
Mate Perception and Gene Networks Regulating the Early Phase of Sex in Pseudo-nitzschia multistriata
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains Collection and Culture Condition
2.2. Genomic DNA and RNA Extraction
2.3. Gene Expression Analysis
2.4. Capillary Assay
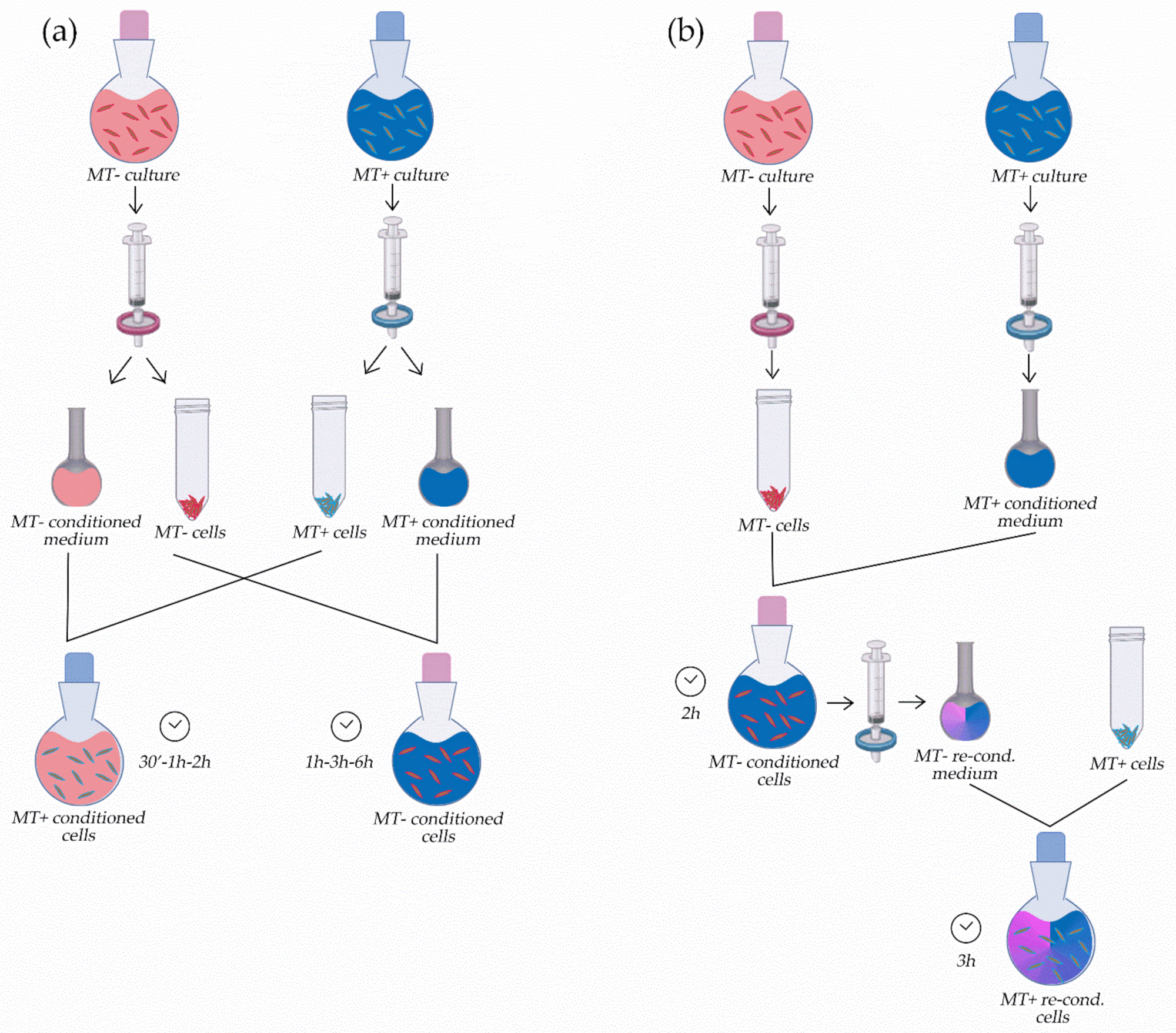
2.5. Experiments with Conditioned Culture Media
2.6. Plasmid Construction
2.7. Biolistic Transformation of P. multistriata
2.8. PCR Analysis on Transformed Cells Overexpressing PmMRP1
3. Results
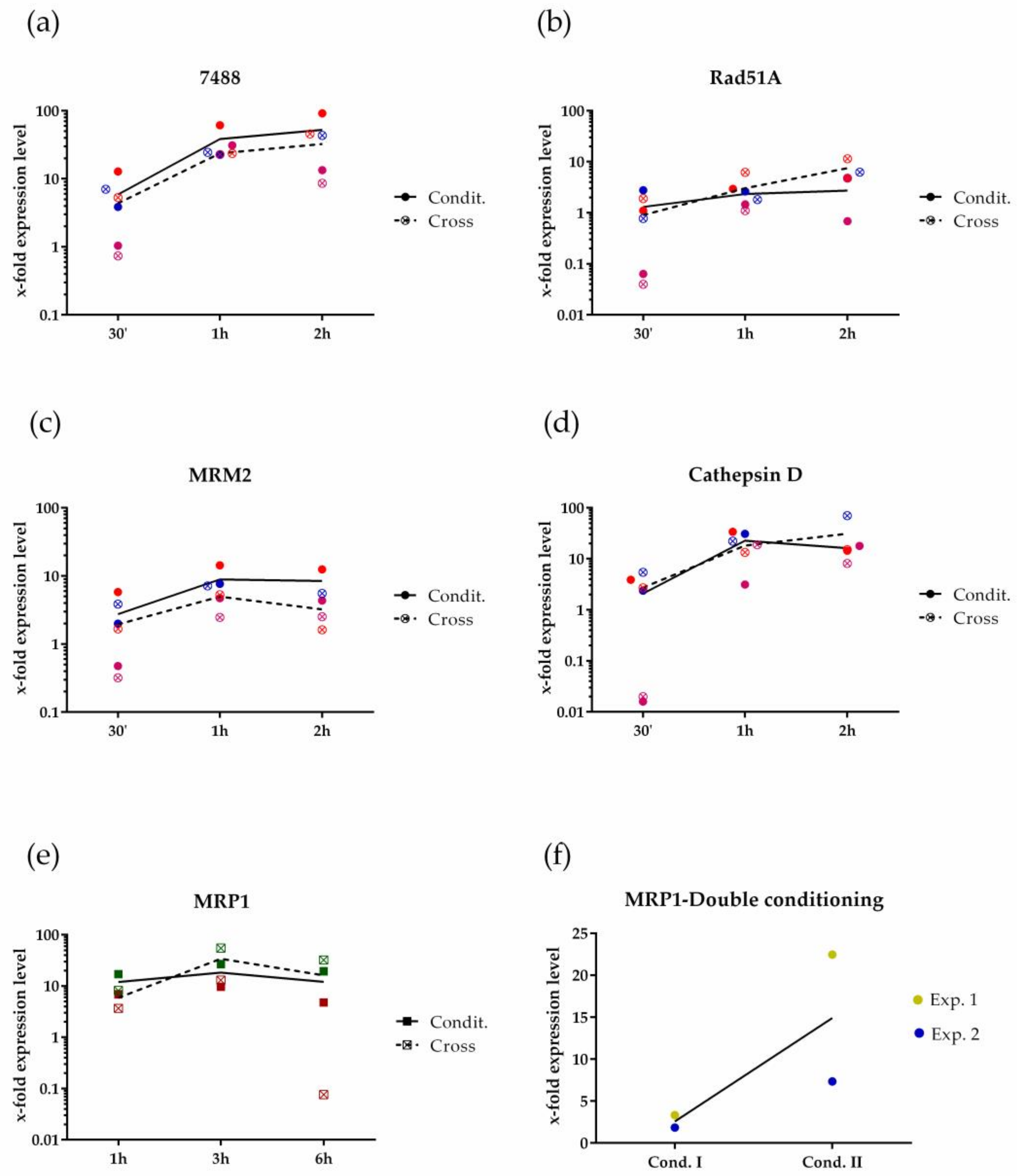
3.1. P. multistriata Molecular Response during Mate Perception
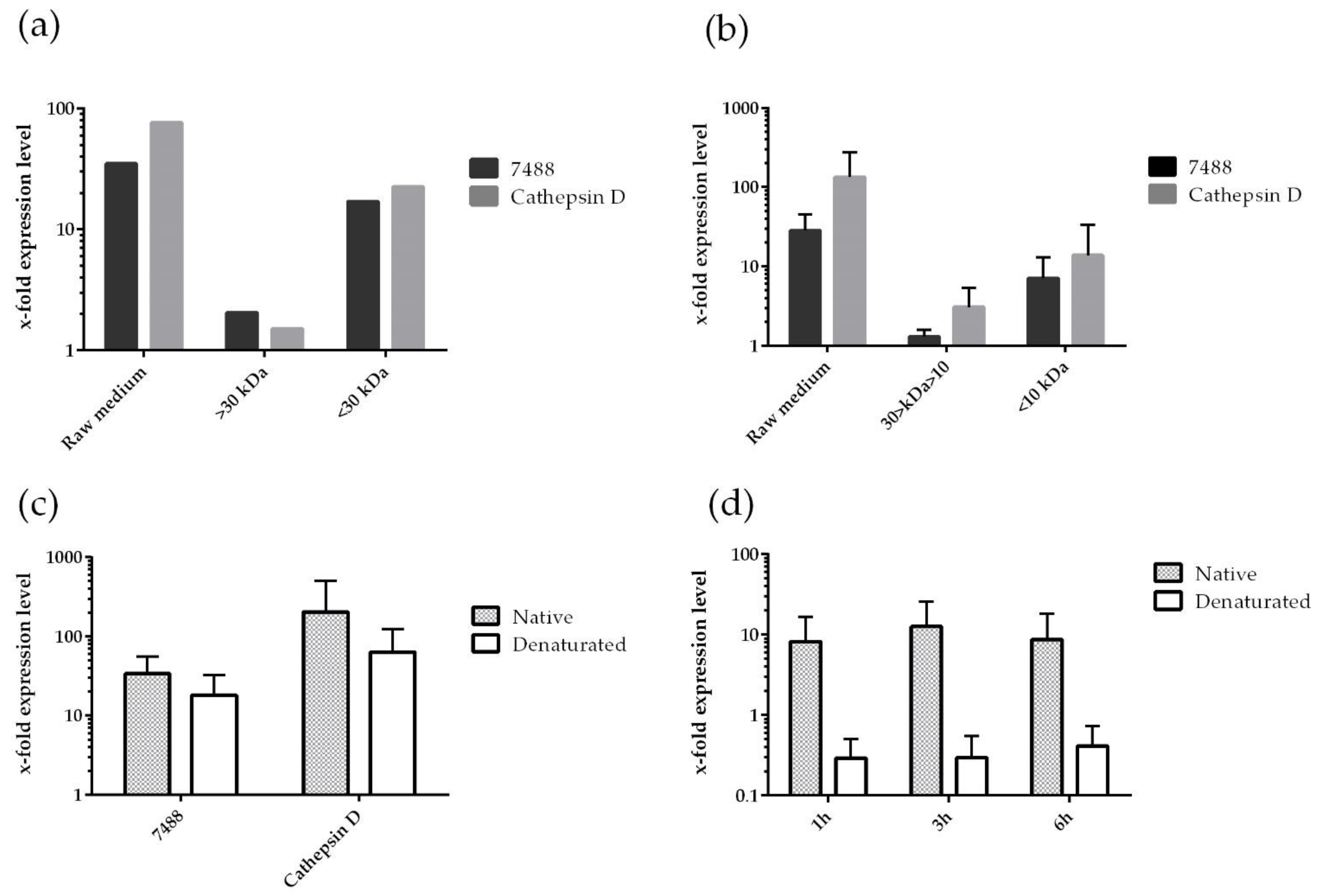
3.2. Profiling of P. multistriata Putative Sex Pheromones Properties
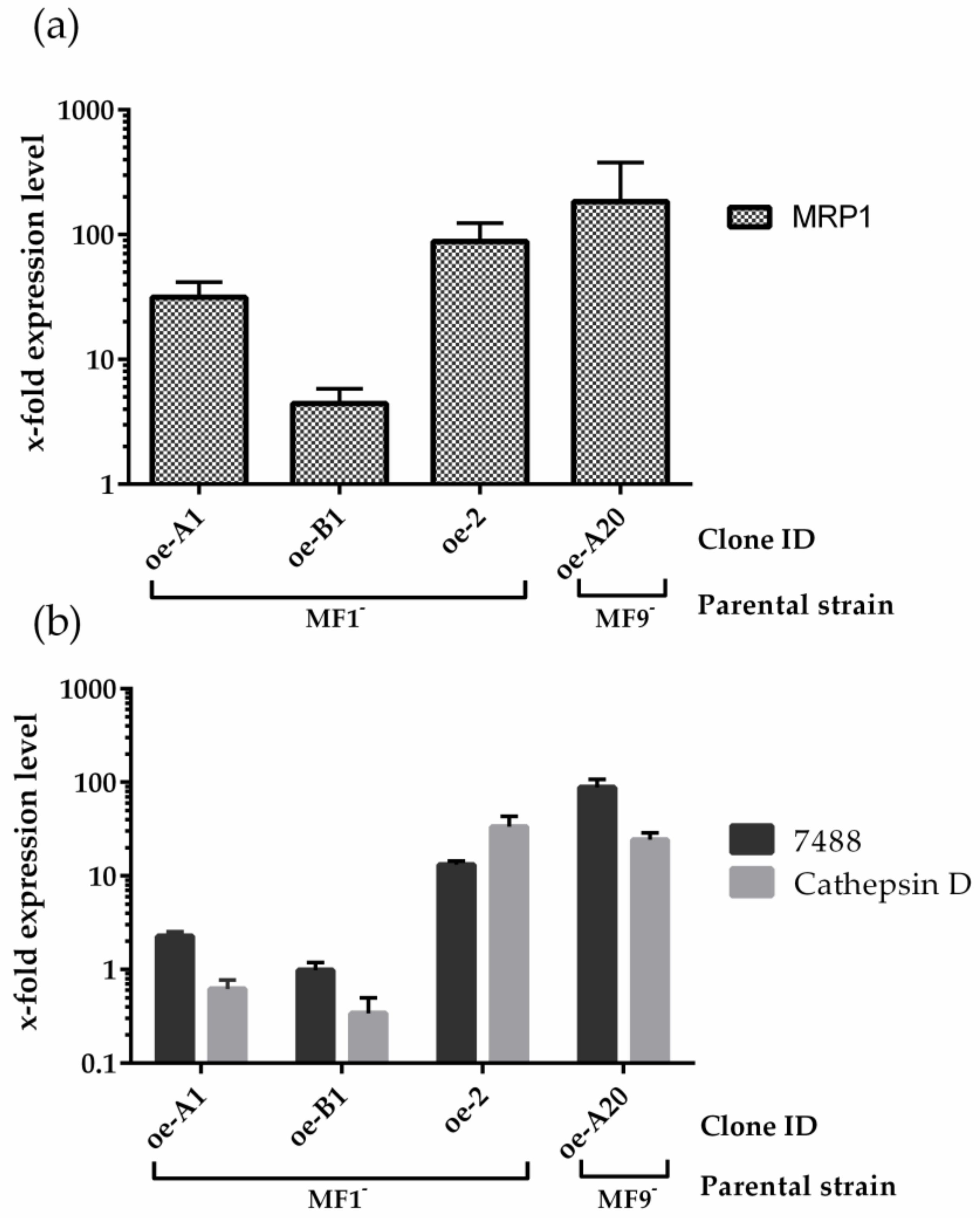
3.3. MRP1 Functional Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armbrust, E.V. The Life of Diatoms in the World’s Oceans. Nature 2009, 459, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N. Biofouling and Antifouling. Nat. Prod. Rep. 2004, 21, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.S.; Poulson-Ellestad, K.L.; Drew Sieg, R.; Poulin, R.X.; Kubanek, J. Chemical Ecology of the Marine Plankton. Nat. Prod. Rep. 2013, 30, 1364–1379. [Google Scholar] [CrossRef] [PubMed]
- Eltanahy, E.; Torky, A. CHAPTER 1: Microalgae as cell factories: Food and feed-grade high-value metabolites. In Microalgal Biotechnology: Recent Advances, Market Potential, and Sustainability; Royal Society of Chemistry: London, UK, 2021; pp. 1–35. [Google Scholar] [CrossRef]
- Balasubramaniam, V.; Gunasegavan, R.D.N.; Mustar, S.; Lee, J.C.; Noh, M.F.M. Isolation of Industrial Important Bioactive Compounds from Microalgae. Molecules 2021, 26, 943. [Google Scholar] [CrossRef] [PubMed]
- Chepurnov, V.A.; Mann, D.G.; Sabbe, K.; Vyverman, W. Experimental Studies on Sexual Reproduction in Diatoms. Int. Rev. Cytol. 2004, 237, 91–154. [Google Scholar] [CrossRef] [PubMed]
- Von Dassow, P.; Montresor, M. Unveiling the Mysteries of Phytoplankton Life Cycles: Patterns and Opportunities behind Complexity. J. Plankton Res. 2011, 33, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Round, F.E.; Frank, E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology & Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; p. 747. [Google Scholar]
- Poulsen, N.C.; Spector, I.; Spurck, T.P.; Schultz, T.F.; Wetherbee, R. Diatom Gliding Is the Result of an Actin-Myosin Motility System. Cell Motil. Cytoskelet. 1999, 44, 23–33. [Google Scholar] [CrossRef]
- Kaczmarska, I.; Poulícková, A.; Sato, S.; Edlund, M.B.; Idei, M.; Watanabe, T.; Mann, D.G. Proposals for a Terminology for Diatom Sexual Reproduction, Auxospores and Resting Stages. Diatom Res. 2013, 28, 263–294. [Google Scholar] [CrossRef]
- Mouget, J.L.; Gastineau, R.; Davidovich, O.; Gaudin, P.; Davidovich, N.A. Light Is a Key Factor in Triggering Sexual Reproduction in the Pennate Diatom Haslea ostrearia. FEMS Microbiol. Ecol. 2009, 69, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Cirri, E.; Vyverman, W.; Pohnert, G. Biofilm Interactions-Bacteria Modulate Sexual Reproduction Success of the Diatom Seminavis Robusta. FEMS Microbiol. Ecol. 2018, 94, fiy161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillard, J.; Frenkel, J.; Devos, V.; Sabbe, K.; Paul, C.; Rempt, M.; Inzé, D.; Pohnert, G.; Vuylsteke, M.; Vyverman, W. Metabolomics Enables the Structure Elucidation of a Diatom Sex Pheromone. Angew. Chem. Int. Ed. Engl. 2013, 52, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.; Vyverman, W.; Pohnert, G. Pheromone Signaling during Sexual Reproduction in Algae. Plant J. 2014, 79, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Bilcke, G.; Ferrante, M.I.; Montresor, M.; De Decker, S.; De Veylder, L.; Vyverman, W. The Molecular Life of Diatoms; Falciatore, A., Mock, T., Eds.; Springer International Publishing: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Sato, S.; Beakes, G.; Idei, M.; Nagumo, T.; Mann, D.G. Novel Sex Cells and Evidence for Sex Pheromones in Diatoms. PLoS ONE 2011, 6, e26923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starr, R.C.; Marnert, F.J.; Jaenicket, L. Chemoattraction of Male Gametes by a Pheromone Produced by Female Gametes of Chlamydomonas. Plant Biol. 1995, 92, 641–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallmann, A.; Godl, K.; Wenzl, S.; Sumper, M. The Highly Efficient Sex-Inducing Pheromone System of Volvox. Trends Microbiol. 1998, 6, 185–189. [Google Scholar] [CrossRef]
- Sekimoto, H. Sexual Reproduction and Sex Determination in Green Algae. J. Plant Res. 2017, 130, 423–431. [Google Scholar] [CrossRef]
- Pohnert, G.; Boland, W. The Oxylipin Chemistry of Attraction and Defense in Brown Algae and Diatoms. Nat. Prod. Rep. 2002, 19, 108–122. [Google Scholar] [CrossRef]
- Kinoshita-Terauchi, N.; Shiba, K.; Umezawa, T.; Matsuda, F.; Motomura, T.; Inaba, K. A Brown Algal Sex Pheromone Reverses the Sign of Phototaxis by CAMP/Ca2+-Dependent Signaling in the Male Gametes of Mutimo Cylindricus (Cutleriaceae). J. Photochem. Photobiol. B Biol. 2019, 192, 113–123. [Google Scholar] [CrossRef]
- Boland, W. The Chemistry of Gamete Attraction: Chemical Structures, Biosynthesis, and (a)Biotic Degradation of Algal Pheromones. Proc. Natl. Acad. Sci. USA 1995, 92, 37. [Google Scholar] [CrossRef] [Green Version]
- Klapper, F.; Audoor, S.; Vyverman, W.; Pohnert, G. Pheromone Mediated Sexual Reproduction of Pennate Diatom Cylindrotheca Closterium. J. Chem. Ecol. 2021, 47, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Moeys, S.; Frenkel, J.; Lembke, C.; Gillard, J.T.F.; Devos, V.; Van den Berge, K.; Bouillon, B.; Huysman, M.J.J.; De Decker, S.; Scharf, J.; et al. A Sex-Inducing Pheromone Triggers Cell Cycle Arrest and Mate Attraction in the Diatom Seminavis Robusta. Sci. Rep. 2016, 6, 19252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilcke, G.; Van den Berge, K.; De Decker, S.; Bonneure, E.; Poulsen, N.; Bulankova, P.; Osuna-Cruz, C.M.; Dickenson, J.; Sabbe, K.; Pohnert, G.; et al. Mating Type Specific Transcriptomic Response to Sex Inducing Pheromone in the Pennate Diatom Seminavis Robusta. ISME J. 2020, 15, 562–576. [Google Scholar] [CrossRef] [PubMed]
- D’Alelio, D.; Amato, A.; Luedeking, A.; Montresor, M. Sexual and Vegetative Phases in the Planktonic Diatom Pseudo-Nitzschia Multistriata. Harmful Algae 2009, 8, 225–232. [Google Scholar] [CrossRef]
- Scalco, E.; Stec, K.; Iudicone, D.; Ferrante, M.I.; Montresor, M. The Dynamics of Sexual Phase in the Marine Diatom Pseudo-Nitzschia Multistriata (Bacillariophyceae). J. Phycol. 2014, 50, 817–828. [Google Scholar] [CrossRef]
- Scalco, E.; Amato, A.; Ferrante, M.I.; Montresor, M. The Sexual Phase of the Diatom Pseudo-Nitzschia Multistriata: Cytological and Time-Lapse Cinematography Characterization. Protoplasma 2015, 253, 1421–1431. [Google Scholar] [CrossRef]
- Sabatino, V.; Russo, M.T.; Patil, S.; d’Ippolito, G.; Fontana, A.; Ferrante, M.I. Establishment of Genetic Transformation in the Sexually Reproducing Diatoms Pseudo-Nitzschia Multistriata and Pseudo-Nitzschia Arenysensis and Inheritance of the Transgene. Mar. Biotechnol. 2015, 17, 452–462. [Google Scholar] [CrossRef]
- Basu, S.; Patil, S.; Mapleson, D.; Russo, M.T.; Vitale, L.; Fevola, C.; Maumus, F.; Casotti, R.; Mock, T.; Caccamo, M.; et al. Finding a Partner in the Ocean: Molecular and Evolutionary Bases of the Response to Sexual Cues in a Planktonic Diatom. New Phytol. 2017, 215, 140–156. [Google Scholar] [CrossRef]
- Annunziata, R.; Mele, B.H.; Marotta, P.; Volpe, M.; Entrambasaguas, L.; Mager, S.; Stec, K.; D’Alcalà, M.R.; Sanges, R.; Finazzi, G.; et al. Trade-off between Sex and Growth in Diatoms: Molecular Mechanisms and Demographic Implications. Sci. Adv. 2022, 8, eabj9466. [Google Scholar] [CrossRef]
- Russo, M.T.; Vitale, L.; Entrambasaguas, L.; Anestis, K.; Fattorini, N.; Romano, F.; Minucci, C.; De Luca, P.; Biffali, E.; Vyverman, W.; et al. MRP3 Is a Sex Determining Gene in the Diatom Pseudo-Nitzschia Multistriata. Nat. Commun. 2018, 9, 5050. [Google Scholar] [CrossRef] [Green Version]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar] [CrossRef]
- Adelfi, M.G.; Borra, M.; Sanges, R.; Montresor, M.; Fontana, A.; Ferrante, M.I. Selection and Validation of Reference Genes for QPCR Analysis in the Pennate Diatoms Pseudo-Nitzschia Multistriata and P. arenysensis. J. Exp. Mar. Bio. Ecol. 2014, 451, 74–81. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative Expression Software Tool (REST) for Group-Wise Comparison and Statistical Analysis of Relative Expression Results in Real-Time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Santin, A.; Russo, M.T.; Marotta, P.; Manfellotto, F.; Ruggiero, A.; Ferrante, M.I. Biolistic Transformation of Pseudo-Nitzschia Multistriata. Available online: https://www.protocols.io/view/biolistic-transformation-of-pseudo-nitzschia-multi-bp2l619qzvqe/v1.dx.doi.org/10.17504/protocols.io.b5jgq4jw (accessed on 24 February 2022).
- Falciatore, A.; Casotti, R.; Leblanc, C.; Abrescia, C.; Bowler, C. Transformation of Nonselectable Reporter Genes in Marine Diatoms. Mar. Biotechnol. 1999, 1, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Moeys, S.; von Dassow, P.; Huysman, M.J.J.; Mapleson, D.; De Veylder, L.; Sanges, R.; Vyverman, W.; Montresor, M.; Ferrante, M.I. Identification of the Meiotic Toolkit in Diatoms and Exploration of Meiosis-Specific SPO11 and RAD51 Homologs in the Sexual Species Pseudo-Nitzschia Multistriata and Seminavis robusta. BMC Genomics 2015, 16, 930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidovich, N.A.; Kaczmarska, I.; Ehrman, J.M. Heterothallic and Homothallic Sexual Reproduction in Tabularia fasciculata (Bacillariophyta). Fottea 2010, 10, 251–266. [Google Scholar] [CrossRef]
- Barkai, N.; Rose, M.D.; Wingreen, N.S. Protease Helps Yeast Find Mating Partners. Nature 1998, 396, 422–423. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Log2 Fold Change (Basu et al., 2017 [31]) | Log2 Fold Change (Annunziata et al., 2022 [32]) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MT+ | MT− | Cross vs. MT+ | Cross vs. MT− | ||||||||
| 2 h | 6 h | 2 h | 6 h | 1 h | 24 h | 5 d | 1 h | 24 h | 5 d | ||
| Cathepsin D | PSNMU-V1.4_AUG-EV-PASAV3_0103000.1 | −0.4 | 0.5 | 5.3 | 7.0 | 9.2 | 8.5 | 8.2 | 5.1 | 4.4 | 4.1 |
| MRM2 | PSNMU-V1.4_AUG-EV-PASAV3_0006960.1 | 0.5 | 0.1 | 1.8 | 2.5 | 9.1 | 8.8 | 8.0 | 1.2 | 1.0 | 0.0 |
| 7488 | PSNMU-V1.4_AUG-EV-PASAV3_0020420.1 | 0.4 | −0.2 | 5.7 | 6.8 | 8.0 | 7.0 | 6.6 | 4.8 | 3.8 | 3.4 |
| Rad51A1 | PSNMU-V1.4_AUG-EV-PASAV3_0056780.1 | 0.8 | 5.8 | 1.8 | 4.8 | 6.1 | 7.8 | 5.2 | 6.2 | 8.0 | 5.3 |
| MRP1 | PSNMU-V1.4_AUG-EV-PASAV3_0024820.1 | −0.1 | 3.7 | −0.1 | −0.9 | 2.5 | 6.0 | 4.4 | 3.7 | 7.4 | 5.7 |
 The color pattern indicates the Log2 Fold Change, ranging from −10 (red) to +10 (green).
The color pattern indicates the Log2 Fold Change, ranging from −10 (red) to +10 (green).Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marotta, P.; Borgonuovo, C.; Santin, A.; Russo, M.T.; Manfellotto, F.; Montresor, M.; De Luca, P.; Ferrante, M.I. Mate Perception and Gene Networks Regulating the Early Phase of Sex in Pseudo-nitzschia multistriata. J. Mar. Sci. Eng. 2022, 10, 1941. https://doi.org/10.3390/jmse10121941
Marotta P, Borgonuovo C, Santin A, Russo MT, Manfellotto F, Montresor M, De Luca P, Ferrante MI. Mate Perception and Gene Networks Regulating the Early Phase of Sex in Pseudo-nitzschia multistriata. Journal of Marine Science and Engineering. 2022; 10(12):1941. https://doi.org/10.3390/jmse10121941
Chicago/Turabian StyleMarotta, Pina, Camilla Borgonuovo, Anna Santin, Monia Teresa Russo, Francesco Manfellotto, Marina Montresor, Pasquale De Luca, and Maria Immacolata Ferrante. 2022. "Mate Perception and Gene Networks Regulating the Early Phase of Sex in Pseudo-nitzschia multistriata" Journal of Marine Science and Engineering 10, no. 12: 1941. https://doi.org/10.3390/jmse10121941