
Molecular, Morphological and Chemical Diversity of Two New Species of Antarctic Diatoms, Craspedostauros ineffabilis sp. nov. and Craspedostauros zucchellii sp. nov.
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
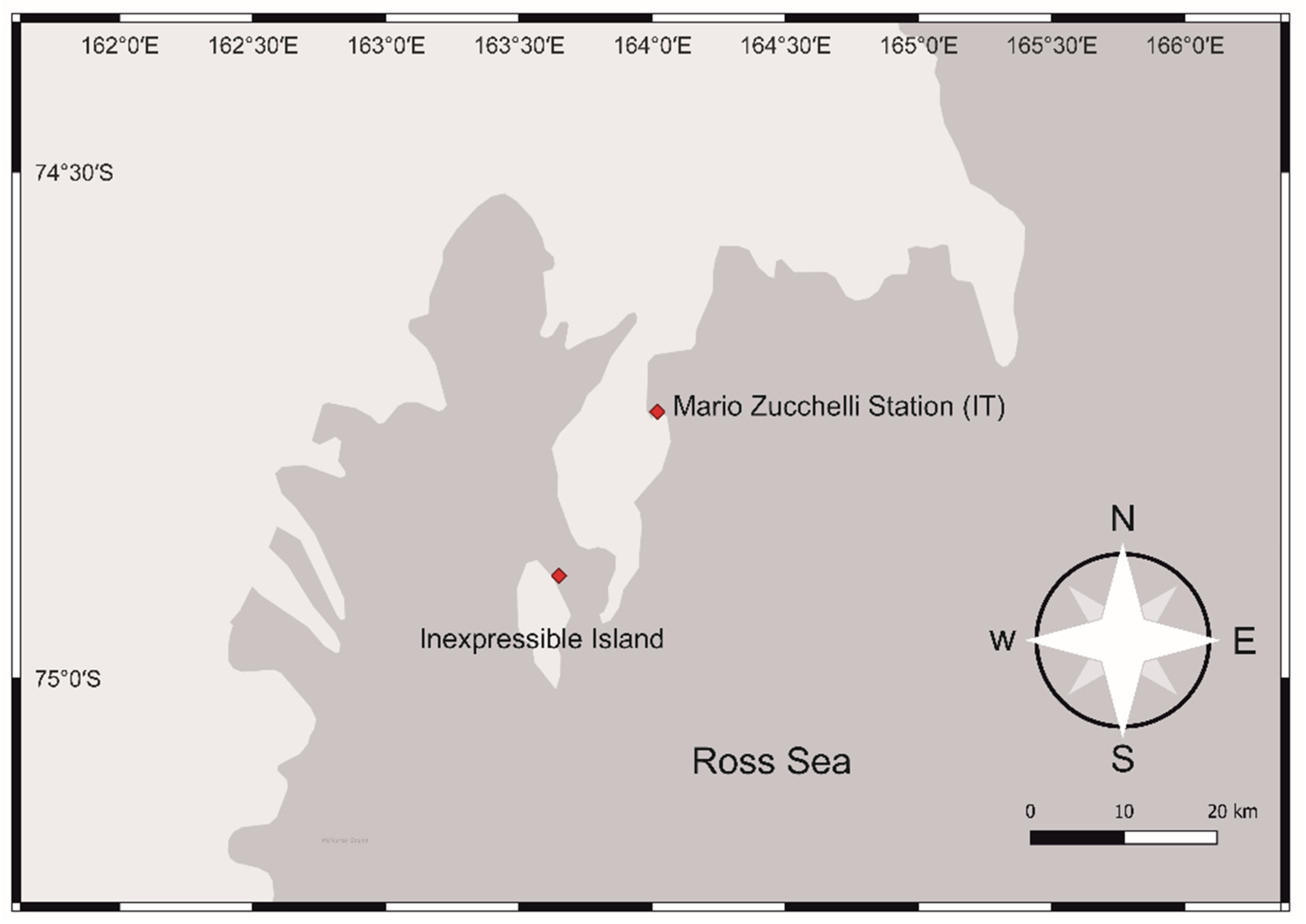
2.1. Isolation and Cultures
2.2. Molecular Analysis
2.2.1. DNA Extraction and Amplification of Selected Molecular Markers
2.2.2. Phylogenetic Analyses
2.3. Morphological Analysis
2.4. Metabolomics
2.4.1. Sample Preparation
2.4.2. UPLC-HR-MS/MS Profiling of Extracts
2.4.3. Metabolomic Data Processing and Statistical Analysis
3. Results
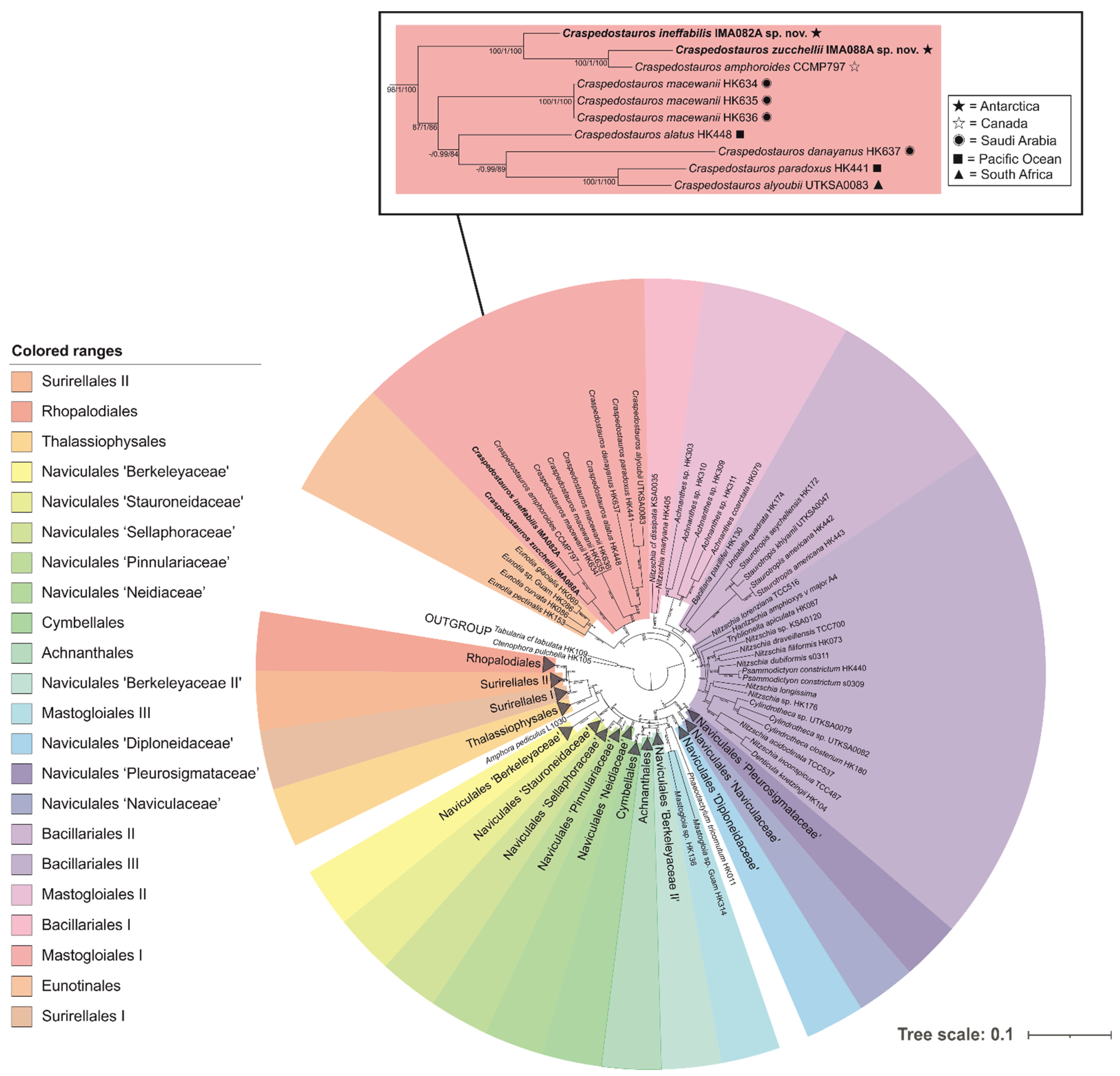
3.1. Phylogenetic Analysis and Hypothesis Testing
3.2. Morphological Observations and Taxonomic Descriptions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Valve Outline | Valve Length (µm) | Valve Width (µm) | Stria Density (in 10 µm) | Areola Size | Internal Central Area | Valve Face–Mantle Junction | Valve Margin at Centre | Average Number of Areolae Pores | Central Lip-Like Silica Flaps | Type Locality | Habitat | Reference | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. neoconstrictus | ±Linear, constricted | 40–110 | 5–7 | ∼25 | Similar | Slight helictoglossae | Indistinct | Expanded | 6 (?) | Unknown | Sussex, England | Marine | [7] |
| C. decipiens | Lanceolate | 20–38 | 3–5 | 20–22 | Variable | Unknown | Distinct | Straight | 10–12 | Unknown | Bosporus | Marine | [7] |
| C. capensis | Lanceolate, constricted | 25–35 | 4.5–5.5 | 19 | Variable | Rectelevatum + Knob | Indistinct | Straight | 5–13 | Rudimentary | Cape Columbine, South Africa | Marine (intertidal) | [7] |
| C. britannicus | Linear to narrow lanceolate | 14–60 | 5–6 | ∼24 | Similar | Helictoglossae | Indistinct | Slightly expanded | 5(+) | Rudimentary | Seascale, Cumbria, UK | Marine (driftwood) | [7] |
| C. australis | Linear | 35–78 | 4–6 | 35 | Similar | Rectelevatum + Knob | Indistinct | Straight | 4 | Rudimentary | Port Phillip Bay, Australia | Marine | [7] |
| C. amphoroides | Lanceolate to slightly constricted | 28–45 | 3.5–7 | 30–32 | Variable | Slight helictoglossae | Distinct | Straight | Unknown | Unknown | Frauenfeld’s marin Aquarium | Marine | [7] |
| C. alyoubii | Linear, slightly-constricted | 83–105 | 6–10 | ∼40 | Similar | Rectelevatum + Knob | Indistinct | Straight | 4–5 | Prominent | Duba, Saudi Arabia | Marine | [6] |
| C. danayanus | Linear, very slightly constricted | 28–61 | 2–2.5 | 49–51 | Similar | Rectelevatum | Indistinct | Straight | 6–8 | Absent | Mabibi Beach, Elephant Coast, South Africa | Marine (carapace of a sea turtle) | [8] |
| C. legouvelloanus a | Linear to linear-lanceolate, slightly constricted | 18–34 (23–39) | 3–5 (–6) | 46–49 (40–44) | Similar | Rectelevatum + knob with central cavity | Indistinct | Clearly expanded | 4 | Well developed | Kosi Bay, South Africa | Marine (carapace of a sea turtle) | [8] |
| C. macewanii | Linear to linear-lanceolate, slightly constricted | 26–51 | 4.5–5.5 | 28–31 | Similar | Rectelevatum + knob | Distinct | Straight | Highly variable | Rudimentary | uShaka Sea World, Durban, South Africa | Marine (carapace of a sea turtle) | [8] |
| C. paradoxus | Linear, slightly-constricted | 80–85 | 6.5–9 | 36–40 | Similar; can be longer near valve edge | Rectelevatum + Knob | Indistinct | Straight | 4–5 | Prominent | Gab Gab reef, Apra Harbor Guam, USA | Marine | [6] |
| C. alatus a | Linear to linear-lanceolate, slightly constricted | 20–37 (16–38) | 3–5 (5–7) | 26–28 (22–25) | Variable | Rectelevatum | Distinct | Very slightly expanded | Highly variable | Rudimentary | Riverhead, New York, USA | Marine (carapace of a sea turtle) | [45] |
| C. laevissimus | Linear to linear-lanceolate, without consctiction | 37–66 | 5.5–7.8 | 24–27.5 | Similar | Helictoglossae | Unknown | Unknown | 3–6 | Unknown | Lakes and pools in the Larsemann Hills, Rauer Islands and Bølingen Islands, Antarctica | Brackish water | [46] |
| C. laevissimus (Tropidoneis laevissima) | Linear to linear-lanceolate, without consctiction | 27–79 | 5–9 | 20–28 | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Lakes and ponds in Kasumi Rock and on Shin-nan Rock, Antarctica | Brackish water | [47,48] |
| C. laevissimus | Linear to Eliptical | 49–98 | 8–9.5 | Absent | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Clear Lake, Green Lake, and lake on west side of McMurdo Sound, Antarctica | Fresh and Brackish water | [49] |
| C. indubitabilis (Stauronella indubitabilis) | Linear to eliptical | 25–60 | 6–7 | 25–27 | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Arctic Ocean | Marine | [50] |
| C. indubitabilis (Stauroneis constricta) | Lanceolate, sometimes slightly constricted | 30 | 5 | 23–27 | Similar | Rectelevatum + Knob | Unknown | Unknown | 4–5 | Unknown | – | – | [51] |
| C. indubitabilis | Linear to eliptical, sometimes slightly constricted | 16–27 | 4.4–7.6 | 22–30 | Variable | Rectelevatum + Knob | Unknown | Unknown | 3–13 | Rudimentary | Bahía Salada, Caldera, Chile | Marine costal water | [52] |
| C. infeffabilis | Linear | 31–58 | 4–7.5 | 25–36 | Similar | Helictoglossae | Indistinct | Straight | 4–7 | Absent | Inexpressible Island, Terra Nova Bay, Antarctica | Marine costal water | This study |
| C. zucchellii | Linear to lanceolate | 14.5–24 | 3–6 | 22–31 | Variable | Helictoglossae | Distinct | Straight | 4(+) | Absent | Zucchelli Station, Terra Nova Bay, Antarctica | Marine costal water | This study |
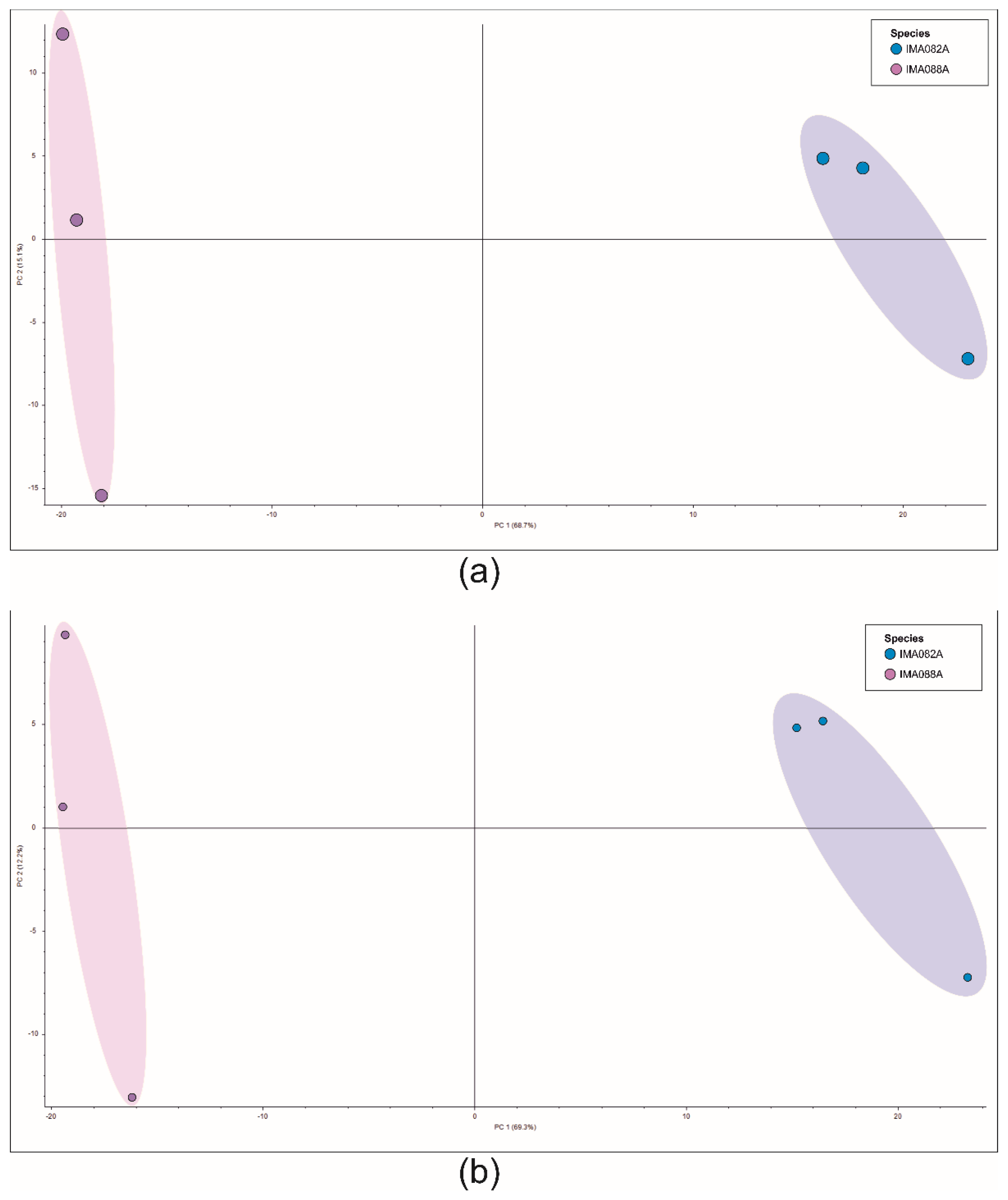
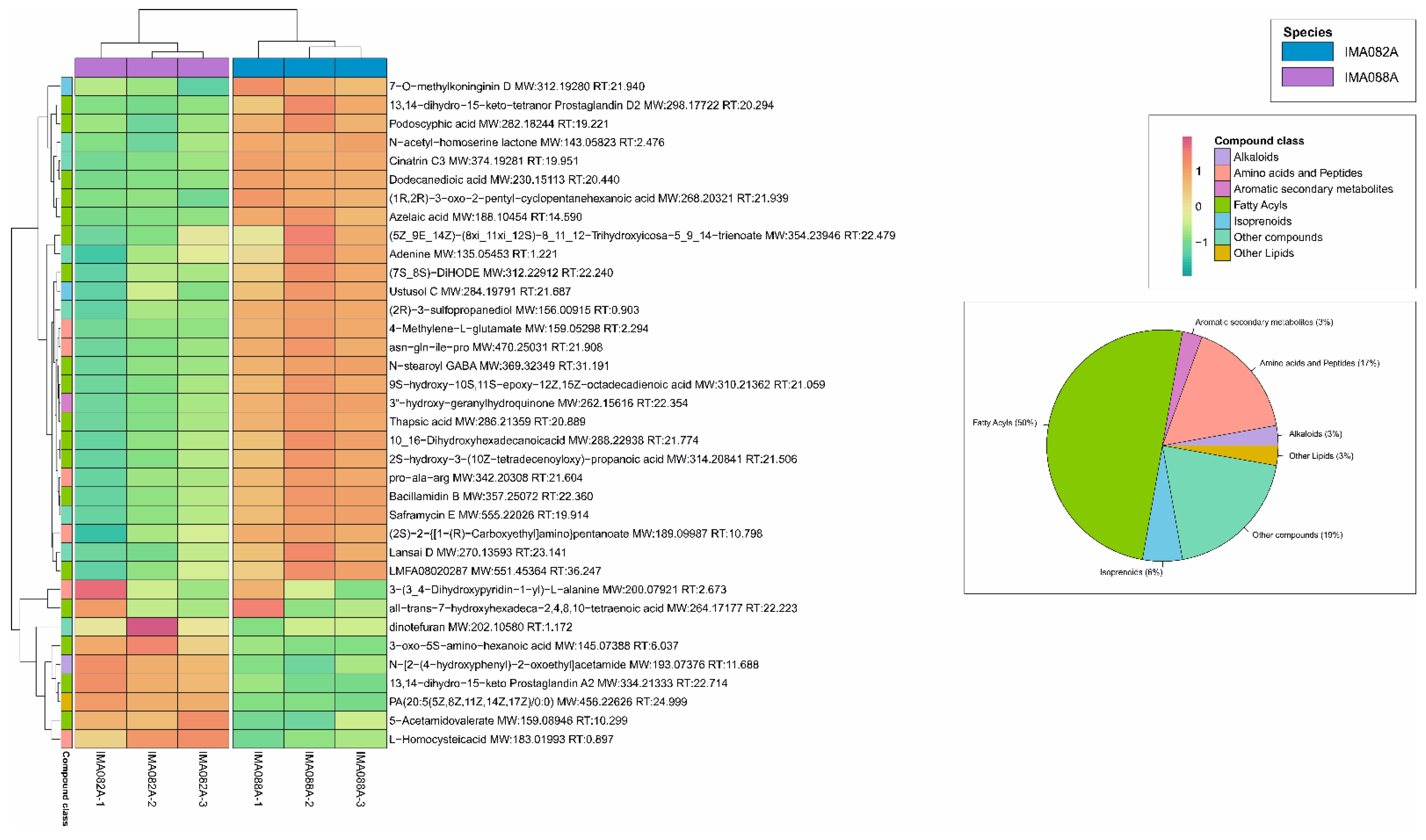
3.3. Liquid Chromatography-Mass Spectrometry (LC-MS)-Based Metabolomics Analysis
4. Discussion
4.1. Molecular Diversity
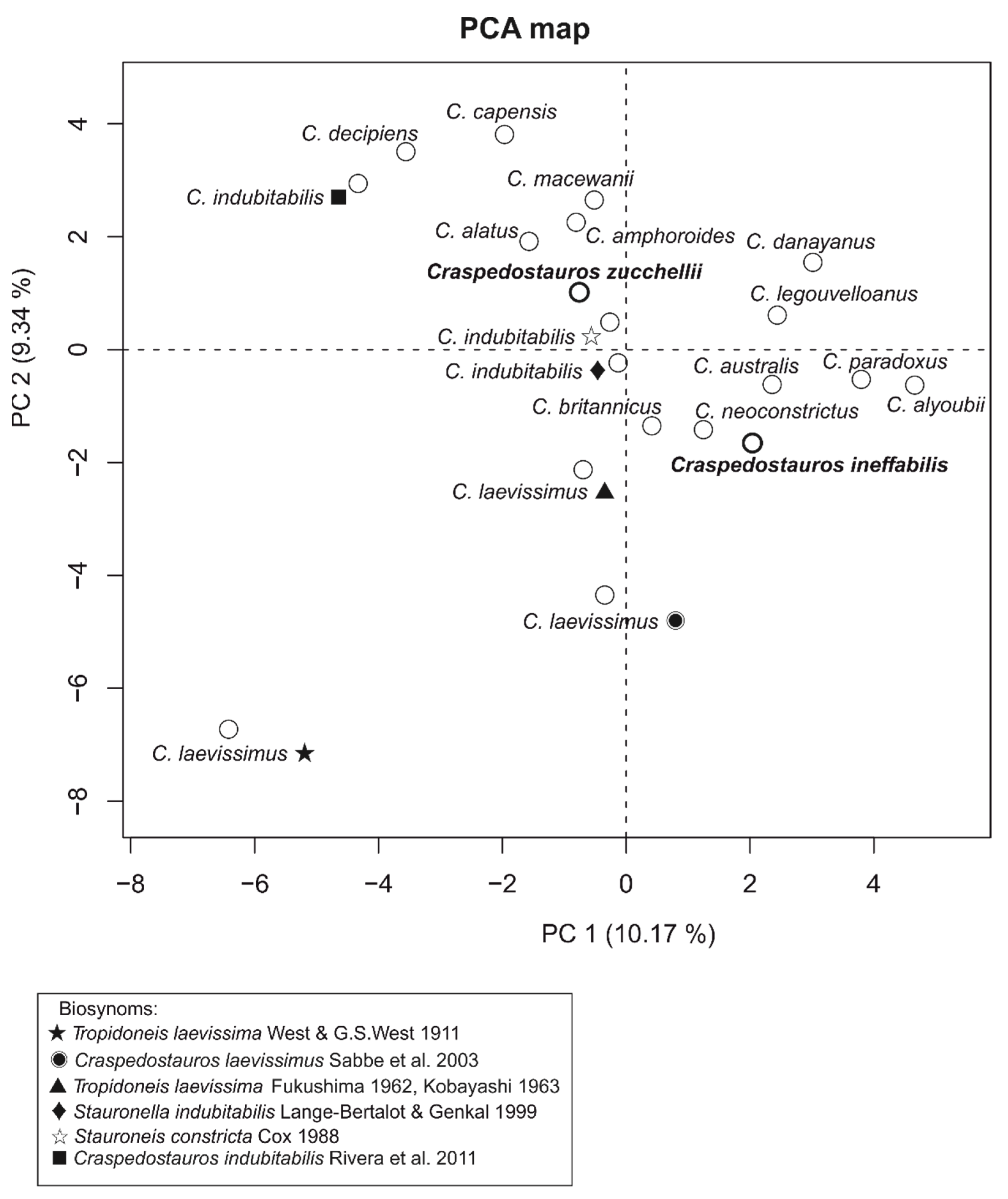
4.2. Morphological Diversity
4.3. Chemical Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huseby, S.; Degerlund, M.; Zingone, A.; Hansen, E. Metabolic fingerprinting reveals differences between northern and southern strains of the cryptic diatom Chaetoceros socialis. Eur. J. Phycol. 2012, 47, 480–489. [Google Scholar] [CrossRef]
- Kooistra, W.H.C.F.; De Stefano, M.; Mann, D.G.; Medlin, K. The Phylogeny of the Diatoms. In Silicon Biomineralization; Springer: Berlin/Heidelberg, Germany, 2003; pp. 59–97. [Google Scholar] [CrossRef]
- Hughes, A.H.; Magot, F.; Tawfike, A.F.; Rad-Menéndez, C.; Thomas, N.; Young, L.C.; Stucchi, L.; Carettoni, D.; Stanley, M.S.; Edrada-Ebel, R.; et al. Exploring the Chemical Space of Macro- and Micro-Algae Using Comparative Metabolomics. Microorganisms 2021, 9, 311. [Google Scholar] [CrossRef] [PubMed]
- Sciuto, K.; Moschin, E.; Alongi, G.; Cecchetto, M.; Schiaparelli, S.; Caragnano, A.; Rindi, F.; Moro, I. Tethysphytum antarcticum gen. et sp. nov. (Hapalidiales, Rhodophyta), a new non-geniculate coralline alga from Terra Nova Bay (Ross Sea, Antarctica): Morpho-anatomical characterization and molecular phylogeny. Eur. J. Phycol. 2021, 56, 416–427. [Google Scholar] [CrossRef]
- Trentin, R.; Negrisolo, E.; Moschin, E.; Veronese, D.; Cecchetto, M.; Moro, I. Microglena antarctica sp. nov. a New Antarctic Green Alga from Inexpressible Island (Terra Nova Bay, Ross Sea) Revealed through an Integrative Approach. Diversity 2022, 14, 337. [Google Scholar] [CrossRef]
- Ashworth, M.P.; Theriot, E.C.; Jansen, R.K.; Lobban, C.S.; Witkowski, A.; Sabir, M.J.; Hajarah, N.H.; Sabir, J.S.; Jansen, R.K.; Baeshen, M.N.; et al. Molecular and Morphological Investigations of the Stauros-bearing, Raphid Pennate Diatoms (Bacillariophyceae): Craspedostauros E.J. Cox, and Staurotropis T.B.B. Paddock, and their Relationship to the Rest of the Mastogloiales. Protist 2017, 168, 48–70. [Google Scholar] [CrossRef]
- Cox, E.J. Craspedostauros gen. nov., a new diatom genus for some unusual marine raphid species previously placed in Stauroneis Ehrenberg and Stauronella Mereschkowsky. Eur. J. Phycol. 1999, 34, 131–147. [Google Scholar] [CrossRef]
- Majewska, R.; Ashworth, M.P.; Bosak, S.; Goosen, W.E.; Nolte, C.; Filek, K.; Van de Vijver, B.; Taylor, J.C.; Manning, S.R.; Nel, R. On Sea Turtle-associated Craspedostauros (Bacillariophyta), with Description of Three Novel Species. J. Phycol. 2021, 57, 199–218. [Google Scholar] [CrossRef]
- Cox, E.; Williams, D. Systematics of naviculoid diatoms: The interrelationships of some taxa with a stauros. Eur. J. Phycol. 2000, 35, 273–282. [Google Scholar] [CrossRef]
- Smedsgaard, J.; Nielsen, J. Metabolite profiling of fungi and yeast: From phenotype to metabolome by MS and informatics. J. Exp. Bot. 2005, 56, 273–286. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef]
- Marcellin-Gros, R.; Piganeau, G.; Stien, D. Metabolomic Insights into Marine Phytoplankton Diversity. Mar. Drugs 2020, 18, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.G.; Paula, P.; da Silva, J.P.; Mil-Homens, D.; Teixeira, M.C.; Fialho, A.M.; Costa, R.; Keller-Costa, T. Insights into the Antimicrobial Activities and Metabolomes of Aquimarina (Flavobacteriaceae, Bacteroidetes) Species from the Rare Marine Biosphere. Mar. Drugs 2022, 20, 423. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar] [CrossRef]
- Dawson, S.C.; Pace, N.R. Novel kingdom-level eukaryotic diversity in anoxic environments. Proc. Natl. Acad. Sci. USA 2002, 99, 8324–8329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, S.J.; DeLong, E.F.; Olsen, G.J.; Pace, N.R. Phylogenetic group-specific oligodeoxynucleotide probes for identification of single microbial cells. J. Bacteriol. 1988, 170, 720–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgcomb, V.P.; Kysela, D.T.; Teske, A.; de Vera Gomez, A.; Sogin, M.L. Benthic eukaryotic diversity in the Guaymas Basin hydrothermal vent environment. Proc. Natl. Acad. Sci. USA 2002, 99, 7658–7662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Stoeck, T.; Schwarz, M.V.J.; Boenigk, J.; Schweikert, M.; von der Heyden, S.; Behnke, A. Cellular identity of an 18S rRNA gene sequence clade within the class Kinetoplastea: The novel genus Actuariola gen. nov. (Neobodonida) with description of the type species Actuariola framvarensis sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 2623–2635. [Google Scholar] [CrossRef]
- Hugerth, L.W.; Muller, E.E.L.; Hu, Y.O.O.; Lebrun, L.A.M.; Roume, H.; Lundin, D.; Wilmes, P.; Andersson, A.F. Systematic Design of 18S rRNA Gene Primers for Determining Eukaryotic Diversity in Microbial Consortia. PLoS ONE 2014, 9, e95567. [Google Scholar] [CrossRef]
- Alverson, A.J.; Jansen, R.K.; Theriot, E.C. Bridging the Rubicon: Phylogenetic analysis reveals repeated colonizations of marine and fresh waters by thalassiosiroid diatoms. Mol. Phylogenet. Evol. 2007, 45, 193–210. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D67–D72. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, E.P.; Kolbe, D.L.; Eddy, S.R. Infernal 1.0: Inference of RNA alignments. Bioinformatics 2009, 25, 1335–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA-X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gascuel, O. Approximate Likelihood-Ratio Test for Branches: A Fast, Accurate, and Powerful Alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Kishino, H.; Miyata, T.; Hasegawa, M. Maximum likelihood inference of protein phylogeny and the origin of chloroplasts. J. Mol. Evol. 1990, 31, 151–160. [Google Scholar] [CrossRef]
- Kishino, H.; Hasegawa, M. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in hominoidea. J. Mol. Evol. 1989, 29, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, H.; Hasegawa, M. Multiple Comparisons of Log-Likelihoods with Applications to Phylogenetic Inference. Mol. Biol. Evol. 1999, 16, 1114–1116. [Google Scholar] [CrossRef] [Green Version]
- Strimmer, K.; Rambaut, A. Inferring confidence sets of possibly misspecified gene trees. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, H. An Approximately Unbiased Test of Phylogenetic Tree Selection. Syst. Biol. 2002, 51, 492–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Santen, J.A.; Poynton, E.F.; Iskakova, D.; Mcmann, E.; Alsup, T.A.; Clark, T.N.; Fergusson, C.H.; Fewer, D.P.; Hughes, A.H.; Mccadden, C.A.; et al. The Natural Products Atlas 2.0: A database of microbially-derived natural products. Nucleic Acids Res. 2022, 50, D1317–D1323. [Google Scholar] [CrossRef]
- Sud, M.; Fahy, E.; Cotter, D.; Brown, A.; Dennis, E.A.; Glass, C.K.; Merrill, A.H.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; et al. LMSD: LIPID MAPS structure database. Nucleic Acids Res. 2007, 35, D527–D532. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0—The Human Metabolome Database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar] [CrossRef]
- Neveu, V.; Perez-Jiménez, J.; Vos, F.; Crespy, V.; du Chaffaut, L.; Mennen, L.; Knox, C.; Eisner, R.; Cruz, J.; Wishart, D.; et al. Phenol-Explorer: An online comprehensive database on polyphenol contents in foods. Database 2010, 2010, bap024. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Altman, T.; Billington, R.; Dreher, K.; Foerster, H.; Fulcher, C.A.; Holland, T.A.; Keseler, I.M.; Kothari, A.; Kubo, A.; et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nucleic Acids Res. 2014, 42, D459–D471. [Google Scholar] [CrossRef] [Green Version]
- Majewska, R.; Ashworth, M.P.; Lazo-Wasem, E.; Robinson, N.J.; Rojas, L.; Van de Vijver, B.; Pinou, T. Craspedostauros alatus sp. nov., a new diatom (Bacillariophyta) species found on museum sea turtle specimens. Diatom Res. 2018, 33, 229–240. [Google Scholar] [CrossRef]
- Sabbe, K.; Verleyen, E.; Hodgson, D.A.; Vanhoutte, K.; Vyverman, W. Benthic diatom flora of freshwater and saline lakes in the Larsemann Hills and Rauer Islands, East Antarctica. Antarct. Sci. 2003, 15, 227–248. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, H. Diatoms from the Shin-Nan Rock Ice-Free Area, Prince Olav Coast, the Antarctic Continent. Antarct. Rec. 1962, 14, 80–91. [Google Scholar]
- Kobayashi, T. Variations on Some Pennate Diatoms from Antarctica. Japanese Antarctic Research Expedition 19561962. Sci. Rep. Ser. E Biol. 1963, 18, 20. [Google Scholar]
- West, W.; West, G.S. Freshwater Algae. In British Antarctic Expedition 1907-9, under the Command of Sir; Part VII; Shackleton, E.H., Ed.; Forgotten Books: London, UK, 1911; Volume 1, pp. 263–298. [Google Scholar]
- Lange-Bertalot, H. Diatomeen aus Sibirien I. Inseln im Arktischen Osean (Yugorsky-Shar Strait). Iconogr. Diatomol. 1999, 6, 1–271. [Google Scholar]
- Cox, E.J. Taxonomic studies on the diatom genus Navicula V. The establishment of Parlibellus ge. nov. for some members of Navicula sect. Microstigmaticae. Diatom Res. 1988, 3, 9–38. [Google Scholar] [CrossRef]
- Abou-Shanab, R.A.I.; Hwang, J.H.; Cho, Y.; Min, B.; Jeon, B.H. Characterization of microalgal species isolated from fresh water bodies as a potential source for biodiesel production. Appl. Energy 2011, 88, 3300–3306. [Google Scholar] [CrossRef]
- Rivera, P.; Cruces, F.; Ector, L. Comentarios y observaciones acerca de la diatomea Craspedostauros indubitabilis (Lange-Bertalot & Genkal) E.J. Cox encontrada por primera vez en aguas marinas costeras de Chile. Gayana. Botánica 2011, 68, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Watanuki, T. Isolation and culture of Antarctic diatoms from the saline lakes in the Soya Coast, East Antarctica. Mem. Natl. Inst. Polar Res. 1979, 11, 35–41. [Google Scholar]
- Hodgson, D.A.; Noon, P.E.; Vyverman, W.; Bryant, C.L.; Gore, D.B.; Appleby, P.; Gilmour, M.; Verleyen, E.; Sabbe, K.; Jones, V.J.; et al. Were the Larsemann Hills ice-free through the Last Glacial Maximum? Antarct. Sci. 2001, 13, 440–454. [Google Scholar] [CrossRef]
- Prada, C.; Schizas, N.V.; Yoshioka, P.M. Phenotypic plasticity or speciation? A case from a clonal marine organism. BMC Evol. Biol. 2008, 8, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, A.; Pancha, I.; Jain, D.; Paliwal, C.; Ghosh, T.; Patidar, S.; Bhattacharya, S.; Mishra, S. Fatty acids as biomarkers of microalgae. Phytochemistry 2013, 89, 53–58. [Google Scholar] [CrossRef]
- Goutte, A.; Cherel, Y.; Houssais, M.N.; Klein, V.; Ozouf-Costaz, C.; Raccurt, M.; Robineau, C.; Massé, G. Diatom-Specific Highly Branched Isoprenoids as Biomarkers in Antarctic Consumers. PLoS ONE 2013, 8, e56504. [Google Scholar] [CrossRef] [Green Version]
- Ivanišević, J.; Thomas, O.P.; Lejeusne, C.; Chevaldonné, P.; Pérez, T. Metabolic fingerprinting as an indicator of biodiversity: Towards understanding inter-specific relationships among Homoscleromorpha sponges. Metabolomics 2011, 7, 289–304. [Google Scholar] [CrossRef]








| Tree | logL | deltaL 1 | bp-RELL 2 | p-KH 3 | p-SH 4 | p-WKH 5 | p-WSH 6 | c-ELW 7 | p-AU 8 |
|---|---|---|---|---|---|---|---|---|---|
| BT | −51.868 | 0 | 0.646+ | 0.642+ | 1+ | 0.642+ | 0.862+ | 0.638+ | 0.627+ |
| CA | −51.870 | 2 | 0.354+ | 0.358+ | 0.712+ | 0.358+ | 0.646+ | 0.362+ | 0.373+ |
| CAM | −51.945 | 77 | 0− | 0− | 0− | 0− | 0− | 4.04 × 10−12− | 1.94 × 10−07− |
| CSAM | −51.977 | 108.99 | 0− | 0− | 0− | 0− | 0− | 1.05 × 10−18− | 3.04 × 10−47− |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trentin, R.; Moschin, E.; Duarte Lopes, A.; Schiaparelli, S.; Custódio, L.; Moro, I. Molecular, Morphological and Chemical Diversity of Two New Species of Antarctic Diatoms, Craspedostauros ineffabilis sp. nov. and Craspedostauros zucchellii sp. nov. J. Mar. Sci. Eng. 2022, 10, 1656. https://doi.org/10.3390/jmse10111656
Trentin R, Moschin E, Duarte Lopes A, Schiaparelli S, Custódio L, Moro I. Molecular, Morphological and Chemical Diversity of Two New Species of Antarctic Diatoms, Craspedostauros ineffabilis sp. nov. and Craspedostauros zucchellii sp. nov. Journal of Marine Science and Engineering. 2022; 10(11):1656. https://doi.org/10.3390/jmse10111656
Chicago/Turabian StyleTrentin, Riccardo, Emanuela Moschin, André Duarte Lopes, Stefano Schiaparelli, Luísa Custódio, and Isabella Moro. 2022. "Molecular, Morphological and Chemical Diversity of Two New Species of Antarctic Diatoms, Craspedostauros ineffabilis sp. nov. and Craspedostauros zucchellii sp. nov." Journal of Marine Science and Engineering 10, no. 11: 1656. https://doi.org/10.3390/jmse10111656