
Artificial Intelligence as a Tool to Study the 3D Skeletal Architecture in Newly Settled Coral Recruits: Insights into the Effects of Ocean Acidification on Coral Biomineralization



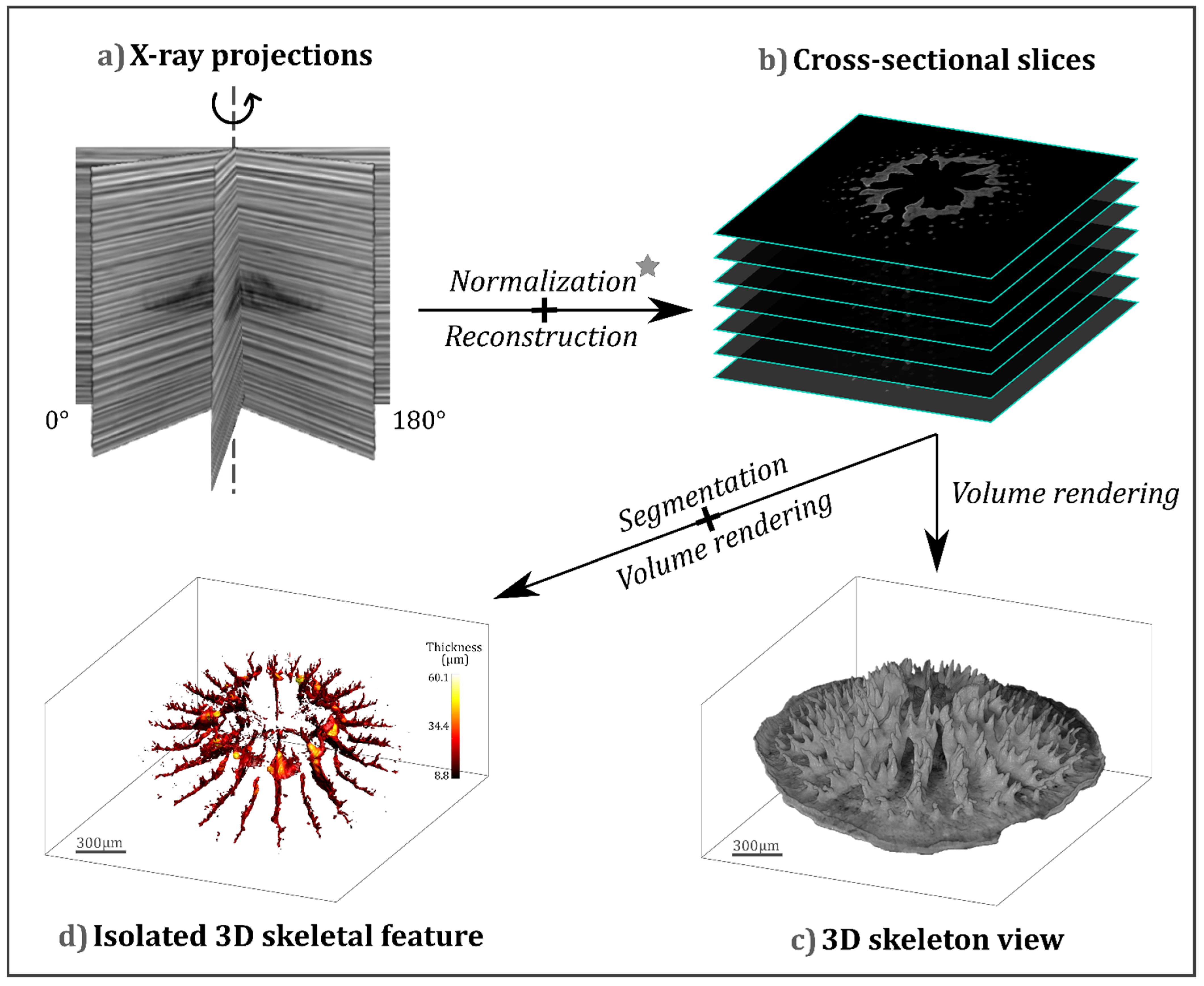{kind=link}
{kind=link}
{kind=link}
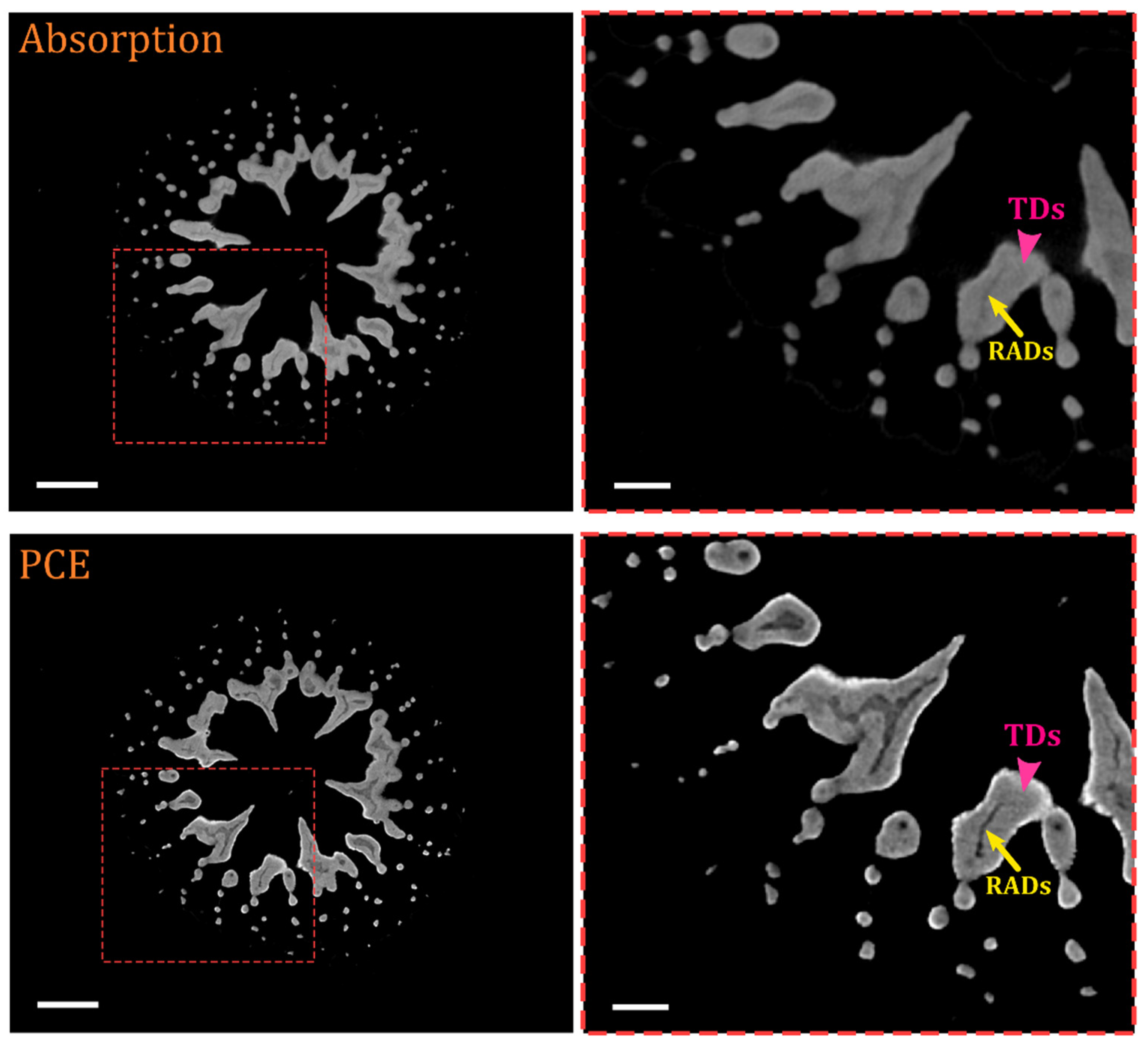{kind=link}
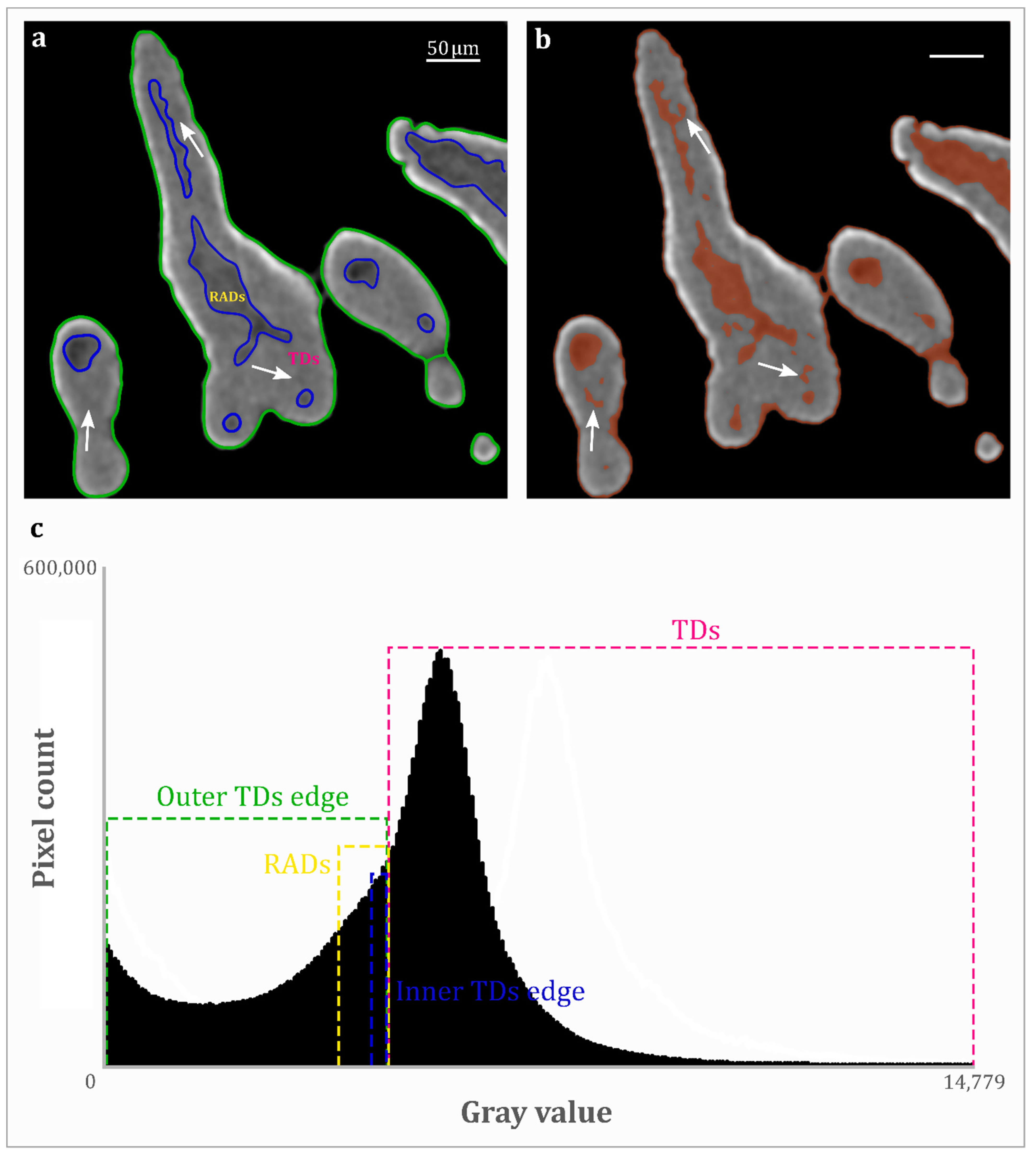{kind=link}
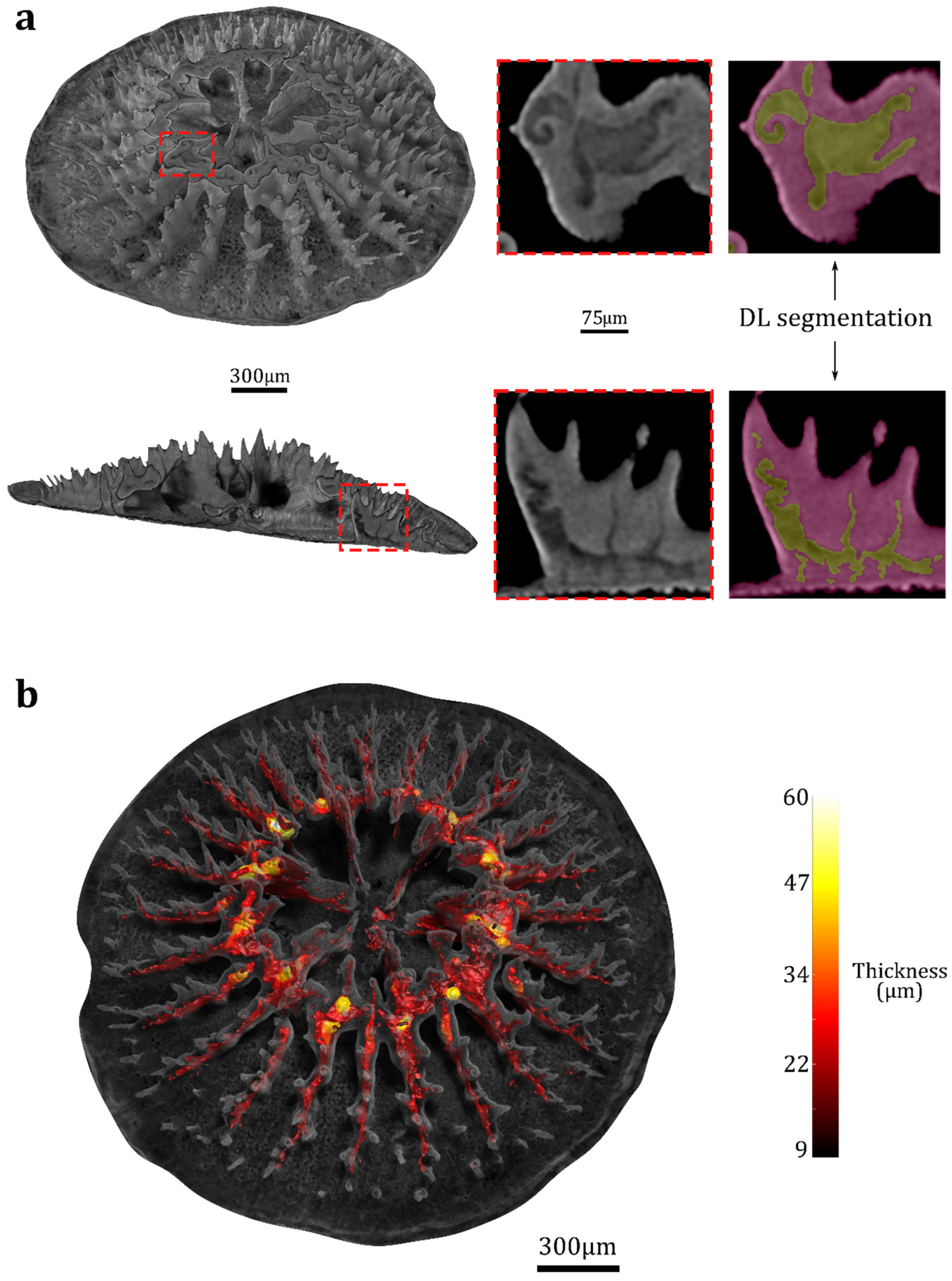{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and OA Experiment
2.2. X-ray µCT: Image Acquisition and Tomographic Reconstruction
2.3. Deep-Learning-Based Image Segmentation
2.4. Performance of the DL-Based Image Segmentation and Evaluation of the RADs/TDs Shape Variability within and between Tomographic Datasets
2.5. Statistical Analysis
3. Results
3.1. Detection and Distinction of Coral Skeletal Structures
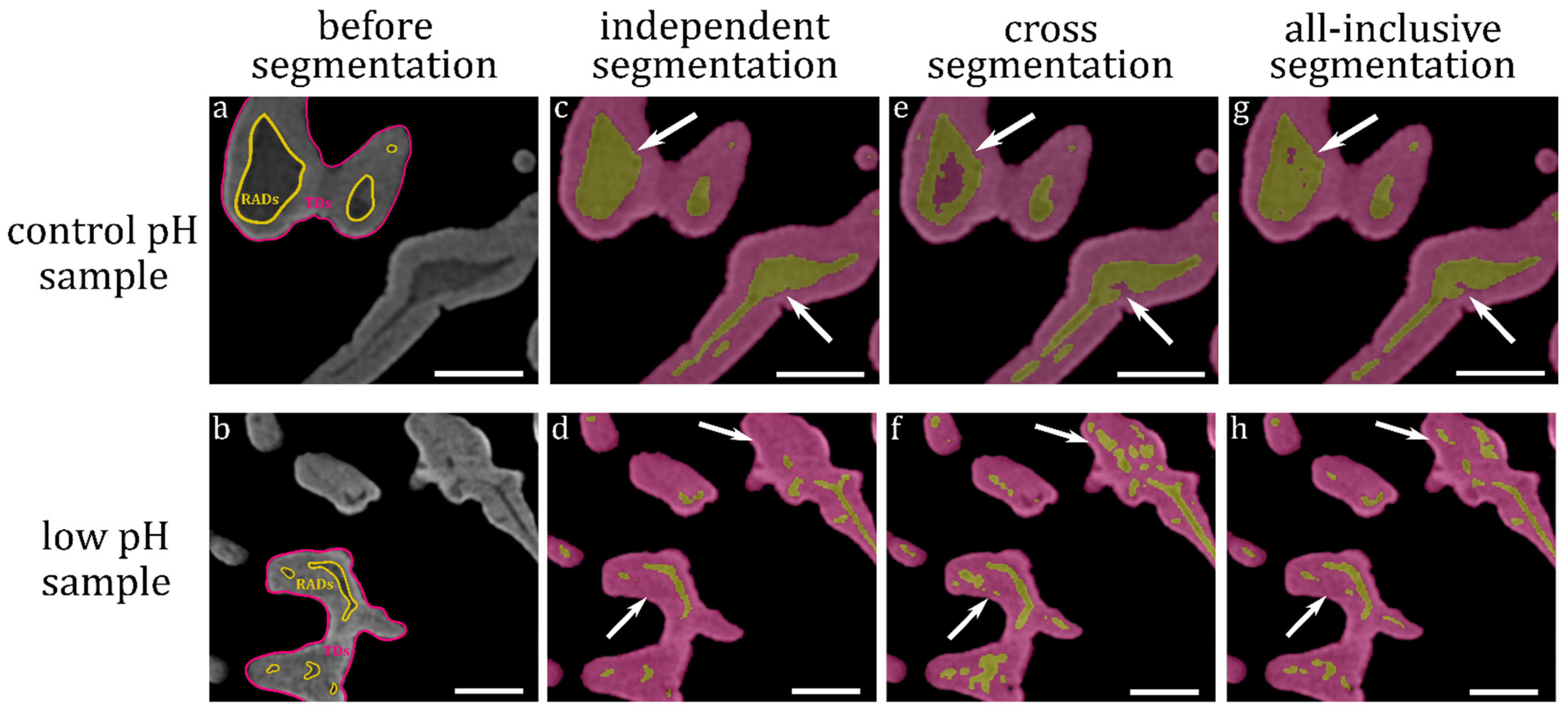
3.2. Segmentation of RADs and TDs
3.3. Evaluation of the DL-Based Segmentation Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brahmi, C.; Domart-Coulon, I.; Rougée, L.; Pyle, D.G.; Stolarski, J.; Mahoney, J.J.; Richmond, R.H.; Ostrander, G.K.; Meibom, A. Pulsed 86Sr-labeling and NanoSIMS imaging to study coral biomineralization at ultra-structural length scales. Coral Reefs 2012, 31, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Cuif, J.-P.; Dauphin, Y. The two-step mode of growth in the scleractinian coral skeletons from the micrometre to the overall scale. J. Struct. Biol. 2005, 150, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Stolarski, J. Three-dimensional micro-and nanostructural characteristics of the scleractinian coral skeleton: A biocalcification proxy. Acta Palaeontol. Pol. 2003, 48, 4. [Google Scholar]
- Meibom, A.; Cuif, J.-P.; Hillion, F.; Constantz, B.R.; Juillet-Leclerc, A.; Dauphin, Y.; Watanabe, T.; Dunbar, R.B. Distribution of magnesium in coral skeleton: MG microdistribution in coral. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef]
- Von Euw, S.; Zhang, Q.; Manichev, V.; Murali, N.; Gross, J.; Feldman, L.C.; Gustafsson, T.; Flach, C.; Mendelsohn, R.; Falkowski, P.G. Biological control of aragonite formation in stony corals. Science 2017, 356, 933–938. [Google Scholar] [CrossRef] [Green Version]
- Mass, T.; Giuffre, A.J.; Sun, C.-Y.; Stifler, C.A.; Frazier, M.J.; Neder, M.; Tamura, N.; Stan, C.V.; Marcus, M.A.; Gilbert, P.U.P.A. Amorphous calcium carbonate particles form coral skeletons. Proc. Natl. Acad. Sci. USA 2017, 114, E7670–E7678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neder, M.; Laissue, P.P.; Akiva, A.; Akkaynak, D.; Albéric, M.; Spaeker, O.; Politi, Y.; Pinkas, I.; Mass, T. Mineral formation in the primary polyps of pocilloporoid corals. Acta Biomater. 2019, 96, 631–645. [Google Scholar] [CrossRef]
- Cuif, J.-P.; Dauphin, Y. Microstructural and physico-chemical characterization of ‘centers of calcification’ in septa of some Recent scleractinian corals. Paläontol. Z. 1998, 72, 257–269. [Google Scholar] [CrossRef]
- Frankowiak, K.; Mazur, M.; Gothmann, A.M.; Stolarski, J. Diagenetic alteration of triassic coral from the aragonite konservat-lagerstatte in alakir cay, turkey: Implications for geochemical measurements. Palaios 2013, 28, 333–342. [Google Scholar] [CrossRef]
- Mass, T.; Drake, J.L.; Peters, E.C.; Jiang, W.; Falkowski, P.G. Immunolocalization of skeletal matrix proteins in tissue and mineral of the coral Stylophora pistillata. Proc. Natl. Acad. Sci. USA 2014, 111, 12728–12733. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, M.; Yasumoto, K.; Iijima, M.; Oaki, Y.; Imai, H. Morphological study of fibrous aragonite in the skeletal framework of a stony coral. CrystEng. Comm. 2021, 23, 3693–3700. [Google Scholar] [CrossRef]
- Benzerara, K.; Menguy, N.; Obst, M.; Stolarski, J.; Mazur, M.; Tylisczak, T.; Brown, G.E.; Meibom, A. Study of the crystallographic architecture of corals at the nanoscale by scanning transmission X-ray microscopy and transmission electron microscopy. Ultramicroscopy 2011, 111, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, C.; Meibom, A.; Smith, D.C.; Stolarski, J.; Auzoux-Bordenave, S.; Nouet, J.; Doumenc, D.; Djediat, C.; Domart-Coulon, I. Skeletal growth, ultrastructure and composition of the azooxanthellate scleractinian coral Balanophyllia regia. Coral Reefs 2010, 29, 175–189. [Google Scholar] [CrossRef]
- Cohen, A.; Holcomb, M. Why Corals Care About Ocean Acidification: Uncovering the Mechanism. Oceanograpgy 2009, 22, 118–127. [Google Scholar] [CrossRef]
- Erez, J.; Reynaud, S.; Silverman, J.; Schneider, K.; Allemand, D. Coral Calcification Under Ocean Acidification and Global Change. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2011; pp. 151–176. ISBN 978-94-007-0113-7. [Google Scholar]
- Kleypas, J.A.; Langdon, C. Coral reefs and changing seawater carbonate chemistry. In Coastal and Estuarine Studies; Phinney, J.T., Hoegh-Guldberg, O., Kleypas, J., Skirving, W., Strong, A., Eds.; American Geophysical Union: Washington, DC, USA, 2006; Volume 61, pp. 73–110. ISBN 978-0-87590-359-0. [Google Scholar]
- Marubini, F.; Ferrier–Pages, C.; Cuif, J. Suppression of skeletal growth in scleractinian corals by decreasing ambient carbonate-ion concentration: A cross-family comparison. Proc. R. Soc. Lond. B 2003, 270, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Mollica, N.R.; Guo, W.; Cohen, A.L.; Huang, K.-F.; Foster, G.L.; Donald, H.K.; Solow, A.R. Ocean acidification affects coral growth by reducing skeletal density. Proc. Natl. Acad. Sci. USA 2018, 115, 1754–1759. [Google Scholar] [CrossRef] [Green Version]
- Scucchia, F.; Malik, A.; Zaslansky, P.; Putnam, H.M.; Mass, T. Combined responses of primary coral polyps and their algal endosymbionts to decreasing seawater pH. Proc. R. Soc. B 2021, 288, 20210328. [Google Scholar] [CrossRef]
- Cohen, A.L.; McCorkle, D.C.; de Putron, S.; Gaetani, G.A.; Rose, K.A. Morphological and compositional changes in the skeletons of new coral recruits reared in acidified seawater: Insights into the biomineralization response to ocean acidification. Geochem. Geophys. Geosyst. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Nothdurft, L.D.; Webb, G.E. Microstructure of Common Reef-Building Coral Genera Acropora, Pocillopora, Goniastrea and Porites: Constraints on Spatial Resolution in Geochemical Sampling. Facies 2007, 53, 1–26. [Google Scholar] [CrossRef]
- Cnudde, V.; Boone, M.N. High-resolution X-ray computed tomography in geosciences: A review of the current technology and applications. Earth-Sci. Rev. 2013, 123, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kyle, J.R.; Ketcham, R.A. Application of high resolution X-ray computed tomography to mineral deposit origin, evaluation, and processing. Ore Geol. Rev. 2015, 65, 821–839. [Google Scholar] [CrossRef]
- Cloetens, P.; Pateyron-Salomé, M.; Buffière, J.Y.; Peix, G.; Baruchel, J.; Peyrin, F.; Schlenker, M. Observation of microstructure and damage in materials by phase sensitive radiography and tomography. J. Appl. Phys. 1997, 81, 5878–5886. [Google Scholar] [CrossRef]
- Baruchel, J. X-ray Tomography in Material Science; Hermes Science: Paris, France, 2000; ISBN 978-2-7462-0115-6. [Google Scholar]
- De Albuquerque, V.H.C.; Cortez, P.C.; de Alexandria, A.R.; Tavares, J.M.R.S. A new solution for automatic microstructures analysis from images based on a backpropagation artificial neural network. Nondestruct. Test. Eval. 2008, 23, 273–283. [Google Scholar] [CrossRef]
- De Albuquerque, V.H.C.; Tavares, J.M.R.S.; Durão, L.M.P. Evaluation of Delamination Damage on Composite Plates using an Artificial Neural Network for the Radiographic Image Analysis. J. Compos. Mater. 2010, 44, 1139–1159. [Google Scholar] [CrossRef]
- Iglesias, J.C.Á.; Santos, R.B.M.; Paciornik, S. Deep learning discrimination of quartz and resin in optical microscopy images of minerals. Miner. Eng. 2019, 138, 79–85. [Google Scholar] [CrossRef]
- Buyya, R.; Calheiros, R.N.; Dastjerdi, A.V. (Eds.) Big Data: Principles and Paradigms; Elsevier/Morgan Kaufmann: Cambridge, MA, USA, 2016; ISBN 978-0-12-805394-2. [Google Scholar]
- Chauhan, S.; Rühaak, W.; Anbergen, H.; Kabdenov, A.; Freise, M.; Wille, T.; Sass, I. Phase segmentation of X-ray computer tomography rock images usingmachine learning techniques: An accuracy and performancestudy. Solid Earth 2016, 7, 1125–1139. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, S.; Rühaak, W.; Khan, F.; Enzmann, F.; Mielke, P.; Kersten, M.; Sass, I. Processing of rock core microtomography images: Using seven different machine learning algorithms. Comput. Geosci. 2016, 86, 120–128. [Google Scholar] [CrossRef]
- Varfolomeev, I.; Yakimchuk, I.; Safonov, I. An Application of Deep Neural Networks for Segmentation of Microtomographic Images of Rock Samples. Computers 2019, 8, 72. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lin, C.L.; Miller, J.D. Improved 3D image segmentation for X-ray tomographic analysis of packed particle beds. Miner. Eng. 2015, 83, 185–191. [Google Scholar] [CrossRef]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Roberts, D.C.; Masson-Delmotte, V.; Zhai, P.; Tignor, M.; Poloczanska, E.; Mintenbeck, K.; Alegría, A.; Nicolai, M.; Okem, A.; et al. (Eds.) Special Report on the Ocean and Cryosphere in a Changing Climate; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Goodbody-Gringley, G. Diel Planulation by the Brooding Coral Favia Fragum (Esper, 1797). J. Exp. Mar. Biol. Ecol. 2010, 389, 70–74. [Google Scholar] [CrossRef]
- Goodbody-Gringley, G.; Wong, K.H.; Becker, D.M.; Glennon, K.; de Putron, S.J. Reproductive Ecology and Early Life History Traits of the Brooding Coral, Porites astreoides, from Shallow to Mesophotic Zones. Coral Reefs 2018, 37, 483–494. [Google Scholar] [CrossRef]
- Pierrot, D.; Lewis, E.; Wallace, D.W.R. MS Excel Program Developed for CO2 System Calculations; ORNL/CDIAC-105a; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, US Department of Energy: Oak Ridge, TN, USA, 2006; Volume 10.
- Mehrbach, C.; Culberson, C.H.; Hawley, J.E.; Pytkowicx, R.M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure 1. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Dickson, A.G.; Millero, F.J. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep. Sea Res. Part A Oceanogr. Res. Pap. 1987, 34, 1733–1743. [Google Scholar] [CrossRef]
- Görner, W.; Hentschel, M.P.; Müller, B.R.; Riesemeier, H.; Krumrey, M.; Ulm, G.; Diete, W.; Klein, U.; Frahm, R. BAMline: The first hard X-ray beamline at BESSY II. Nucl. Instrum. Methods Phys. Res. Sect. A 2001, 467–468, 703–706. [Google Scholar] [CrossRef]
- Rack, A.; Zabler, S.; Müller, B.R.; Riesemeier, H.; Weidemann, G.; Lange, A.; Goebbels, J.; Hentschel, M.; Görner, W. High resolution synchrotron-based radiography and tomography using hard X-rays at the BAMline (BESSY II). Nucl. Instrum. Methods Phys. Res. Sect. A 2008, 586, 327–344. [Google Scholar] [CrossRef]
- Zaslansky, P.; Fratzl, P.; Rack, A.; Wu, M.-K.; Wesselink, P.R.; Shemesh, H. Identification of root filling interfaces by microscopy and tomography methods: Microtomography and microscopy observations of root fillings. Int. Endod. J. 2011, 44, 395–401. [Google Scholar] [CrossRef]
- Reznikov, N.; Buss, D.J.; Provencher, B.; McKee, M.D.; Piché, N. Deep learning for 3D imaging and image analysis in biomineralization research. J. Struct. Biol. 2020, 212, 107598. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Makovetsky, R.; Piche, N.; Marsh, M. Dragonfly as a Platform for Easy Image-based Deep Learning Applications. Microsc. Microanal. 2018, 24, 532–533. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y. Applications. In Gpu-Based Parallel Implementation of Swarm Intelligence Algorithms; Elsevier: Amsterdam, The Netherlands, 2016; pp. 167–177. ISBN 978-0-12-809362-7. [Google Scholar]
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net: Convolutional Networks for Biomedical Image Segmentation. In Medical Image Computing and Computer-Assisted Intervention—MICCAI 2015; Navab, N., Hornegger, J., Wells, W.M., Frangi, A.F., Eds.; Lecture Notes in Computer Science; Springer International Publishing: Cham, Switzerland, 2015; Volume 9351, pp. 234–241. ISBN 978-3-319-24573-7. [Google Scholar]
- Hamwood, J.; Alonso-Caneiro, D.; Read, S.A.; Vincent, S.J.; Collins, M.J. Effect of patch size and network architecture on a convolutional neural network approach for automatic segmentation of OCT retinal layers. Biomed. Opt. Express 2018, 9, 3049. [Google Scholar] [CrossRef] [Green Version]
- Sudre, C.H.; Li, W.; Vercauteren, T.; Ourselin, S.; Jorge Cardoso, M. Generalised Dice Overlap as a Deep Learning Loss Function for Highly Unbalanced Segmentations. In Deep Learning in Medical Image Analysis and Multimodal Learning for Clinical Decision Support; Cardoso, M.J., Arbel, T., Carneiro, G., Syeda-Mahmood, T., Tavares, J.M.R.S., Moradi, M., Bradley, A., Greenspan, H., Papa, J.P., Madabhushi, A., et al., Eds.; Lecture Notes in Computer Science; Springer International Publishing: Cham, Switzerland, 2017; Volume 10553, pp. 240–248. ISBN 978-3-319-67557-2. [Google Scholar]
- Khalifa, F.; El-Baz, A.; Gimel’farb, G.; Ouseph, R.; El-Ghar, M.A. Shape-Appearance Guided Level-Set Deformable Model for Image Segmentation. In Proceedings of the 2010 20th International Conference on Pattern Recognition, Istanbul, Turkey, 23–26 August 2010; pp. 4581–4584. [Google Scholar]
- El-Baz, A.; Gimel’farb, G. Robust image segmentation using learned priors. In Proceedings of the 2009 IEEE 12th International Conference on Computer Vision, Kyoto, Japan, 29 September–2 October 2009; pp. 857–864. [Google Scholar]
- Chen, F.; Yu, H.; Hu, R.; Zeng, X. Deep Learning Shape Priors for Object Segmentation. In Proceedings of the 2013 IEEE Conference on Computer Vision and Pattern Recognition, Portland, OR, USA, 23–28 June 2013; pp. 1870–1877. [Google Scholar]
- Renard, F.; Guedria, S.; Palma, N.D.; Vuillerme, N. Variability and reproducibility in deep learning for medical image segmentation. Sci. Rep. 2020, 10, 13724. [Google Scholar] [CrossRef] [PubMed]
- Sharon, E.; Brandt, A.; Basri, R. Segmentation and boundary detection using multiscale intensity measurements. In Proceedings of the 2001 IEEE Computer Society Conference on Computer Vision and Pattern Recognition, Kauai, HI, USA, 8–14 December 2001; Volume 1. [Google Scholar]
- Guntoro, P.I.; Ghorbani, Y.; Koch, P.-H.; Rosenkranz, J. X-ray Microcomputed Tomography (µCT) for Mineral Characterization: A Review of Data Analysis Methods. Minerals 2019, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Campos, G.F.C.; Mastelini, S.M.; Aguiar, G.J.; Mantovani, R.G.; de Melo, L.F.; Barbon, S. Machine learning hyperparameter selection for Contrast Limited Adaptive Histogram Equalization. J. Image Video Proc. 2019, 2019, 59. [Google Scholar] [CrossRef] [Green Version]
- Saleem, A.; Beghdadi, A.; Boashash, B. Image fusion-based contrast enhancement. J. Image Video Proc. 2012, 2012, 10. [Google Scholar] [CrossRef] [Green Version]
- Withers, P.J.; Bouman, C.; Carmignato, S.; Cnudde, V.; Grimaldi, D.; Hagen, C.K.; Maire, E.; Manley, M.; Du Plessis, A.; Stock, S.R. X-ray computed tomography. Nat. Rev. Methods Primers 2021, 1, 18. [Google Scholar] [CrossRef]
- Paganin, D.; Mayo, S.C.; Gureyev, T.E.; Miller, P.R.; Wilkins, S.W. Simultaneous phase and amplitude extraction from a single defocused image of a homogeneous object. J. Microsc. 2002, 206, 33–40. [Google Scholar] [CrossRef]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef]
- Liu, X.; Song, L.; Liu, S.; Zhang, Y. A Review of Deep-Learning-Based Medical Image Segmentation Methods. Sustainability 2021, 13, 1224. [Google Scholar] [CrossRef]
- Hesamian, M.H.; Jia, W.; He, X.; Kennedy, P. Deep Learning Techniques for Medical Image Segmentation: Achievements and Challenges. J. Digit. Imag. 2019, 32, 582–596. [Google Scholar] [CrossRef] [Green Version]
- Kamnitsas, K.; Ledig, C.; Newcombe, V.F.J.; Simpson, J.P.; Kane, A.D.; Menon, D.K.; Rueckert, D.; Glocker, B. Efficient multi-scale 3D CNN with fully connected CRF for accurate brain lesion segmentation. Med. Image Anal. 2017, 36, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Shorten, C.; Khoshgoftaar, T.M. A survey on Image Data Augmentation for Deep Learning. J. Big Data 2019, 6, 60. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scucchia, F.; Sauer, K.; Zaslansky, P.; Mass, T. Artificial Intelligence as a Tool to Study the 3D Skeletal Architecture in Newly Settled Coral Recruits: Insights into the Effects of Ocean Acidification on Coral Biomineralization. J. Mar. Sci. Eng. 2022, 10, 391. https://doi.org/10.3390/jmse10030391
Scucchia F, Sauer K, Zaslansky P, Mass T. Artificial Intelligence as a Tool to Study the 3D Skeletal Architecture in Newly Settled Coral Recruits: Insights into the Effects of Ocean Acidification on Coral Biomineralization. Journal of Marine Science and Engineering. 2022; 10(3):391. https://doi.org/10.3390/jmse10030391
Chicago/Turabian StyleScucchia, Federica, Katrein Sauer, Paul Zaslansky, and Tali Mass. 2022. "Artificial Intelligence as a Tool to Study the 3D Skeletal Architecture in Newly Settled Coral Recruits: Insights into the Effects of Ocean Acidification on Coral Biomineralization" Journal of Marine Science and Engineering 10, no. 3: 391. https://doi.org/10.3390/jmse10030391