
Combinatorial Olfactory Signaling in Short-Distance Determines Host Plant Recognition in Locust

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Locusts
2.2. Plants
2.3. Chemicals Preparation
2.4. Volatile Collection of Host and Nonhost Plants
2.5. Chemical Analysis
2.6. Duration of Excitatory Response Assay
2.7. Palp Opening Response Assay
2.8. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.9. RNA Interference
2.10. Statistical Analysis
3. Results
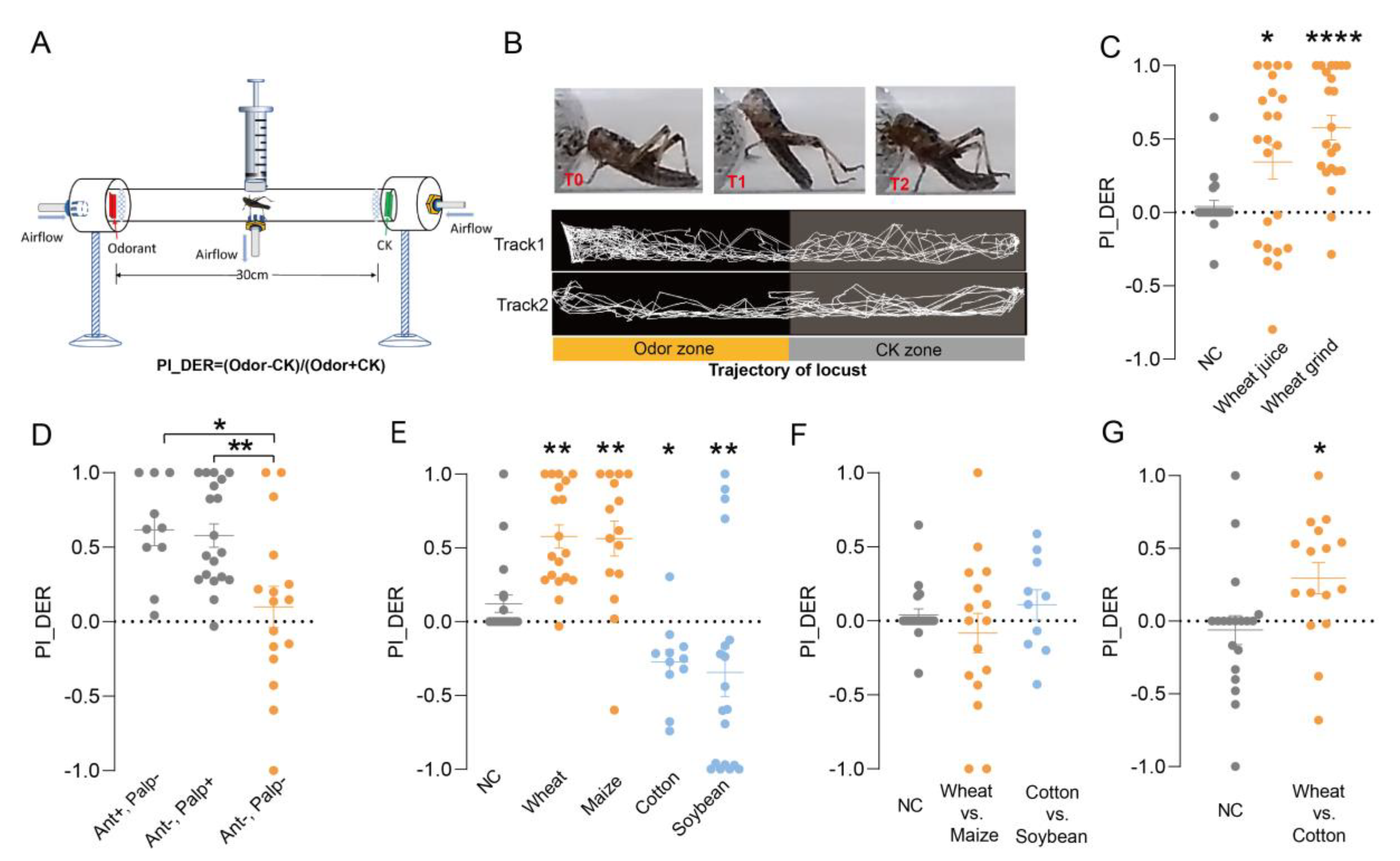
3.1. Establishment of Olfactory Feeding Behavior Responses of Palps
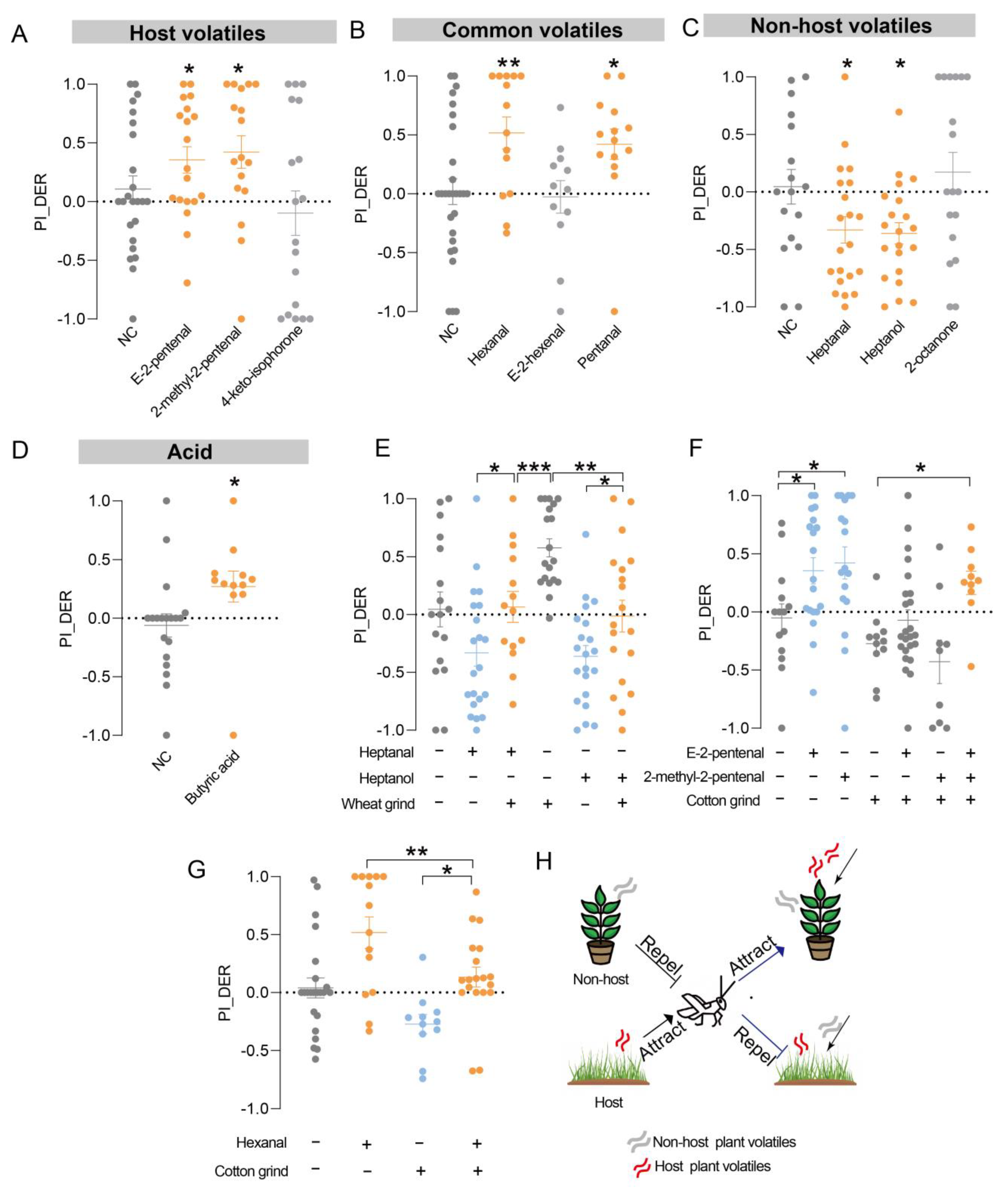
3.2. Volatiles from Host and Nonhost Plants Contribute to Food Selection in Locust
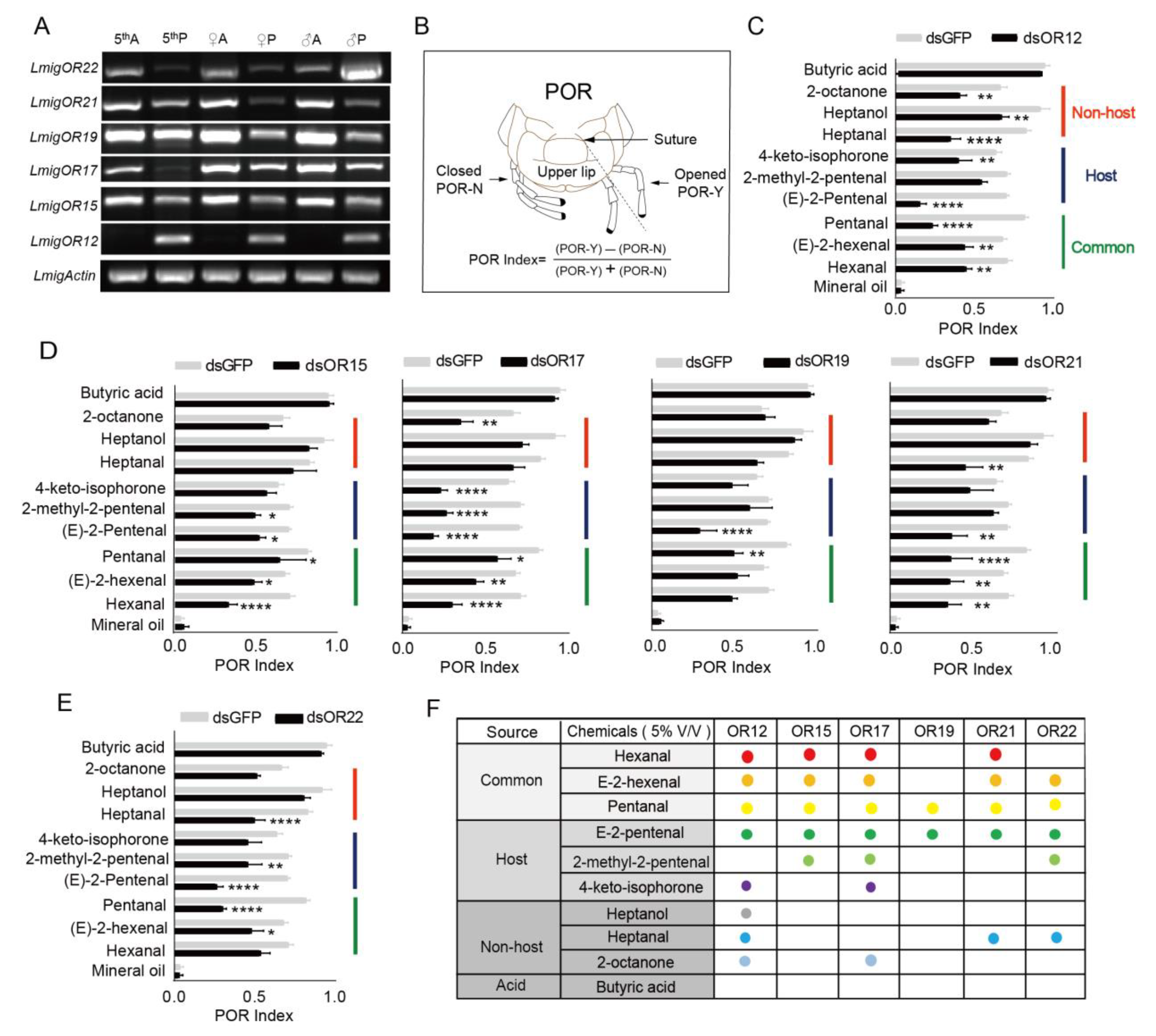
3.3. Odorant Receptors in Palps Involved in Detection of Odors from Host and Nonhost Plants
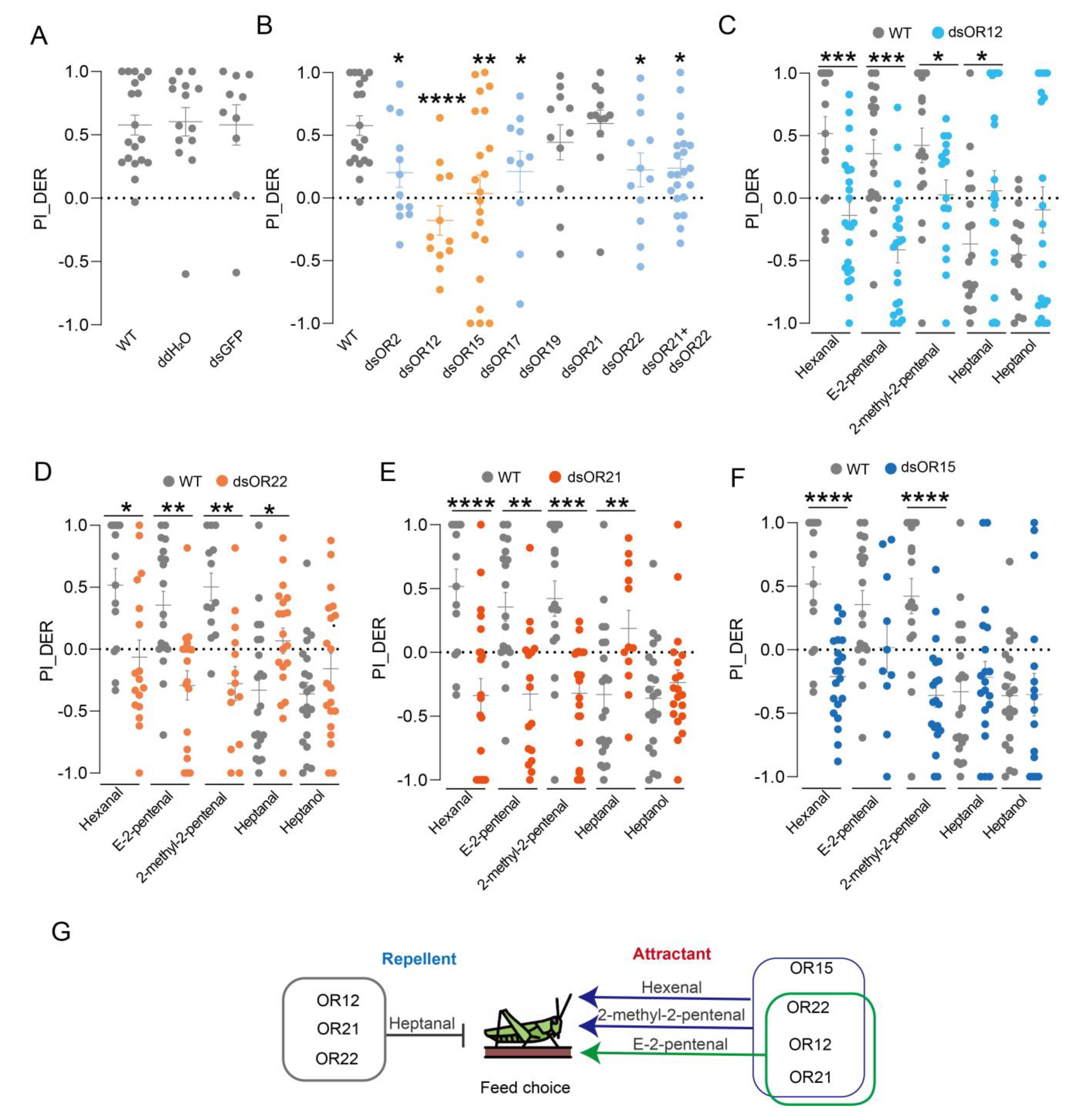
3.4. Combinatorial Olfactory Signaling Controls the Locust’s Food Choice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chapman, R.F. Contact chemoreception in feeding by phytophagous insects. Annu. Rev. Entomol. 2003, 48, 455–484. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T. Insect herbivore nutrient regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in Insect Olfaction: To Smell or Not to Smell? Annu. Rev. Entomol. 2016, 61, 317–333. [Google Scholar] [CrossRef]
- Sachse, S.; Krieger, J. Olfaction in insects. e-Neuroforum 2011, 17, 49–60. [Google Scholar] [CrossRef]
- Bartlet, E.; Blight, M.M.; Lane, P.; Williams, I.H. The responses of the cabbage seed weevil Ceutorhynchus assimilis to volatile compounds from oilseed rape in a linear track olfactometer. Entomol. Exp. Appl. 1997, 85, 257–262. [Google Scholar] [CrossRef]
- Thibout, E.; Auger, J.; Lecomte, C. Host plant chemicals responsible for attraction and oviposition in Acrolepiopsis assectella. In Proceedings of the 5th International Symposium on Insect-Plant Relationships, Wageningen, The Netherlands, 1–4 March 1982; pp. 107–116. [Google Scholar]
- Guerin, P.M.; Städler, E. Host odour perception in three phytophagous Diptera: A comparative study. In Proceedings of the 5th International Symposium on Insect-Plant Relationships, Wageningen, The Netherlands, 1–4 March 1982; pp. 95–106. [Google Scholar]
- Bowers, M.D.; Williams, E.H. Variable chemical defence in the checkerspot butterfly Euphydryas gillettii (Lepidoptera: NymphaIidae). Ecolog. Entomol. 1995, 20, 208–212. [Google Scholar] [CrossRef]
- Okamura, Y.; Dort, H.; Reichelt, M.; Tunström, K.; Wheat, C.W.; Vogel, H. Testing hypotheses of a coevolutionary key innovation reveals a complex suite of traits involved in defusing the mustard oil bomb. Proc. Natl. Acad. Sci. USA 2022, 119, e2208447119. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Schlyter, F. Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric. For. Entomol. 2004, 6, 1–19. [Google Scholar] [CrossRef]
- Simon, J.C.; d’Alencon, E.; Guy, E.; Jacquin-Joly, E.; Jaquiery, J.; Nouhaud, P.; Peccoud, J.; Sugio, A.; Streiff, R. Genomics of adaptation to host-plants in herbivorous insects. Brief Funct. Genomics 2015, 14, 413–423. [Google Scholar] [CrossRef]
- Li, N.; Li, S.; Ge, J.; Schuman, M.C.; Wei, J.N.; Ma, R.Y.; Bonser, S. Manipulating two olfactory cues causes a biological control beetle to shift to non-target plant species. J. Ecol. 2017, 105, 1534–1546. [Google Scholar] [CrossRef]
- de Fouchier, A.; Sun, X.; Caballero-Vidal, G.; Travaillard, S.; Jacquin-Joly, E.; Montagne, N. Behavioral Effect of Plant Volatiles Binding to Spodoptera littoralis Larval Odorant Receptors. Front. Behav. Neurosci. 2018, 12, 264. [Google Scholar] [CrossRef] [PubMed]
- Finch, S.; Collier, R.H. Host-plant selection by insects-a theory based on ‘appropriate/inappropriate landings’ by pest insects of cruciferous plants. Entomol. Exp. Et Appl. 2000, 96, 91–102. [Google Scholar] [CrossRef]
- Clark, J.T.; Ray, A. Olfactory Mechanisms for Discovery of Odorants to Reduce Insect-Host Contact. J. Chem. Ecol. 2016, 42, 919–930. [Google Scholar] [CrossRef]
- Carrasco, D.; Larsson, M.C.; Anderson, P. Insect host plant selection in complex environments. Curr. Opin. Insect Sci. 2015, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.R.; Benton, R. Molecular mechanisms of olfactory detection in insects: Beyond receptors. Open Biol. 2020, 10, 200252. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the odor world: Olfactory receptors and their role for signal transduction in insects. Cell Mol. Life Sci. 2018, 75, 485–508. [Google Scholar] [CrossRef]
- Xiao, Y.; An, X.K.; Khashaveh, A.; Shan, S.; Wang, Q.; Wang, S.N.; Gu, S.H.; Li, Z.Y.; Zhang, Y.J. Broadly Tuned Odorant Receptor AlinOR59 Involved in Chemoreception of Floral Scent in Adelphocoris lineolatus. J. Agric. Food Chem. 2020, 68, 13815–13823. [Google Scholar] [CrossRef]
- Wen, M.; Li, E.; Chen, Q.; Kang, H.; Zhang, S.; Li, K.; Wang, Y.; Jiao, Y.; Ren, B. A herbivore-induced plant volatile of the host plant acts as a collective foraging signal to the larvae of the meadow moth, Loxostege sticticalis (Lepidoptera: Pyralidae). J. Insect Physiol. 2019, 118, 103941. [Google Scholar] [CrossRef]
- Haverkamp, A.; Hansson, B.S.; Knaden, M. Combinatorial Codes and Labeled Lines: How Insects Use Olfactory Cues to Find and Judge Food, Mates, and Oviposition Sites in Complex Environments. Front. Physiol. 2018, 9, 49. [Google Scholar] [CrossRef]
- Blaney, W.M.; Duckett, A.M. The significance of palpation by the maxillary palps of Locusta migratoria (L): An electrophysiological and behavioural study. J. Exp. Biol. 1975, 63, 701–712. [Google Scholar] [CrossRef]
- Tsuneto, K.; Endo, H.; Shii, F.; Sasaki, K.; Nagata, S.; Sato, R. Diet choice: The two-factor host acceptance system of silkworm larvae. PLoS Biol. 2020, 18, e3000828. [Google Scholar] [CrossRef] [PubMed]
- Luntz, A.J.M. The role of the maxillary and labial palps in the feeding behavior of Schistocerca Gregaria. Entomol. Exp. Appl. 1979, 25, 279–288. [Google Scholar] [CrossRef]
- Kasubuchi, M.; Shii, F.; Tsuneto, K.; Yamagishi, T.; Adegawa, S.; Endo, H.; Sato, R. Insect taste receptors relevant to host identification by recognition of secondary metabolite patterns of non-host plants. Biochem. Biophys. Res. Commun. 2018, 499, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Amat, C.; Marion-Poll, F.; Navarro-Roldan, M.A.; Gemeno, C. Gustatory function of sensilla chaetica on the labial palps and antennae of three tortricid moths (Lepidoptera: Tortricidae). Sci. Rep. 2022, 12, 18882. [Google Scholar] [CrossRef]
- Li, H.W.; Wang, P.; Zhang, L.W.; Xu, X.; Cao, Z.W.; Zhang, L. Expressions of Olfactory Proteins in Locust Olfactory Organs and a Palp Odorant Receptor Involved in Plant Aldehydes Detection. Front. Physiol. 2018, 9, 663. [Google Scholar] [CrossRef]
- Wang, P.; Yin, X.; Zhang, L. Plant Approach-Avoidance Response in Locusts Driven by Plant Volatile Sensing at Different Ranges. J. Chem. Ecol. 2019, 45, 410–419. [Google Scholar] [CrossRef]
- Zhang, L.W.; Li, H.W.; Zhang, L. Two Olfactory Pathways to Detect Aldehydes on Locust Mouthpart. Int. J. Biol. Sci. 2017, 13, 759–771. [Google Scholar] [CrossRef]
- Sun, L.; Pan, X.; Li, H.; Zhang, X.; Zhao, X.; Zhang, L.; Zhang, L. Odor-Induced Vomiting Is Combinatorially Triggered by Palp Olfactory Receptor Neurons That Project to the Lobus Glomerulatus in Locust Brain. Front. Physiol. 2022, 20, 855522. [Google Scholar] [CrossRef]
- Buttery, R.G.; Ling, L.C. Corn leaf volatiles: Identification using tenax trapping for possible insect attractants. J. Agric. Food Chem. 1984, 32, 1104–1106. [Google Scholar] [CrossRef]
- Buttery, R.G.; Xu, C.J.; Ling, L.C. Volatile components of wheat leaves (and stems): Possible insect attractants. J. Agric. Food Chem. 1985, 33, 115–117. [Google Scholar] [CrossRef]
- Zeringue, H.J.; McCormick, S.P. Relationships between cotton leaf-derived volatiles and growth of Aspergillus flavus. J. Am. Oil Chem. Soc. 1989, 66, 581–585. [Google Scholar] [CrossRef]
- Njagi, G.N.N.; Torto, B. Responses of nymphs of desert locust, Schistocerca gregaria, to volatiles of plants used as rearing diet. Chemoecology 1996, 7, 172–178. [Google Scholar] [CrossRef]
- Shibamoto, T.; Horiuchi, M.; Umano, K. Composition of the young green barley and wheat leaves. J. Essent. Oil Res. 2007, 19, 134–137. [Google Scholar] [CrossRef]
- Pan, H.S.; Zhao, Q.J.; Zhao, K.J.; Zhang, Y.J.; Wu, K.M.; Guo, Y.Y. Application of electronic nose zNose (TM) for construction of volatiles fingerprint library and analysis of real time release rhythm of volatiles from damaged cotton. Chin. J. Appl. Environ. Biol. 2010, 16, 468–473. [Google Scholar]
- Michereff, M.F.F.; Laumann, R.A.; Borges, M.; Michereff, M.; Diniz, I.R.; Neto, A.L.F.; Moraes, M.C.B. Volatiles mediating a plant-herbivore-natural enemy interaction in resistant and susceptible soybean cultivars. J. Chem. Ecol. 2011, 37, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.X.; Xu, W.; Lu, A.J.; Bi, J.R.; Su, X.; Shi, S.S. Relationship between components change of soybean volatile and Leguminivora glycinivorella infestation. J. Jilin Agric. Univ. 2014, 36, 389–394. [Google Scholar]
- Leppik, E.; Frerot, B. Maize field odorscape during the oviposition flight of the European corn borer. Chemoecology 2014, 24, 221–228. [Google Scholar] [CrossRef]
- Bruce, T.J.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Dethier, V.G. Mechanism of Host-Plant Recognition. Entomol. Exp. Et Appl. 2011, 31, 49–56. [Google Scholar] [CrossRef]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects--finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Dweck, H.K.; Ebrahim, S.A.; Khallaf, M.A.; Koenig, C.; Farhan, A.; Stieber, R.; Weissflog, J.; Svatos, A.; Grosse-Wilde, E.; Knaden, M.; et al. Olfactory channels associated with the Drosophila maxillary palp mediate short- and long-range attraction. Elife 2016, 23, e14925. [Google Scholar] [CrossRef]
- Knolhoff, L.M.; Heckel, D.G. Behavioral assays for studies of host plant choice and adaptation in herbivorous insects. Annu. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Hansson, B.S.; Larsson, M.C.; Leal, W.S. Green leaf volatile-detecting olfactory receptor neurones display very high sensitivity and specificity in a scarab beetle. Physiol. Entomol. 2002, 24, 121–126. [Google Scholar] [CrossRef]
- Zhao, Y.X.; Kang, L. Role of plant volatiles in host plant location of the leafminer, Liriomyza sativae (Diptera: Agromyzidae). Physiol. Entomol. 2002, 27, 103–111. [Google Scholar] [CrossRef]
- von Arx, M.; Schmidt-Busser, D.; Guerin, P.M. Plant volatiles enhance behavioral responses of grapevine moth males, Lobesia botrana to sex pheromone. J. Chem. Ecol. 2012, 38, 222–225. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, S.-G.; Zhang, L. Expression of odorant-binding and chemosensory proteins and spatial map of chemosensilla on labial palps of Locusta migratoria (Orthoptera: Acrididae). Arthropod. Struct. Dev. 2006, 35, 47–56. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, X.; Liu, J.; Xu, X.; Zhang, L.; Zhang, L. Combinatorial Olfactory Signaling in Short-Distance Determines Host Plant Recognition in Locust. Agriculture 2023, 13, 1030. https://doi.org/10.3390/agriculture13051030
Pan X, Liu J, Xu X, Zhang L, Zhang L. Combinatorial Olfactory Signaling in Short-Distance Determines Host Plant Recognition in Locust. Agriculture. 2023; 13(5):1030. https://doi.org/10.3390/agriculture13051030
Chicago/Turabian StylePan, Xueqin, Jun Liu, Xiao Xu, Liwei Zhang, and Long Zhang. 2023. "Combinatorial Olfactory Signaling in Short-Distance Determines Host Plant Recognition in Locust" Agriculture 13, no. 5: 1030. https://doi.org/10.3390/agriculture13051030