] [CrossRef]
- Reddy, A.V.B.; Moniruzzaman, M.; Aminabhavi, T.M. Polychlorinated biphenyls (PCBs) in the environment: Recent updates on sampling, pretreatment, cleanup technologies and their analysis. Chem. Eng. J. 2019, 358, 1186–1207. [Google Scholar] [CrossRef]
- Perugini, M.; Visciano, P.; Manera, M.; Turno, G.; Lucisano, A.; Amorena, M. Polycyclic aromatic hydrocarbons in marine organisms from the Gulf of Naples Tyrrhenian Sea. J. Agric. Food Chem. 2007, 55, 2049–2054. [Google Scholar] [CrossRef] [PubMed]
- Nácher-Mestre, J.; Serrano, R.; Benedito-Palos, L.; Navarro, J.C.; López, F.J.; Kaushik, S.; Pérez-Sánchez, J. Bioaccumulation of polycyclic aromatic hydrocarbons in Gilthead Sea Bream (Sparus aurata L.) exposed to longterm feeding trials with different experimental diets. Arch. Environ. Contam. Toxicol. 2010, 59, 137–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, K.; Perrotta, E. Polycyclic aromatic hydrocarbons (PAHs) in livers of California sea otters. Chemosphere 2008, 71, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Vilca, F.Z.; Rossi, S.; de Olinda, R.A.; Sánchez-Sarmiento, A.M.; Setim Prioste, F.E.; Matushima, E.R.; Tornisielo, V.L. Concentrations of polycyclic aromatic hydrocarbons in liver samples of juvenile green sea turtles from Brazil: Can these compounds play a role in the development of fibropapillomatosis? Mar. Pollut. Bull. 2018, 130, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Nakata, H.; Sakai, Y.; Miyawaki, T.; Takemura, A. Bioaccumulation and toxic potencies of polychlorinated biphenyls and polycyclic aromatic hydrocarbons in tidal flat and coastal ecosystems of the Ariake Sea, Japan. Environ. Sci. Technol. 2003, 37, 3513–3521. [Google Scholar] [CrossRef] [PubMed]
- Meador, J.P.; Stein, J.E.; Reichert, W.L.; Varanasi, U. Bioaccumulation of polycyclic aromatic hydrocarbons by marine organisms. Rev. Environ. Contam. Toxicol. 1995, 143, 79–165. [Google Scholar] [CrossRef]
- Camacho, M.; Luzardo, O.P.; Boada, L.D.; López Jurado, L.F.; Medina, M.; Zumbado, M.; Orós, J. Potential adverse health effects of persistent organic pollutants on sea turtles: Evidence from a cross-sectional study on Cape Verde loggerhead sea turtles. Sci. Total Environ. 2013, 458–460, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Finlayson, K.A.; Limpus, C.J.; van de Merwe, J.P. Temporal changes in chemical contamination of green turtles (Chelonia mydas) foraging in a heavily industrialised seaport. Sci. Total Environ. 2022, 817, 152848. [Google Scholar] [CrossRef] [PubMed]
- Dogruer, G.; Weijs, L.; Tang, J.Y.-M.; Hollert, H.; Kock, M.; Bell, I.; Madden Hof, C.A.; Gaus, C. Effect-based approach for screening of chemical mixtures in whole blood of green turtles from the Great Barrier Reef. Sci. Total Environ. 2018, 612, 321–329. [Google Scholar] [CrossRef]
- Salvarani, P.I.; Morgado, F.; Vieira, L.R.; Rendón-von Osten, J. Organochlorines contaminants in eggs of hawksbill (Eretmochelys imbricata) and green sea turtles (Chelonia mydas) from Mexico coast. Arch. Environ. Contam. Toxicol. 2019, 76, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sarmiento, A.M.; Rossi, S.; Vilca, F.Z.; Thijl Vanstreels, R.E.; Monteiro, S.H.; Vale Luiz Américo, S.; Dos Santos, R.G.; Marigo, G.; Bertozzi, C.P.; Grisi Filho, G.H.H.; et al. Organochlorine pesticides in green sea turtles (Chelonia mydas) with and without fibropapillomatosis caught at three feeding areas off Brazil. Marine Biological Association of the United Kingdom. J. Mar. Biol. Assoc. UK 2017, 97, 215–223. [Google Scholar] [CrossRef]
- Vijayasarathy, S.; Baduel, C.; Hof, C.; Bell, I.; Del Mar Gómez Ramos, M.; Ramos, M.J.G.; Gaus, C. Multi-residue screening of non-polar hazardous chemicals in green turtle blood from different foraging regions of the great barrier reef. Sci. Total Environ. 2019, 652, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Arienzo, M.; Trifuoggi, M.; Espostito, M.; Ferrara, L.; Di Natale, G.; Toscanesi, M. Comparative study of polycyclic aromatic hydrocarbons (PAHs) in salt glands and liver of loggerheads turtles Caretta Caretta stranded along the Mediterranean coast, Southern Italy. Chemosphere 2023, in press. [Google Scholar]
- Zhang, L.B.; Tang, L.; Ying, S.H.; Feng, M.G. Regulative roles of glutathione reductase and four glutaredoxins in glutathione redox, antioxidant activity, and iron homeostasis of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2016, 100, 5907–5917. [Google Scholar] [CrossRef]
- Eid, R.; Boucher, E.; Gharib, N.; Khoury, C.; Arab, N.T.T.; Murray, A.; Young, P.G.; Mandato, C.A.; Greenwood, M.T. Identification of human ferritin, heavy polypeptide 1 (FTH1) and yeast RGI1 (YER067W) as pro-survival sequences that counteract the effects of Bax and copper in Saccharomyces cerevisiae. Exp. Cell Res. 2016, 342, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Morcillo, P.; Esteban, M.; Cuesta, A. Heavy metals produce toxicity, oxidative stress and apoptosis in the marine teleost fish SAF-1 cell line. Chemosphere 2016, 144, 225–233. [Google Scholar] [CrossRef]
- Scudiero, R.; Verderame, M.; Motta, C.M.; Simoniello, P. Unravelling the role of metallothionein on development, reproduction and detoxification in the Wall Lizard Podarcis sicula. Int. J. Mol. Sci. 2017, 18, 1569. [Google Scholar] [CrossRef] [Green Version]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The role of metallothionein in oxidative stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Bridle, A.; Leef, M.; dos Santos, C.N.; Nowak, B. Hepatic expression of metal-related genes and gill histology in sand flathead (Platycephalus bassensis) from a metal contaminated estuary. Mar. Environ. Res. 2017, 131, 80–89. [Google Scholar] [CrossRef]
- Kaska, Y.; Çelik, A.; Bağ, H.; Aureggi, M.; Özel, K.; Elci, A.; Kaska, A.; Elçi, L. Heavy metal monitoring in stranded sea turtles along the Mediterranean coast of Turkey. Fres. Environ. Bull. 2004, 13, 769–776. [Google Scholar]
- Alam, S.K.; Brim, M.S. Organochlorine, PCB, PAH, and metal concentrations in eggs of loggerhead sea turtles (Caretta caretta) from northwest Florida, USA. J. Environ. Sci. Health 2000, B35, 705–724. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.J.; Belisle, A.A.; Sileo, L. Residues of petroleum hydrocarbons in tissues of sea turtles exposed to the Ixtoc I oil spill. J. Wildl. Dis. 1983, 19, 106–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, D.J.; Gay, M.L. Embryotoxic effects of benzo-a-pyrene, chrysene and 7-12-dimethylbenz-a-anthracene in petroleum hydrocarbon mixtures in mallard ducks. J. Toxicol. Environ. Health 1981, 7, 775–787. [Google Scholar] [CrossRef]
- Varanasi, U.; Brown, D.W.; Hom, T.; Burrows, D.G.; Sloan, C.A. Supplemental Information Concerning a Survey of Alaska Subsistence Fish, Marine Mammal, and Invertebrate Samples Collected 1989–1991 for Exposure to Oil Spilled from the Exxon Valdez. Technical Memo; U.S. Department of Energy Office of Scientific and Technical Information: Seattle, WA, USA, 1993. Available online: https://www.osti.gov/biblio/5276248 (accessed on 24 December 2022).
- Gordon, A.N.; Kellyand, W.R.; Cribb, T.H. Lesions caused by cardiovascular flukes (Digenea: Spirorchiidae) in stranded green turtle (Chelonia mydas). Vet. Pathol. 1998, 35, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Finlayson, K.; Leusch, F.; Limpus, C.J.; van der Merwe, J.P. Towards the development of standardised sea turtle primary cell cultures for toxicity testing. Ecotox. Environ. Saf. 2019, 173, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Escher, B.I.; Limpus, C.J.; Gaus, C. Coupling passive sampling with in vitro bioassays and chemical analysis to understand combined effects of bioaccumulative chemicals in blood of marine turtles. Chemosphere 2015, 138, 292–299. [Google Scholar] [CrossRef]
- Keller, J.M.; McClellan-Green, P.D.; Kucklick, J.R.; Keil, D.E.; Peden-Adams, M.M. Effects of organochlorine contaminants on loggerhead sea turtle immunity: Comparison of a correlative field study and in vitro exposure experiments. Environ. Health Perspect. 2006, 114, 70–76. [Google Scholar] [CrossRef]
- Webb, S.J.; Zychowski, G.V.; Bauman, S.W.; Higgins, B.M.; Raudsepp, T.; Gollahon, L.S.; Wooten, K.J.; Cole, J.M.; Godard-Codding, C. Establishment, characterization, and toxicological application of loggerhead sea turtle (Caretta caretta) primary skin fibroblast cell cultures. Environ. Sci. Technol. 2014, 48, 14728–14737. [Google Scholar] [CrossRef]
- Ikonomopoulou, M.P.; Olszowy, H.; Limpus, C.; Francis, R.; Whittier, J. Trace element concentrations in nesting flatback turtles (Natator depressus) from Curtis Island, Queensland, Australia. Mar. Environ. Res. 2011, 71, 10–16. [Google Scholar] [CrossRef]
- Hartung, T. From alternative methods to a new toxicology. Eur. J. Pharm. Biopharm. 2011, 77, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Escher, B.; Leusch, F.D.L. Bioanalytical Tools in Water Quality Assessment, Water Intelligence; IWA Publishing: London, UK, 2012; p. 253. [Google Scholar] [CrossRef]
- Wise Sr., J. P.; Payne, R.; Wise, S.S.; Lacerte, C.; Wise, J.; Gianios, C.; Thompson, W.D.; Perkins, C.; Zheng, T.; Zhu, C.; et al. A global assessment of chromium pollution using sperm whales (Physeter macrocephalus) as an indicator species. Chemosphere 2009, 75, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.D.; Urbano, A.M.; Matoso, E.; Carreira, I.; Almeida, A.; Santos, P.; Botelho, L.; Alves, M.; Monterio, C.; Costa, A.N.; et al. Human bronchial epithelial cells malignantly transformed by hexavalent chromium exhibit and aneuploidy phenotype but no microsatellite instability. Mutat. Res. 2009, 670, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Browning, C.L.; Qin, Q.; Kelly, D.F.; Prakash, R.; Vanoli, F.; Jasin, M.; Wise Sr., J. P. Prolonged particulate hexavalent chromium exposure suppresses homologous recombination repair in human lung cells. Toxicol. Sci. 2016, 153, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.L.; Wise, S.S.; Xie, H.; Zhu, C.; Fukuda, T.; Wise Sr., J.P. Comparative cytotoxicity and genotoxicity of soluble and particulate hexavalent chromium in human and hawksbill sea turtle (Eretmochelys Imbricata) skin cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2015, 178, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Van Den Berg, M.; Birnbaum, L.; Bosveld, A.T.C.; Brunström, B.; Cook, P.; Feeley, M.; Giesy, J.P.; Hanberg, A.; Hasegawa, R.; Kennedy, S.W.; et al. Toxic equivalency factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environ. Health Perspect. 1998, 106, 775–792. [Google Scholar] [CrossRef]
- Aguirre, A.A. Green Turtle Fibropapilloma: An Epidemiologic Perspective. In Research Plan for Marine Turtle Fibropapilloma; Balazs, G.H., Pooley, S.G., Eds.; NOAA-TM-NMFSSWFSC-156; U.S. Department of Commerce, National Oceanographic and Atmospheric Administration, National Marine Fisheries Service: Honolulu, HI, USA, 1991; pp. 107–113. [Google Scholar]
- Bolten, A.B.; Bjorndal, K.A.; Eliazar, P.J.; Gregory, L.F. Seasonal Abundance, Size Distribution, and Blood Biochemical Values of Loggerheads (Caretta caretta) in Port Canaveral Ship Channel, Florida; N. T. M. NMFSSEFSC-353; U.S. Department of Commerce, National Oceanographic and Atmospheric Administration National Marine Fisheries Service Southeast Fisheries Science Center: Miami, FL, USA, 1999; p. 39.
- Aguirre, A.A.; Gardner, S.C.; Marsh, J.C.; Delgado, S.G.; Limpus, C.J.; Nichols, W.J. Hazards associated with the consumption of sea turtle meat and eggs: A review for health care workers and the general public. EcoHealth 2006, 3, 141–153. [Google Scholar] [CrossRef]
- Deem, S.L.; Dierenfeld, E.S.; Sounguet, G.P.; Alleman, A.R.; Cray, C.; Poppenga, R.H.; Norton, T.M.; Karesh, W.B. Bood values in free-ranging nesting leatherback sea turtles (Dermochelys coriacea) on the coast of the Republic of Gabon. J. Zoo Wildl. Med. 2006, 37, 464–471. [Google Scholar] [CrossRef]
- Casal, A.B.; Camacho, M.; Lopez-Jurado, L.F.; Juste, C.; Oros, J. Comparative study of hematologic and plasma biochemical variables in Eastern Atlantic juvenile and adult nesting loggerhead sea turtles (Caretta caretta). Vet. Clin. Pathol. 2009, 38, 213–218. [Google Scholar] [CrossRef]
Figure 4.
Percent frequency of detection of POPs and average levels in the tissues of green sea turtles Chelonia mydas. Adapted from Yaghmour et al. [7]. With the permission of Elsevier.
Figure 4.
Percent frequency of detection of POPs and average levels in the tissues of green sea turtles Chelonia mydas. Adapted from Yaghmour et al. [7]. With the permission of Elsevier.

Figure 5.
Different routes of exposures, elimination, and targets of PPs according to the PBK model [92]. With the permission of Elsevier.
Figure 5.
Different routes of exposures, elimination, and targets of PPs according to the PBK model [92]. With the permission of Elsevier.

Figure 6.
Fibropapillomatosis around the neck in green sea turtles from Hawaii [123].
Figure 6.
Fibropapillomatosis around the neck in green sea turtles from Hawaii [123].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mean ± SD concentrations of heavy metals in sea turtle tissues and pattern of abundancy order over the last decades.
Table 1.
Mean ± SD concentrations of heavy metals in sea turtle tissues and pattern of abundancy order over the last decades.
| Site | N | Tissue | As | Cd | Cr | Cu | Mn | Pb | Zn | Tissue | Abundancy Order | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Turkey | 6 | muscle | 72.8 ± 39.3 | 0.48 ± 0.91 | 0.39 ± 0.49 | 6.50 ± 8.22 | 1.53 ± 0.81 | 2.00 ± 1.30 | 91.09 ± 33.01 | muscle | Zn > As > Cu > Pb > Mn > Cd > Cr | [53] |
| Italy | 10 | muscle | 0.81 ± 0.04 | 2.40 ± 0.24 | 1.35 ± 0.24 | 105 ± 14 | muscle | Zn > Cu > Mn > Cd > Pb | [56] | |||
| Italy | 26 | muscle | 0.20 ± 0.20 | 2.70 ± 1.40 | 107 ± 26.1 | muscle | Zn > Cu > Cd | [55] | ||||
| Spain | 13 | muscle | 40.9 ± 37.3 | 0.08 ± 0.06 | 0.20 ± 0.25 | 113 ± 267 | muscle | Zn > As > Pb > Cd | [46] | |||
| Spain | 20 | muscle | 0.20 ± 0.14 | 5.04 ± 1.93 | 0.26 ± 0.23 | 65.4 ± 28.3 | muscle | Zn > Cu > Pb > Cd | [49] | |||
| Mexico | 5 | muscle | 0.1 ± 0.1 | 0.41 ± 0.11 | 0.84 ± 0.12 | 0.01 ± 0.01 | 31.1 ± 15.4 | muscle | Zn > Mn > Cu > Cd > Pb | [48] | ||
| Turkey | 6 | liver | 37.6 ± 37.6 | 4.59 ± 2.98 | 0.47 ± 0.36 | 27.7 ± 15.8 | 5.08 ± 4.06 | 2.73 ± 0.93 | 83.2 ± 52.1 | Liver | Zn > As > Cu > Mn > Cd > Pb > Cr | [53] |
| Italy | 10 | liver | 2.40 ± 0.40 | 17.5 ± 2.44 | 7.48 ± 1.04 | 0.10 ± 0.08 | 103 ± 14 | Liver | Zn > Cu > Mn > Cd > Pb | [56] | ||
| Italy | 26 | liver | 19.3 ± 34.2 | 37.3 ± 8.7 | 66 ± 42.7 | Liver | Zn > Cu > Cd | [55] | ||||
| Spain | 13 | liver | 12.73 ± 13.03 | 0.81 ± 0.48 | 0.20 ± 0.11 | 30.2 ± 12.2 | Liver | Zn > As > Cd > Pb | [46] | |||
| Spain | 20 | liver | 23.4 ± 53.6 | 21.6 ± 8.03 | 2.75 ± 1.64 | 107.3 ± 82.5 | Liver | Zn > Cd > Cu > Pb | [49] | |||
| Mexico | 5 | liver | 1.75 ± 1.10 | 33.9 ± 11.2 | 1.28 ± 085 | 69.1 ± 10.2 | Liver | Zn > Cu > Cd > Mn > Pb | [48] |
Table 2.
POPs levels in sea turtles worldwide. Concentration for blood was expressed as ng/mL, whereas in all other cases they were expressed in ng/g.
Table 2.
POPs levels in sea turtles worldwide. Concentration for blood was expressed as ng/mL, whereas in all other cases they were expressed in ng/g.
| Location | N | Tissue | POPs | Reference |
|---|---|---|---|---|
| ΣOCPs | ||||
| Spain | 21 | Blood | 0.33 | [33] |
| US | 42 | Blood | 0.20 ± 0.25 | [1] |
| Mexico | 30 | Eggs | 1.81 ± 0.62 | [93] |
| Brazil | 64 | Fat Liver | 104.9 ± 290.7 54.5 ± 99.34 | [94] |
| United Arab Emirates | 22 | Muscle liver | 563.5 ± 295.1 4.53 ± 4.5 | [7] |
| ΣPAHs | ||||
| Brazil | 44 | Liver | 1.48 ± 17.3 | [87] |
| Spain | 21 | Blood | 12.06 | [33] |
| Australia | 86 | Blood | 13 | [95] |
| United Arab Emirates | 22 | Liver Muscle | 15.0 ± 5.0 1.39 ± 0.85 | [7] |
| Italy | 19 | Liver Salt gland | 139 ± 55.0 320± 97.6 | [96] |
| ΣPCBs | ||||
| Spain | 21 | Blood | 0.73 | [33] |
| USA | 42 | Blood | 0.85 | [1] |
| Pelagic Pacific | 6 | Fat | 2.71 | [73] |
| United Arab Emirates | 22 | Muscle liver | ND ND | [7] |
| Italy | 84 | liver | 28.0 ± 52.2 | [2] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arienzo, M. Progress on the Impact of Persistent Pollutants on Marine Turtles: A Review. J. Mar. Sci. Eng. 2023, 11, 266. https://doi.org/10.3390/jmse11020266
AMA Style
Arienzo M. Progress on the Impact of Persistent Pollutants on Marine Turtles: A Review. Journal of Marine Science and Engineering. 2023; 11(2):266. https://doi.org/10.3390/jmse11020266
Chicago/Turabian StyleArienzo, Michele. 2023. "Progress on the Impact of Persistent Pollutants on Marine Turtles: A Review" Journal of Marine Science and Engineering 11, no. 2: 266. https://doi.org/10.3390/jmse11020266
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.