

Germination Ecology of African Lovegrass (Eragrostis curvula) and Herbicide Options for Its Control



Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
2.2. General Germination Protocol
2.3. Experiment 1: Alternating Day/Night Temperatures
2.4. Experiment 2: Effect of Light
2.5. Experiment 3: Sodium Chloride (NaCl) Concentration
2.6. Experiment 4: Osmotic Potential
2.7. Experiment 5: Seed Burial Depth
2.8. Experiment 6: Sorghum Residue Amount
2.9. Experiment 7: Performance of Postemergence (POST) Herbicides
2.10. Statistical Analyses
3. Results
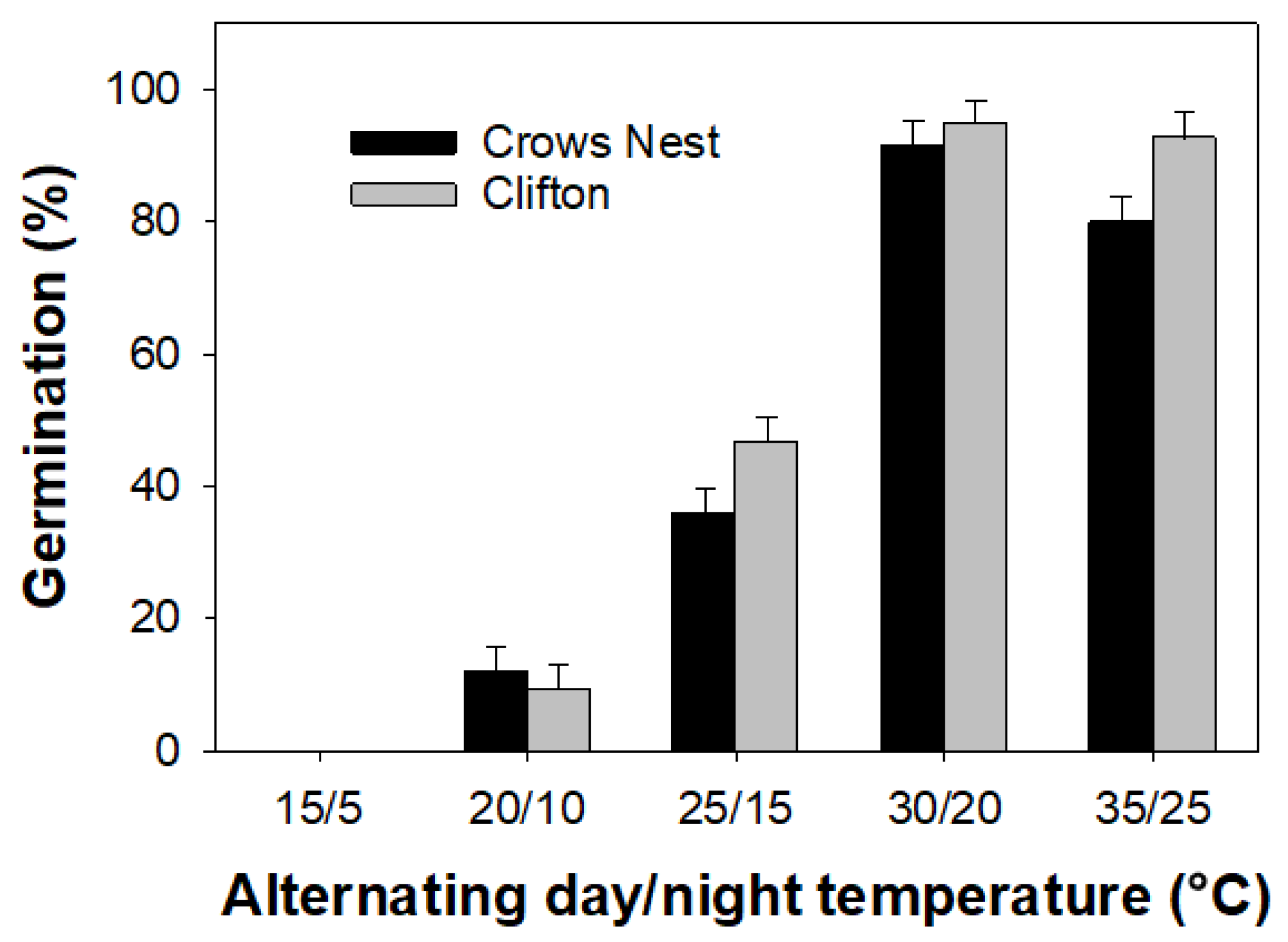
3.1. Experiment 1: Temperature
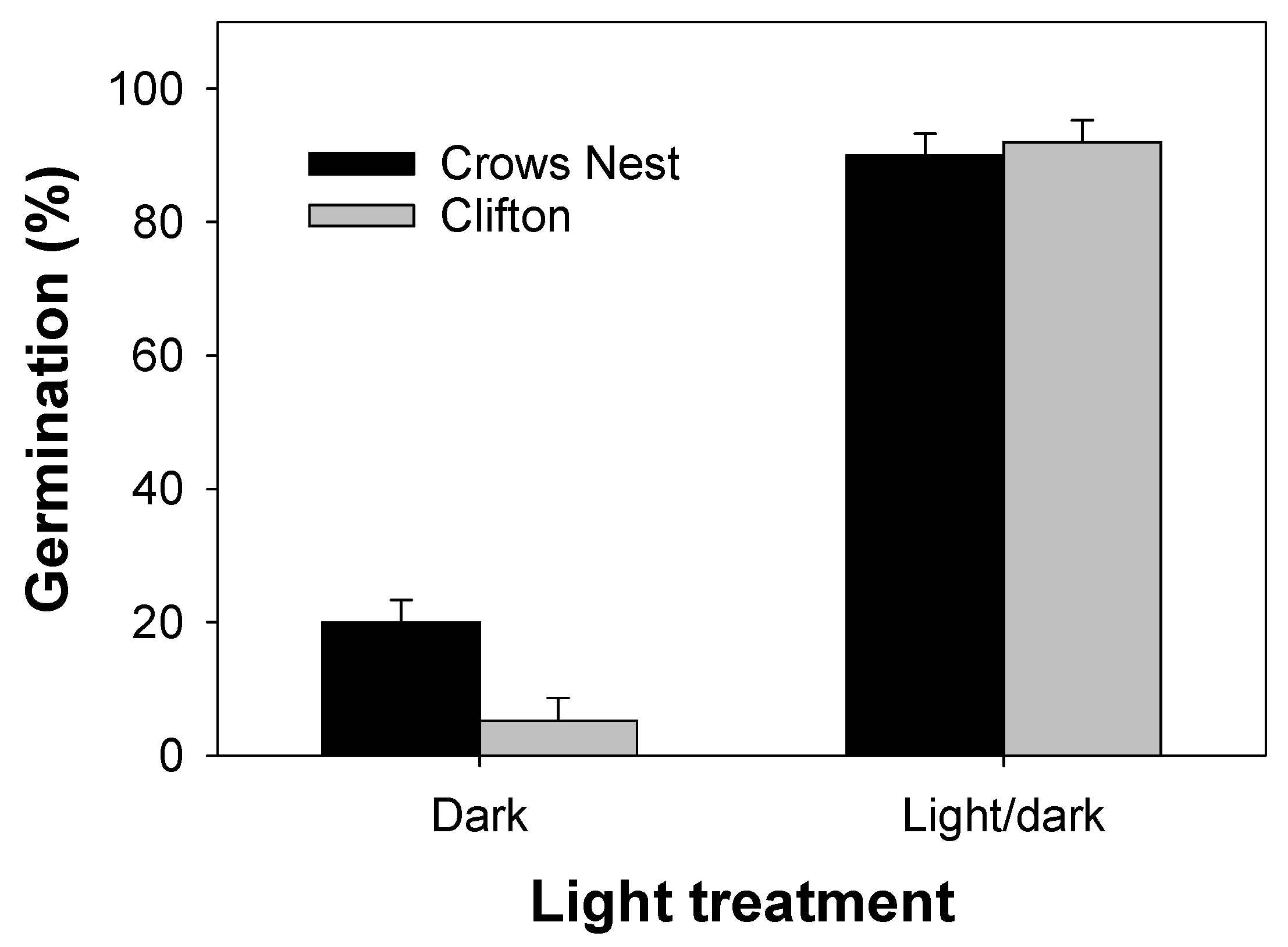
3.2. Experiment 2: Light
3.3. Experiment 3: Salt Stress
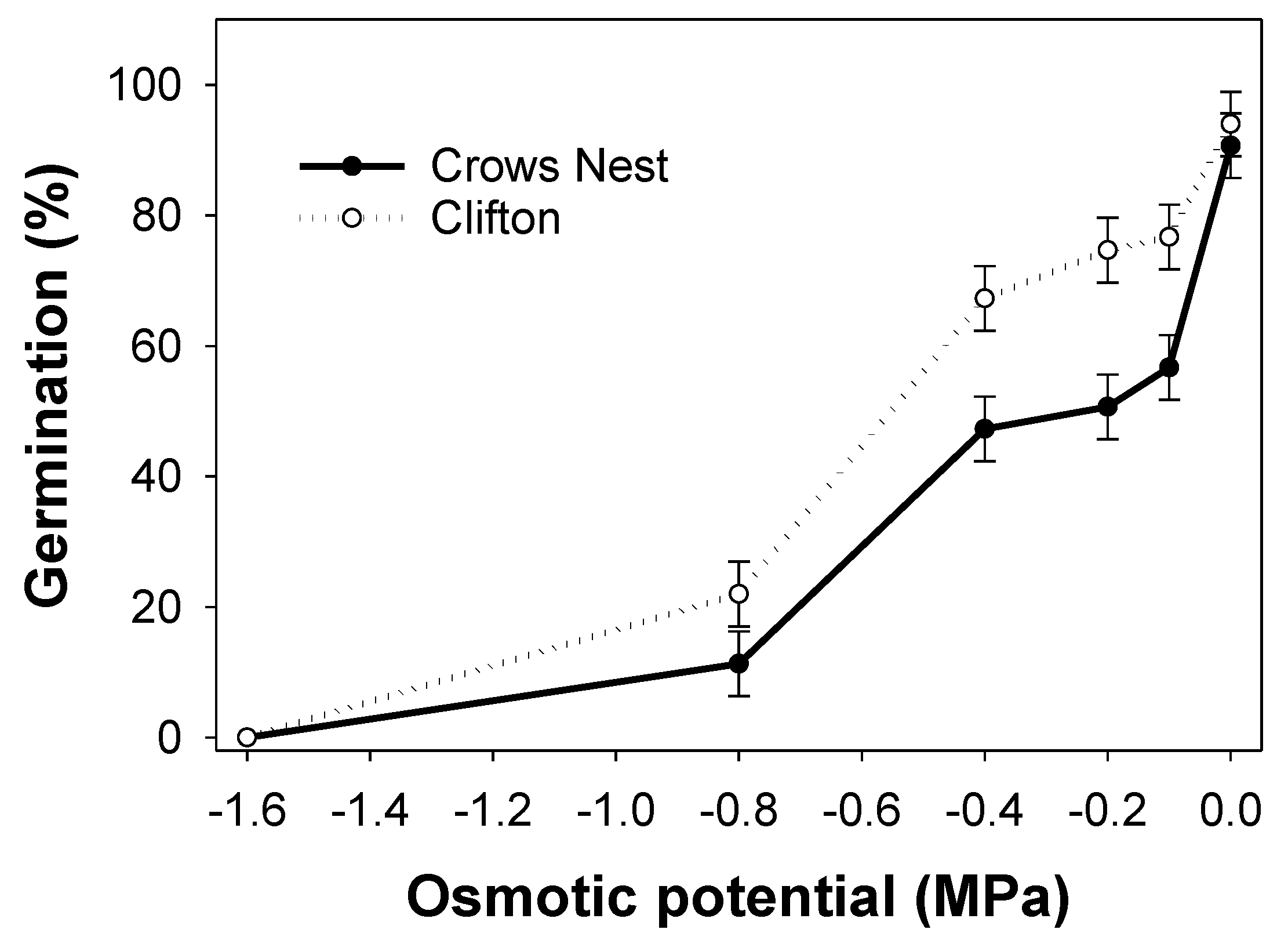
3.4. Experiment 4: Water Stress
3.5. Experiment 5: Burial Depth
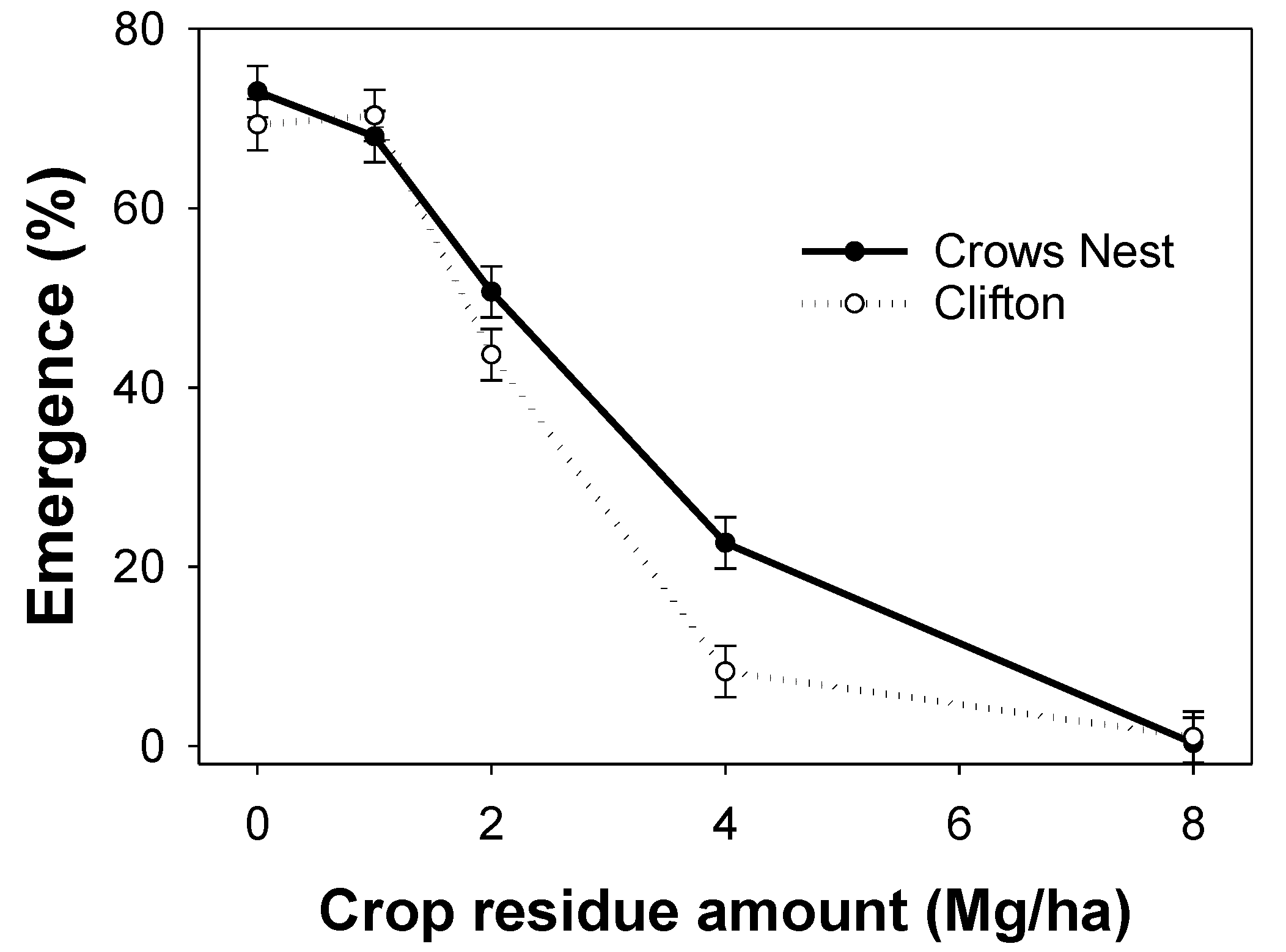
3.6. Experiment 6: Sorghum Residue Cover
3.7. Experiment 7: Post-Emergent Herbicides
4. Discussion
4.1. Experiment 1: Temperature
4.2. Experiment 2: Light
4.3. Experiment 3: Salt Stress
4.4. Experiment 4: Water Stress
4.5. Experiment 5: Burial Depth
4.6. Experiment 6: Sorghum Residue Cover
4.7. Experiment 7: Post-Emergent Herbicides
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, J.; Florentine, S.; van Etten, E.; Turville, C. Germination biology, distribution and control of the invasive species Eragrostis curvula [Schard. Nees] (African lovegrass): A global synthesis of current and future management challenges. Weed Res. 2021, 61, 154–163. [Google Scholar] [CrossRef]
- International, E. Eragrostis curvula Global Database. 2020. Available online: www.cabi.org/isc/datasheet/21630#REF-DDB-183911 (accessed on 29 October 2020).
- Firn, J. African lovegrass in Australia: A valuable pasture species or embarrassing invader ? Trop. Grassl. 2009, 43, 86–97. [Google Scholar]
- Firn, J.; MacDougall, A.S.; Schmidt, S.; Buckley, Y.M. Early emergence and resource availability can competitively favour natives over a functionally similar invader. Oecologia 2010, 163, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Buckley, Y.M.; Firn, J. An invasive grass shows colonization advantages over native grasses under conditions of low resource availability. Plant Ecol. 2012, 213, 1117–1130. [Google Scholar] [CrossRef]
- Johnston, W.; Cregan, P. The pastoral and soil conservation potential of Eragrostis curvula in semiarid New South Wales. In Proceedings of the Seventh Asian–Pacific Weed Science Society Conference, Sydney, Australia, 26–30 November 1979; Medd, R.W., Auld, B.A., Eds.; Council of Australian Weed Science Societies: Sydney, Australia, 1979; pp. 161–164. [Google Scholar]
- Lazarides, M. A revision of Eragrostis (Eragrostideae, Eleusininae, Poaceae) in Australia. Aust. Syst. Bot. 1997, 10, 77–187. [Google Scholar] [CrossRef]
- Johnston, W.H.; Shoemaker, V.F. Establishment and persistence of palatable taxa of Eragrostis curvula complex in southern New South Wales. Aust. J. Exp. Agric. 1997, 37, 55–65. [Google Scholar] [CrossRef]
- Parson, W.T.; Cuthbertson, E. Noxious Weeds of Australia; CSIRO Publishing: Collingwood, Australia, 2001. [Google Scholar]
- Walters, C.; Wheeler, L.M.; Grotenhuis, J.M. Longevity of seeds stored in a genebank: Species characteristics. Seed Sci. Res. 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Roberts, J.; Florentine, S.; Van Etten, E.; Turville, C. Seed longevity and germination in response to changing drought and heat conditions on four populations of the invasive weed African lovegrass (Eragrostis curvula). Weed Sci. 2021, 69, 468–477. [Google Scholar] [CrossRef]
- Leigh, J.H.; Davidson, R.L. Eragrostis curvula (Schrad.) Nees and some other African lovegrasses. Plant Introd. Rev. 1968, 5, 21–44. [Google Scholar]
- Rodrigo, J.M.; Zappacosta, D.C.; Selva, J.P.; Garbus, I.; Albertini, E.; Echenique, V. Apomixis frequency under stress conditions in weeping lovegrass (Eragrostis curvula). PLoS ONE 2017, 12, e0175852. [Google Scholar] [CrossRef]
- Roberts, J.; Florentine, S.; Van Etten, E.; Turville, C. Germination biology of four climatically varied populations of the invasive species African lovegrass (Eragrostis curvula). Weed Sci. 2021, 69, 210–218. [Google Scholar] [CrossRef]
- Campbell, M.H.; Kemp, H.W.; Murison, R.D.; Dellow, J.J.; Ridings, H. Use of herbicides for selective removal of Eragrostis curvula (Schrad.) Nees from a Pennisetum clandestinum pasture. Aust. J. Exp. Agric. 1987, 27, 359–365. [Google Scholar] [CrossRef]
- Firn, J.; Ladouceur, E.; Dorrough, J. Integrating local knowledge and research to refine the management of an invasive non-native grass in critically endangered grassy woodlands. J. Appl. Ecol. 2017, 55, 321–330. [Google Scholar] [CrossRef]
- Rengasamy, P. Transient salinity and subsoil constraints to dryland farming in Australian sodic soils: An overview. Aust. J. Exp. Agric. 2002, 42, 351–361. [Google Scholar] [CrossRef]
- Michel, B.E.; Radcliffe, D. A Computer Program Relating Solute Potential to Solution Composition for Five Solutes. Agron. J. 1995, 87, 126–130. [Google Scholar] [CrossRef]
- Genstat for Windows, 20th ed.; VSN International: Hemel Hempstead, UK, 2022; Available online: https://vsni.co.uk/software/genstat (accessed on 15 January 2023).
- Firn, J.; House, A.P.N.; Buckley, Y.M. Alternative states models provide an effective framework for invasive species control and restoration of native communities. J. Appl. Ecol. 2010, 47, 96–105. [Google Scholar] [CrossRef]
- Abrol, I.P.; Yadav, J.S.P.; Massoud, F.I. Salt-Affected Soils and Their Management. FAO Soils Bulletin 39, Food and Agriculture Organisation of the United Nations, Rome. 1988. Available online: http://www.fao.org/ (accessed on 15 January 2023).
- Weller, S.L.; Florentine, S.K.; Chauhan, B.S. Influence of selected environmental factors on seed germination and seedling emergence of Dinebra panicea var. brachiata (Steud.). Crop Prot. 2019, 117, 121–127. [Google Scholar] [CrossRef]
- Wang, H.; Wang, L.; Bai, S.; Guo, W.; Wang, J.; Liu, W. Germination ecology of giant chickweed (Myosoton aquaticum). Weed Sci. 2020, 68, 619–626. [Google Scholar] [CrossRef]
- Mutti, N.K.; Mahajan, G.; Chauhan, B.S. Seed-germination ecology of glyphosate-resistant and glyphosate-susceptible biotypes of Echinochloa colona in Australia. Crop Pasture Sci. 2019, 70, 367–372. [Google Scholar] [CrossRef]
- Mobli, A.; Mollaee, M.; Manalil, S.; Chauhan, B.S. Germination Ecology of Brachiaria eruciformis in Australia and Its Implications for Weed Management. Agronomy 2020, 10, 30. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Manalil, S.; Florentine, S.; Jha, P. Germination ecology of Chloris truncata and its implication for weed management. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Chauhan, B.S. Germination biology of liverseedgrass (Urochloa panicoides) and its response to postemergence herbicides in Australian conditions. Weed Sci. 2022, 70, 553–560. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicide Treatments | Trade Names | Site of Action | Dose (g a.i./a.e. ha−1) | Adjuvant |
|---|---|---|---|---|
| Control | - | - | - | - |
| Amitrole + paraquat | Alliance® | Lycopene cyclase inhibitor + Photosystem I inhibitor | 560 | - |
| Butroxydim | Factor® | ACCase inhibitor | 30 | 1% Uptake® |
| Clethodim | Havoc® | ACCase inhibitor | 120 | 1% Supercharge® |
| Cyhalofop | Barnstorm | ACCase inhibitor | 285 | 1% Uptake® |
| Glufosinate | Biffo® | Glutamine synthetase inhibitor | 750 | - |
| Glyphosate | Roundup Ultra®MAX | EPSP inhibitor | 740 | - |
| Haloxyfop | VerdictTM | ACCase inhibitor | 78 | 1% HastenTM |
| Imazamox + imazapyr | Intervix® | ALS inhibitor | 36 | 1% HastenTM |
| Iodosulfuron | Hussar® | ALS inhibitor | 10 | - |
| Mesosulfuron | Atlantis® | ALS inhibitor | 10 | 1% HastenTM |
| Paraquat | Gramoxone® | Photosystem I inhibitor | 600 | 1% HastenTM |
| Pinoxaden | Axial® | ACCase inhibitor | 10 | 0.5% Adigor® |
| Propaquizafop | Shogun® | ACCase inhibitor | 30 | 1% HastenTM |
| Herbicides | Survival (%) | Spike (Number/Pot) | Biomass (g/pot) |
|---|---|---|---|
| Control | 100 | 3.3 | 2.603 |
| Amitrole + paraquat | 0 | 0 | 0 |
| Mesosulfuron | 100 | 2.9 | 2.398 |
| Pinoxaden | 100 | 3.5 | 2.806 |
| Glufosinate | 0 | 0 | 0 |
| Cyhalofop | 0 | 0 | 0 |
| Butroxydim | 0 | 0 | 0 |
| Glyphosate | 0 | 0 | 0 |
| Clethodim | 0 | 0 | 0 |
| Iodosulfuron | 100 | 4.2 | 2.89 |
| Imazamox + imazapyr | 100 | 0 | 0.711 |
| Paraquat | 0 | 0 | 0 |
| Propaquizafop | 0 | 0 | 0 |
| Haloxyfop | 0 | 0 | 0 |
| LSD | - | 0.47 | 0.187 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alagbo, O.O.; Chauhan, B.S. Germination Ecology of African Lovegrass (Eragrostis curvula) and Herbicide Options for Its Control. Agriculture 2023, 13, 920. https://doi.org/10.3390/agriculture13050920
Alagbo OO, Chauhan BS. Germination Ecology of African Lovegrass (Eragrostis curvula) and Herbicide Options for Its Control. Agriculture. 2023; 13(5):920. https://doi.org/10.3390/agriculture13050920
Chicago/Turabian StyleAlagbo, Oyebanji O., and Bhagirath Singh Chauhan. 2023. "Germination Ecology of African Lovegrass (Eragrostis curvula) and Herbicide Options for Its Control" Agriculture 13, no. 5: 920. https://doi.org/10.3390/agriculture13050920