
Absence of Nitrergic Modulation of Starling Response in Haemoglobin-Less Antarctic Fish Chionodraco hamatus

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Isolated and Perfused Working Heart Preparations
2.2.1. Experimental Protocols
2.2.2. Statistical Analyses
2.2.3. Drugs and Chemicals
2.3. NOS Expression and Activation
2.3.1. Immunodetection
2.3.2. Western Blotting and Densitometric Analyses
2.3.3. Statistical Analysis
3. Results
3.1. Isolated Working Heart Preparations
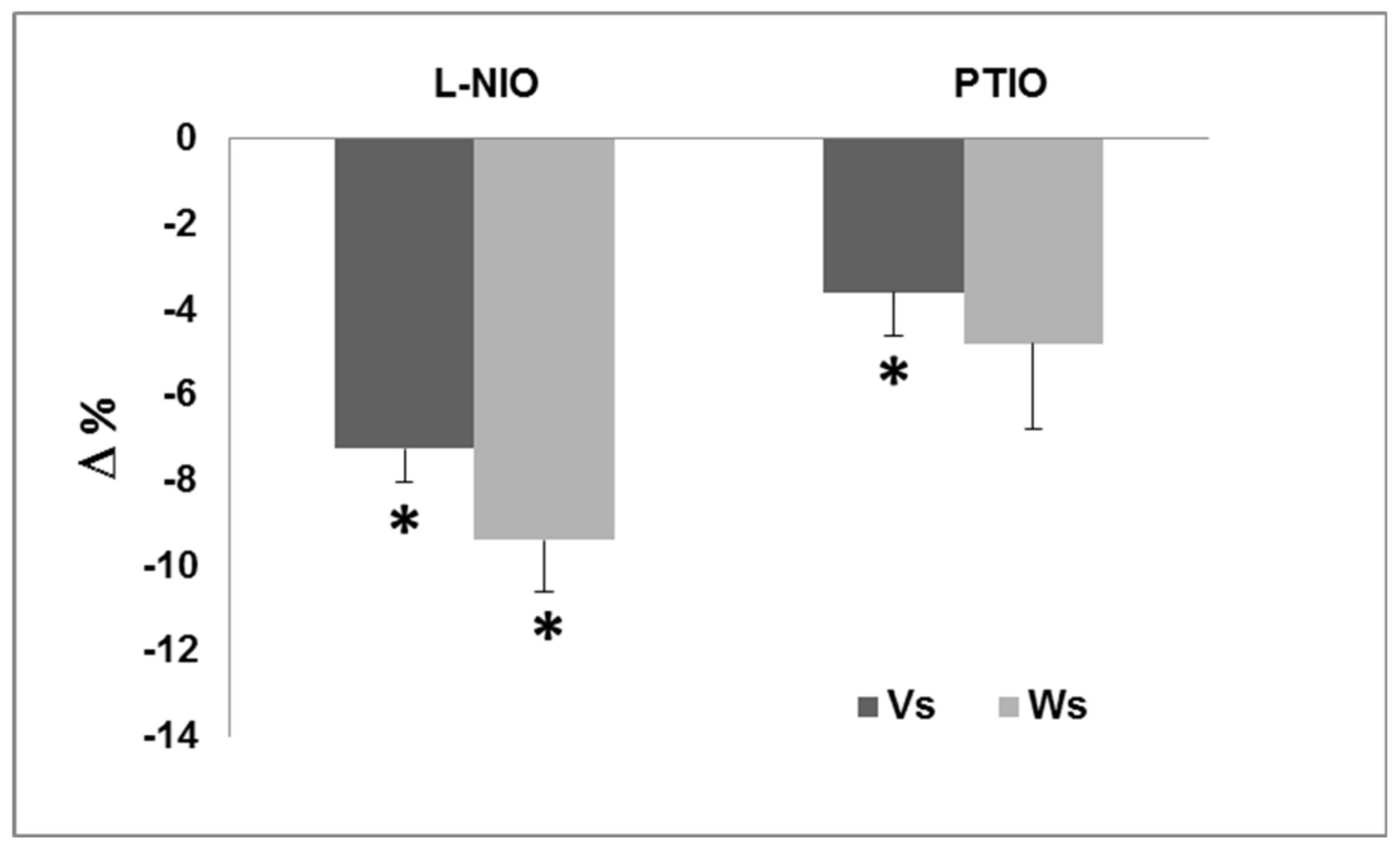
3.1.1. Nitric Oxide and Basal Inotropism
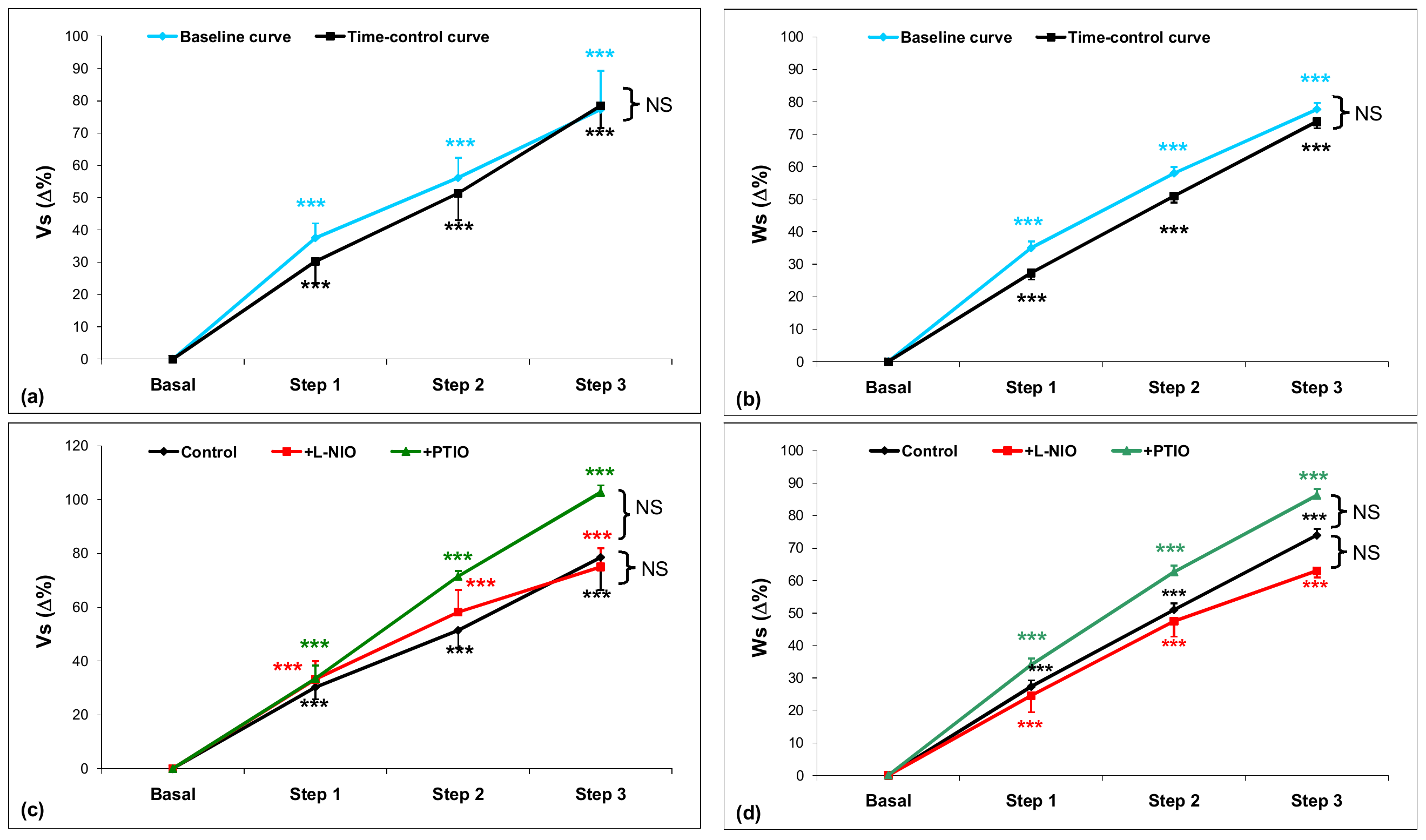
3.1.2. Nitric Oxide and the Frank–Starling Response
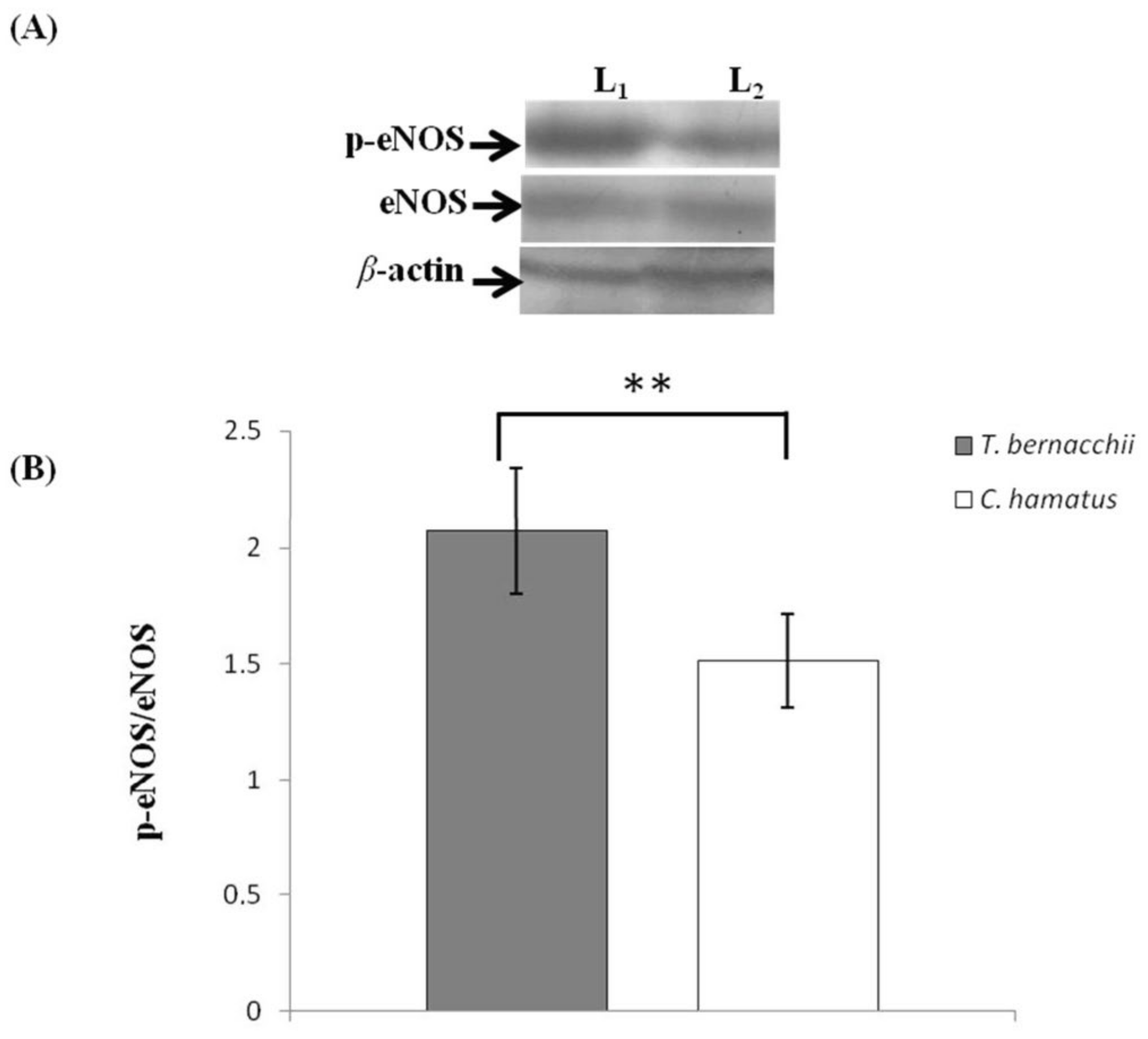
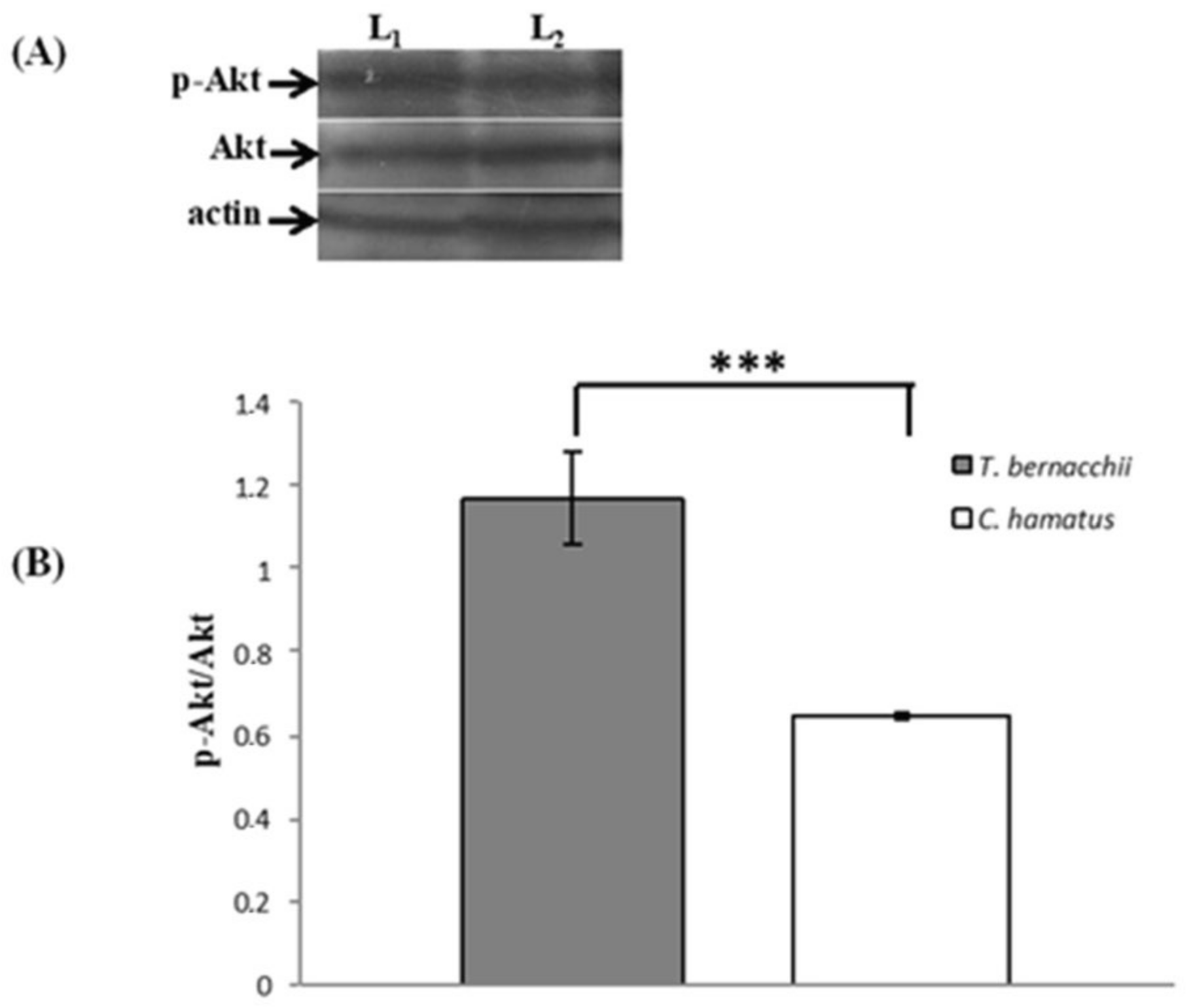
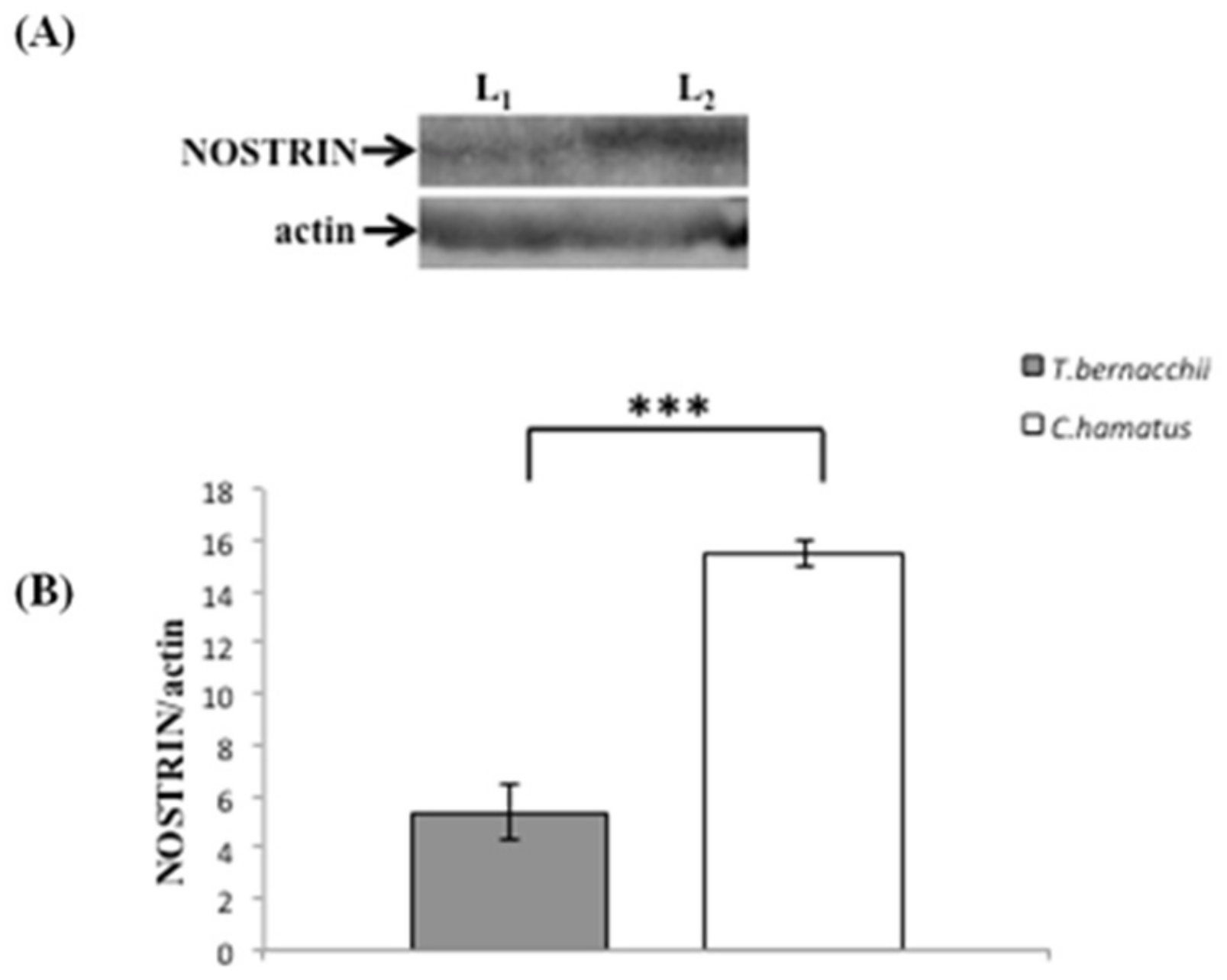
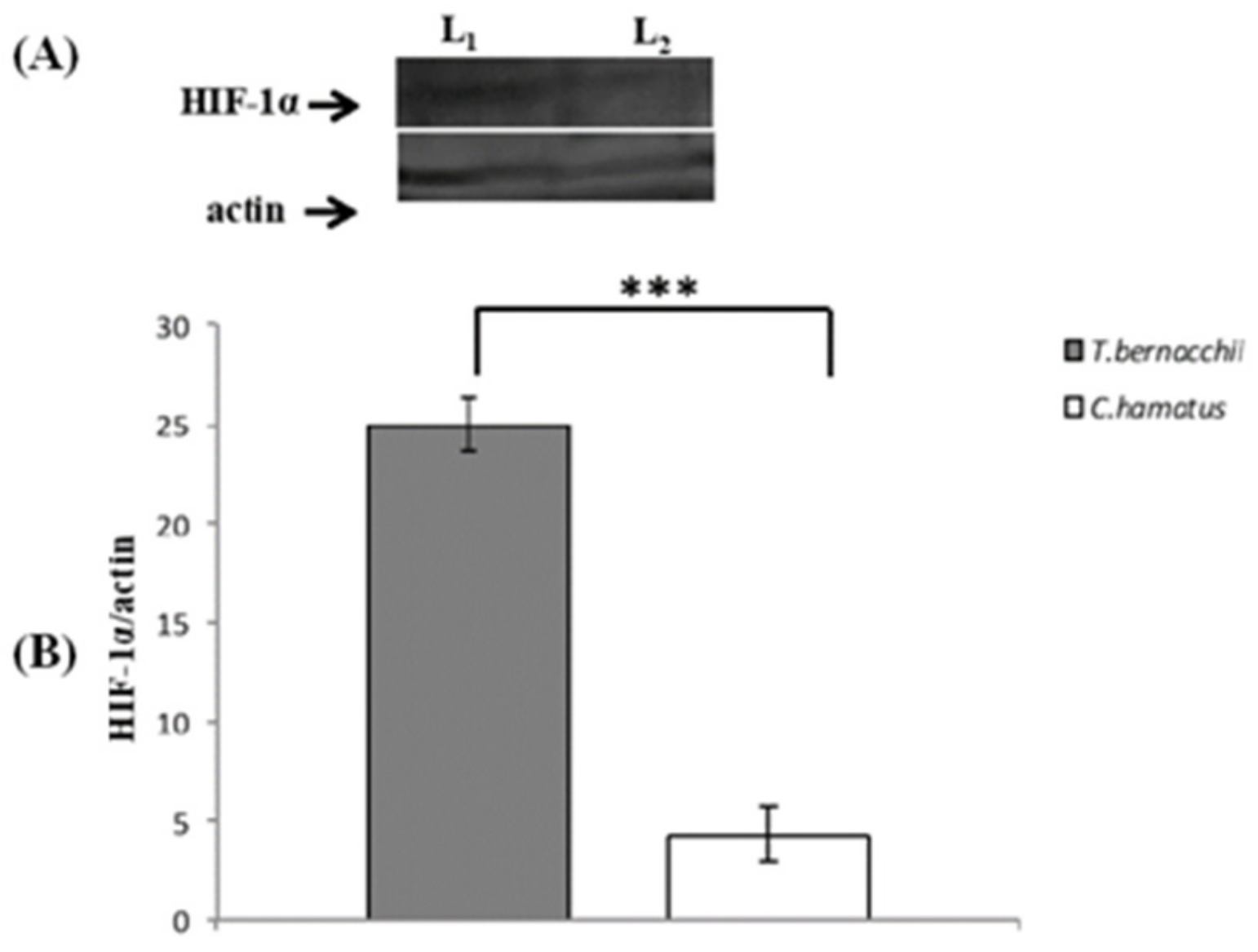
3.2. NOS Expression and Activation
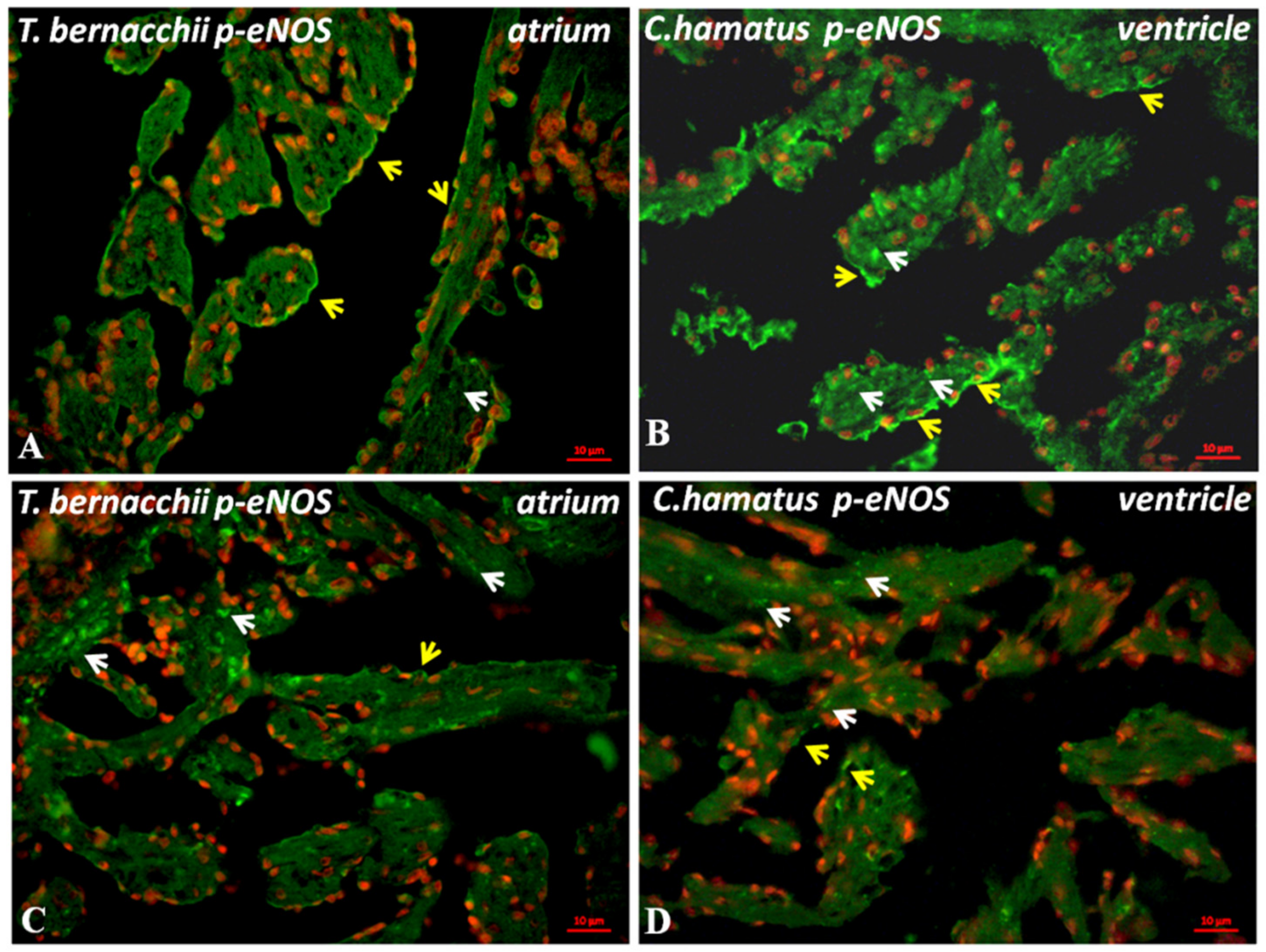
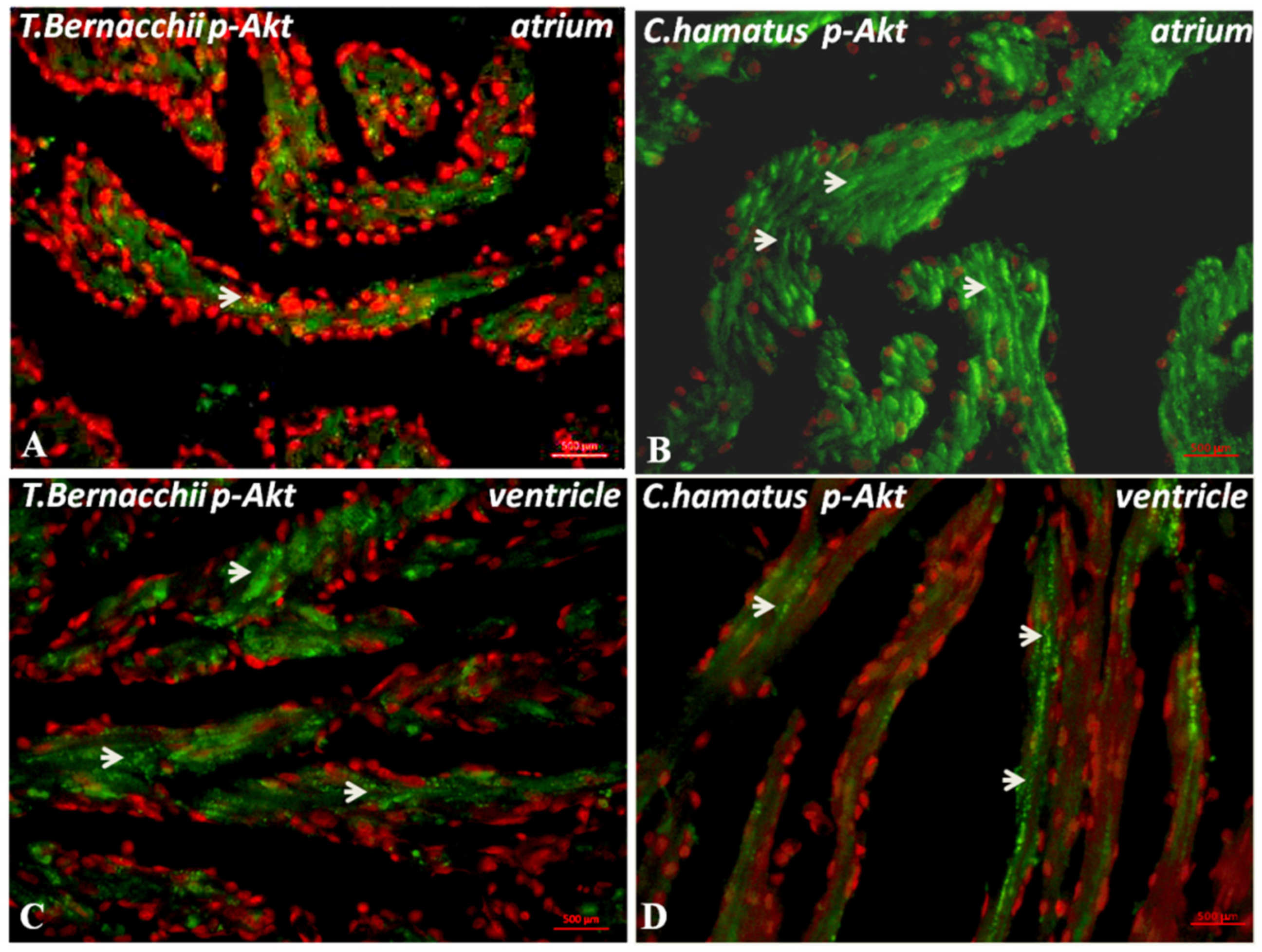
3.2.1. Immunodetection
3.2.2. Western Blotting and Densitometric Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Garofalo, F.; Parisella, M.L.; Amelio, D.; Tota, B.; Imbrogno, S. Phospholamban S-nitrosylation modulates Starling response in fish heart. Proc. R. Soc. B Biol. Sci. 2009, 276, 4043–4052. [Google Scholar] [CrossRef] [Green Version]
- Shiels, H.A.; White, E. The Frank-Starling mechanism in vertebrate cardiac myocytes. J. Exp. Biol. 2008, 211, 2005–2013. [Google Scholar] [CrossRef] [Green Version]
- Gordon, A.M.; Regnier, M.; Homsher, E. Skeletal and cardiac muscle contractile activation: Tropomyosin “rocks and rolls”. News Physiol. Sci. 2001, 16, 49–55. [Google Scholar] [CrossRef]
- Calaghan, S.; White, E. Activation of Na+-H+ exchange and stretch-activated channels underlies the slow inotropic response to stretch in myocytes and muscle from the rat heart. J. Physiol. 2004, 559, 205–214. [Google Scholar] [CrossRef]
- Abraham, D.M.; Davis, R.T.; Warren, C.M.; Mao, L.; Wolska, B.M.; Solaro, R.J.; Rockman, H.A.; Semenza, G.L. β-Arrestin mediates the Frank-Starling mechanism of cardiac contractility. Proc. Natl. Acad. Sci. USA 2016, 113, 14426–14431. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, B.D.; Sagach, V.F.; Shah, A.M. Basal release of nitric oxide augments the Frank-Starling response in the isolated heart. Circulation 1997, 96, 1320–1329. [Google Scholar] [CrossRef]
- Imbrogno, S.; De Iuri, L.; Mazza, R.; Tota, B. Nitric oxide modulates cardiac performance in the heart of Anguilla anguilla. J. Exp. Biol. 2001, 204, 1719–1727. [Google Scholar] [CrossRef]
- Petroff, M.G.V.; Kim, S.H.; Pepe, S.; Dessy, C.; Marbán, E.; Balligand, J.L.; Sollott, S.J. Endogenous nitric oxide mechanisms mediate the stretch dependence of Ca2+ release in cardiomyocytes. Nat. Cell Biol. 2001, 3, 867–873. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Dingle, L.; Hall, R.; Casadei, B. The role of nitric oxide and reactive oxygen species in the positive inotropic response to mechanical stretch in the mammalian myocardium. Biochim. Biophys. Acta-Bioenerg. 2009, 1787, 811–817. [Google Scholar] [CrossRef] [Green Version]
- Devika, N.T.; Jaffar Ali, B.M. Analysing calcium dependent and independent regulation of eNOS in endothelium triggered by extracellular signalling events. Mol. Biosyst. 2013, 9, 2653–2664. [Google Scholar] [CrossRef]
- Dimmeler, S.; Fleming, I.; Fisslthaler, B.; Hermann, C.; Busse, R.; Zeiher, A.M. Activation of nitric oxide synthase in endothelial cells by Akt- dependent phosphorylation. Nature 1999, 399, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Fulton, D.; Gratton, J.P.; McCabe, T.J.; Fontana, J.; Fujio, Y.; Walsh, K.; Franke, T.F.; Papapetropoulos, A.; Sessa, W.C. Regulation of endothelium-derived nitric oxide production by the protein kinase Akt. Nature 1999, 399, 597–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, T.J.; Fulton, D.; Roman, L.J.; Sessa, W.C. Enhanced electron flux and reduced calmodulin dissociation may explain “calcium-independent” eNOS activation by phosphorylation. J. Biol. Chem. 2000, 275, 6123–6128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leucker, T.M.; Bienengraeber, M.; Muravyeva, M.; Baotic, I.; Weihrauch, D.; Brzezinska, A.K.; Warltier, D.C.; Kersten, J.R.; Pratt, P.F. Endothelial-cardiomyocyte crosstalk enhances pharmacological cardioprotection. J. Mol. Cell. Cardiol. 2011, 51, 803–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, S.; Ain, R. Nitric-oxide synthase trafficking inducer is a pleiotropic regulator of endothelial cell function and signaling. J. Biol. Chem. 2017, 292, 6600–6620. [Google Scholar] [CrossRef] [Green Version]
- Michel, T.; Vanhoutte, P.M. Cellular signaling and NO production. Pflugers Arch. Eur. J. Physiol. 2010, 459, 807–816. [Google Scholar] [CrossRef] [Green Version]
- Dudzinski, D.M.; Michel, T. Life history of eNOS: Partners and pathways. Cardiovasc. Res. 2007, 75, 247–260. [Google Scholar] [CrossRef]
- Simmonds, M.J.; Detterich, J.A.; Connes, P. Nitric oxide, vasodilation and the red blood cell. Biorheology 2014, 51, 121–134. [Google Scholar] [CrossRef]
- Muller, B.; Kleschyov, A.L.; Alencar, J.L.; Vanin, A.; Stoclet, J.C. Nitric oxide transport and storage in the cardiovascular system. Ann. N. Y. Acad. Sci. 2002, 962, 131–139. [Google Scholar] [CrossRef]
- Allen, B.W.; Stamler, J.S.; Piantadosi, C.A. Hemoglobin, nitric oxide and molecular mechanisms of hypoxic vasodilation. Trends Mol. Med. 2009, 15, 452–460. [Google Scholar] [CrossRef]
- Gladwin, M.T.; Raat, N.J.H.; Shiva, S.; Dezfulian, C.; Hogg, N.; Kim-Shapiro, D.B.; Patel, R.P. Nitrite as a vascular endocrine nitric oxide reservoir that contributes to hypoxic signaling, cytoprotection, and vasodilation. Am. J. Physiol.-Hear. Circ. Physiol. 2006, 291, H2026–H2035. [Google Scholar] [CrossRef]
- Hendgen-Cotta, U.B.; Kelm, M.; Rassaf, T. A highlight of myoglobin diversity: The nitrite reductase activity during myocardial ischemia-reperfusion. Nitric Oxide-Biol. Chem. 2010, 22, 75–82. [Google Scholar] [CrossRef]
- Gladwin, M.T.; Schechter, A.N. NO Contest: Nitrite Versus S-Nitroso-Hemoglobin. Circ. Res. 2004, 94, 851–855. [Google Scholar] [CrossRef] [Green Version]
- Moylan, T.J.; Sidell, B.D. Concentrations of myoglobin and myoglobin mRNA in heart ventricles from Antarctic fishes. J. Exp. Biol. 2000, 203, 1277–1286. [Google Scholar] [CrossRef]
- Eastman, J.T. Antarctic Fish Biology; Academic Press: Cambridge, MA, USA, 1993; p. 274. [Google Scholar]
- Garofalo, F.; Pellegrino, D.; Amelio, D.; Tota, B. The Antarctic hemoglobinless icefish, fifty five years later: A unique cardiocirculatory interplay of disaptation and phenotypic plasticity. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2009, 154, 10–28. [Google Scholar] [CrossRef]
- Egginton, S.; Sidell, B.D. Thermal acclimation induces adaptive changes in subcellular structure of fish skeletal muscle. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1989, 256. [Google Scholar] [CrossRef]
- O’Brien, K.M.; Sidell, B.D. The interplay among cardiac ultrastructure, metabolism and the expression of oxygen-binding proteins in Antarctic fishes. J. Exp. Biol. 2000, 203, 1287–1297. [Google Scholar] [CrossRef]
- Hardewig, I.; Van Dijk, P.L.M.; Moyes, C.D.; Pörtner, H.O. Temperature-dependent expression of cytochrome-c oxidase in Antarctic and temperate fish. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1999, 277, R508–R516. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.M.; MacCarthy, P.A. Paracrine and autocrine effects of nitric oxide on myocardial function. Pharmacol. Ther. 2000, 86, 49–86. [Google Scholar] [CrossRef]
- Amelio, D.; Garofalo, F.; Pellegrino, D.; Giordano, F.; Tota, B.; Cerra, M.C. Cardiac expression and distribution of nitric oxide synthases in the ventricle of the cold-adapted Antarctic teleosts, the hemoglobinless Chionodraco hamatus and the red-blooded Trematomus bernacchii. Nitric Oxide-Biol. Chem. 2006, 15, 190–198. [Google Scholar] [CrossRef]
- Pellegrino, D.; Palmerini, C.A.; Tota, B. No hemoglobin but NO: The icefish (Chionodraco hamatus) heart as a paradigm. J. Exp. Biol. 2004, 207, 3855–3864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garofalo, F.; Amelio, D.; Cerra, M.C.; Tota, B.; Sidell, B.D.; Pellegrino, D. Morphological and physiological study of the cardiac NOS/NO system in the Antarctic (Hb-/Mb-) icefish Chaenocephalus aceratus and in the red-blooded Trematomus bernacchii. Nitric Oxide-Biol. Chem. 2009, 20, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, D.; Acierno, R.; Tota, B. Control of cardiovascular function in the icefish Chionodraco hamatus: Involvement of serotonin and nitric oxide. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2003, 134. [Google Scholar] [CrossRef]
- Pinsky, D.J.; Patton, S.; Mesaros, S.; Brovkovych, V.; Kubaszewski, E.; Grunfeld, S.; Malinski, T. Mechanical transduction of nitric oxide synthesis in the beating heart. Circ. Res. 1997, 81, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Mebazaa, A.; Wetzel, R.; Cherian, M.; Abraham, M. Comparison between endocardial and great vessel endothelial cells: Morphology, growth, and prostaglandin release. Am. J. Physiol.-Hear. Circ. Physiol. 1995, 268, H250–H259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Casadei, B. Sub-cellular targeting of constitutive NOS in health and disease. J. Mol. Cell. Cardiol. 2012, 52, 341–350. [Google Scholar] [CrossRef]
- Brutsaert, D.L.; Fransen, P.; Andries, L.J.; De Keulenaer, G.W.; Sys, S.U. Cardiac endothelium and myocardial function. Cardiovasc. Res. 1998, 38, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Tota, B.; Amelio, D.; Pellegrino, D.; Ip, Y.K.; Cerra, M.C. NO modulation of myocardial performance in fish hearts. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2005, 142, 164–177. [Google Scholar] [CrossRef]
- Andries, L.J.; Brutsaert, D.L.; Sys, S.U. Nonuniformity of endothelial constitutive nitric oxide synthase distribution in cardiac endothelium. Circ. Res. 1998, 82, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Casadei, B.; Sears, C.E. Nitric-oxide-mediated regulation of cardiac contractility and stretch responses. Prog. Biophys. Mol. Biol. 2003, 82, 67–80. [Google Scholar] [CrossRef]
- Rodriguez-Miguelez, P.; Lima-Cabello, E.; Martínez-Flórez, S.; Almar, M.; Cuevas, M.J.; González-Gallego, J. Hypoxia-inducible factor-1 modulates the expression of vascular endothelial growth factor and endothelial nitric oxide synthase induced by eccentric exercise. J. Appl. Physiol. 2015, 118, 1075–1083. [Google Scholar] [CrossRef] [Green Version]
- Richard, D.E.; Berra, E.; Pouyssegur, J. Nonhypoxic pathway mediates the induction of hypoxia-inducible factor 1α in vascular smooth muscle cells. J. Biol. Chem. 2000, 275, 26765–26771. [Google Scholar] [CrossRef]
- Ke, Q.; Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol. 2006, 70, 1469–1480. [Google Scholar] [CrossRef]
- Tolomeo, A.M.; Carraro, A.; Bakiu, R.; Toppo, S.; Place, S.P.; Ferro, D.; Santovito, G. Peroxiredoxin 6 from the Antarctic emerald rockcod: Molecular characterization of its response to warming. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2016, 186, 59–71. [Google Scholar] [CrossRef]
- Garofalo, F.; Santovito, G.; Amelio, D. Morpho-functional effects of heat stress on the gills of Antarctic T. bernacchii and C. hamatus. Mar. Pollut. Bull. 2019, 141, 194–204. [Google Scholar] [CrossRef]
- Tolomeo, A.M.; Carraro, A.; Bakiu, R.; Toppo, S.; Garofalo, F.; Pellegrino, D.; Gerdol, M.; Ferro, D.; Place, S.P.; Santovito, G. Molecular characterization of novel mitochondrial peroxiredoxins from the Antarctic emerald rockcod and their gene expression in response to environmental warming. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 225. [Google Scholar] [CrossRef]
- Chatzidimitriou, E.; Bisaccia, P.; Corrà, F.; Bonato, M.; Irato, P.; Manuto, L.; Toppo, S.; Bakiu, R.; Santovito, G. Copper/zinc superoxide dismutase from the crocodile icefish Chionodraco hamatus: Antioxidant defense at constant sub-zero temperature. Antioxidants 2020, 9, 325. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. hamatus | |
|---|---|
| Heart rate (beats/min) | 28.6 ± 1.09 |
| Filling pressure (kPa) | 0.11 ± 0.02 |
| Output pressure (kPa) | 1.43 ± 0. 09 |
| Cardiac output (mL/min/kg) | 51.3 ± 3.68 |
| Stroke volume (mL/kg) | 1.8 ± 0.2 |
| Stroke work (mJ/g) | 2.2 ± 0.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amelio, D.; Garofalo, F.; Brunelli, E.; Santovito, G.; Pellegrino, D. Absence of Nitrergic Modulation of Starling Response in Haemoglobin-Less Antarctic Fish Chionodraco hamatus. J. Mar. Sci. Eng. 2022, 10, 1705. https://doi.org/10.3390/jmse10111705
Amelio D, Garofalo F, Brunelli E, Santovito G, Pellegrino D. Absence of Nitrergic Modulation of Starling Response in Haemoglobin-Less Antarctic Fish Chionodraco hamatus. Journal of Marine Science and Engineering. 2022; 10(11):1705. https://doi.org/10.3390/jmse10111705
Chicago/Turabian StyleAmelio, Daniela, Filippo Garofalo, Elvira Brunelli, Gianfranco Santovito, and Daniela Pellegrino. 2022. "Absence of Nitrergic Modulation of Starling Response in Haemoglobin-Less Antarctic Fish Chionodraco hamatus" Journal of Marine Science and Engineering 10, no. 11: 1705. https://doi.org/10.3390/jmse10111705