
Biochemical and Physiological Responses of Cucumis sativus L. to Application of Potential Bioinsecticides—Aqueous Carum carvi L. Seed Distillation By-Product Based Extracts
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of Plant Extracts and Commercial Agrochemicals
2.2. Plant Material and Plant Treatment
2.3. Photosynthesis and Morphological Parameters
2.4. Phytochemical Screening of Secondary Metabolites by 96-Wellplate Spectrophotometric Assays
2.5. The Identification and Quantification of Volatile Compounds in Cucumber Plant Leaves
2.6. Statistical Analysis
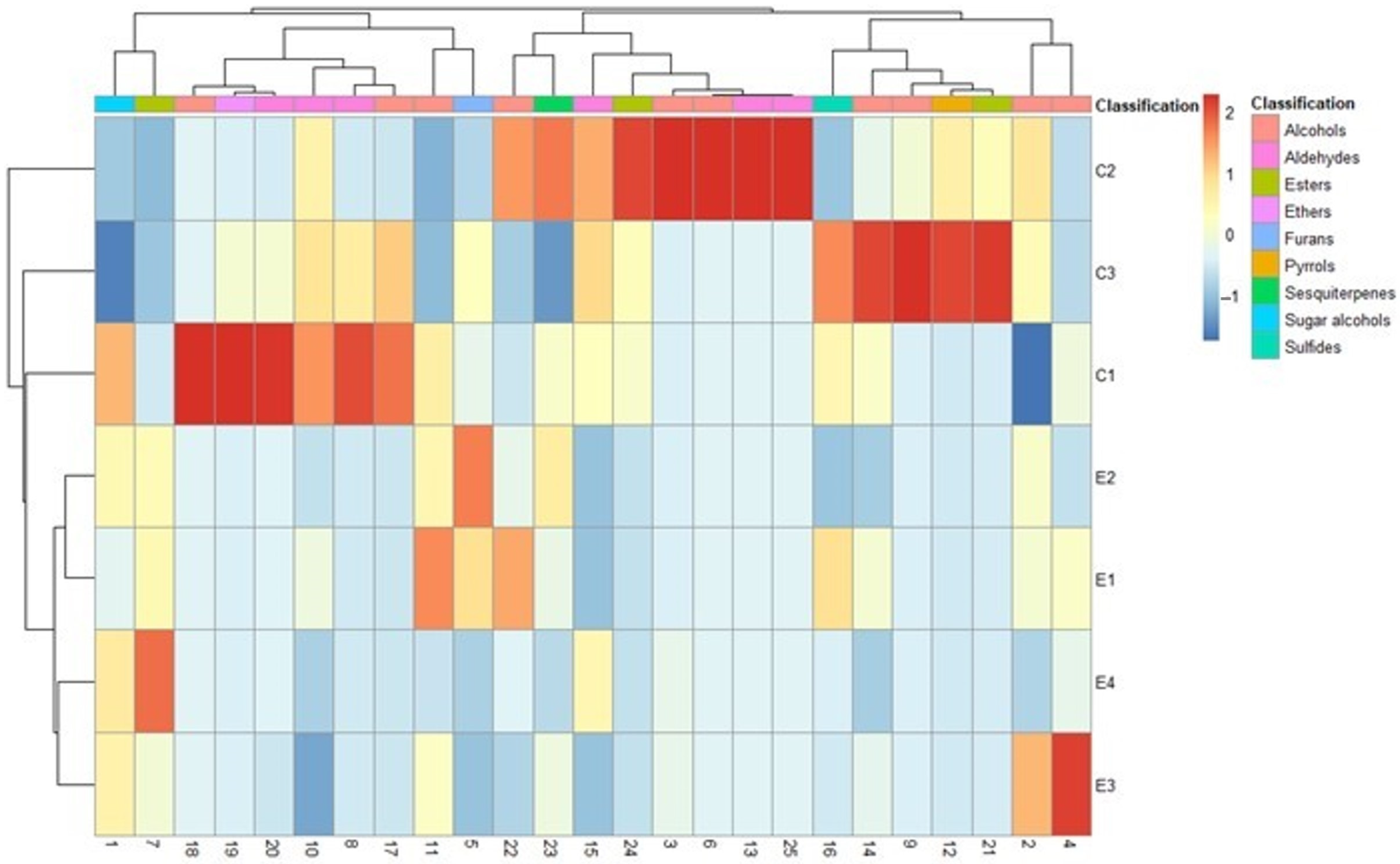
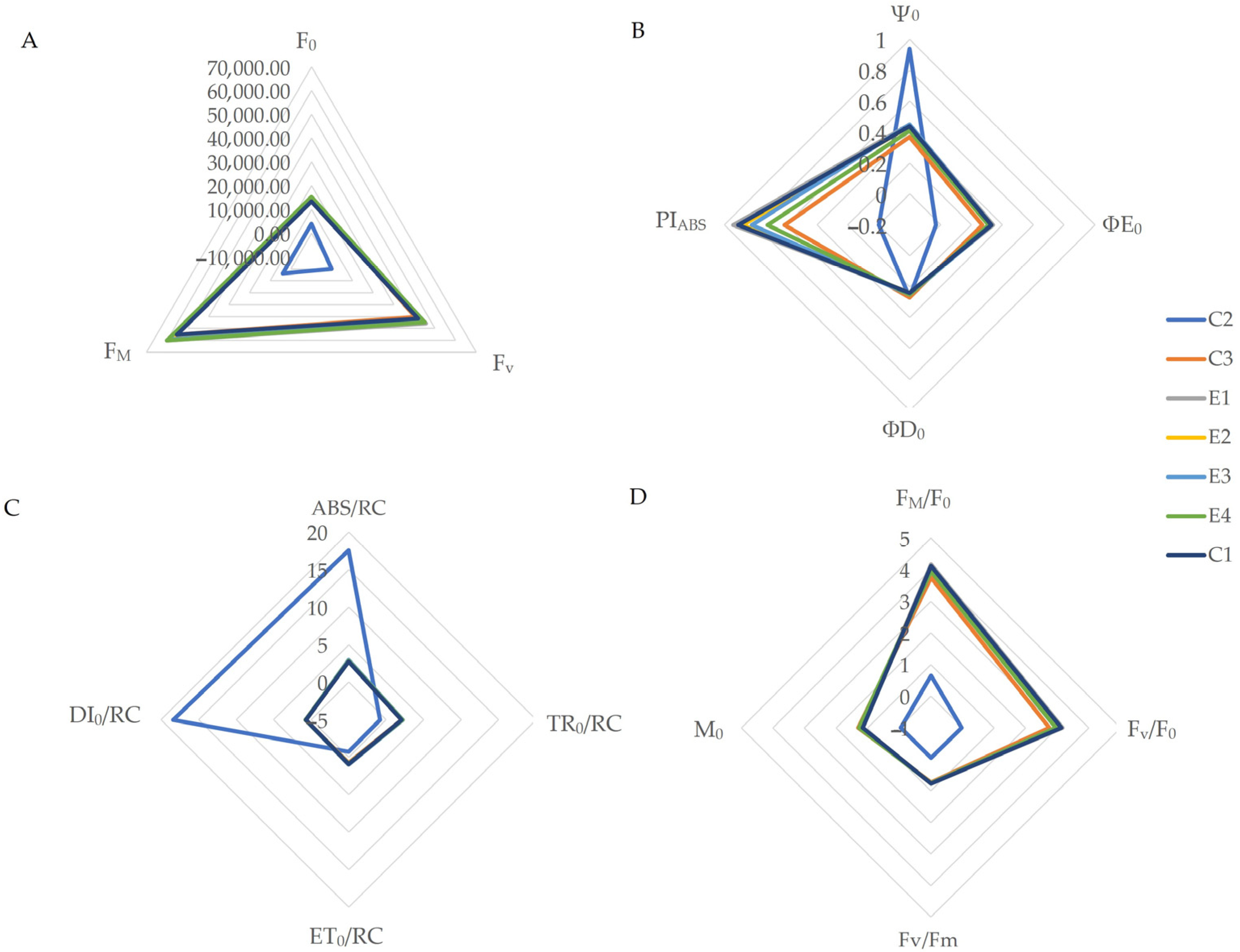
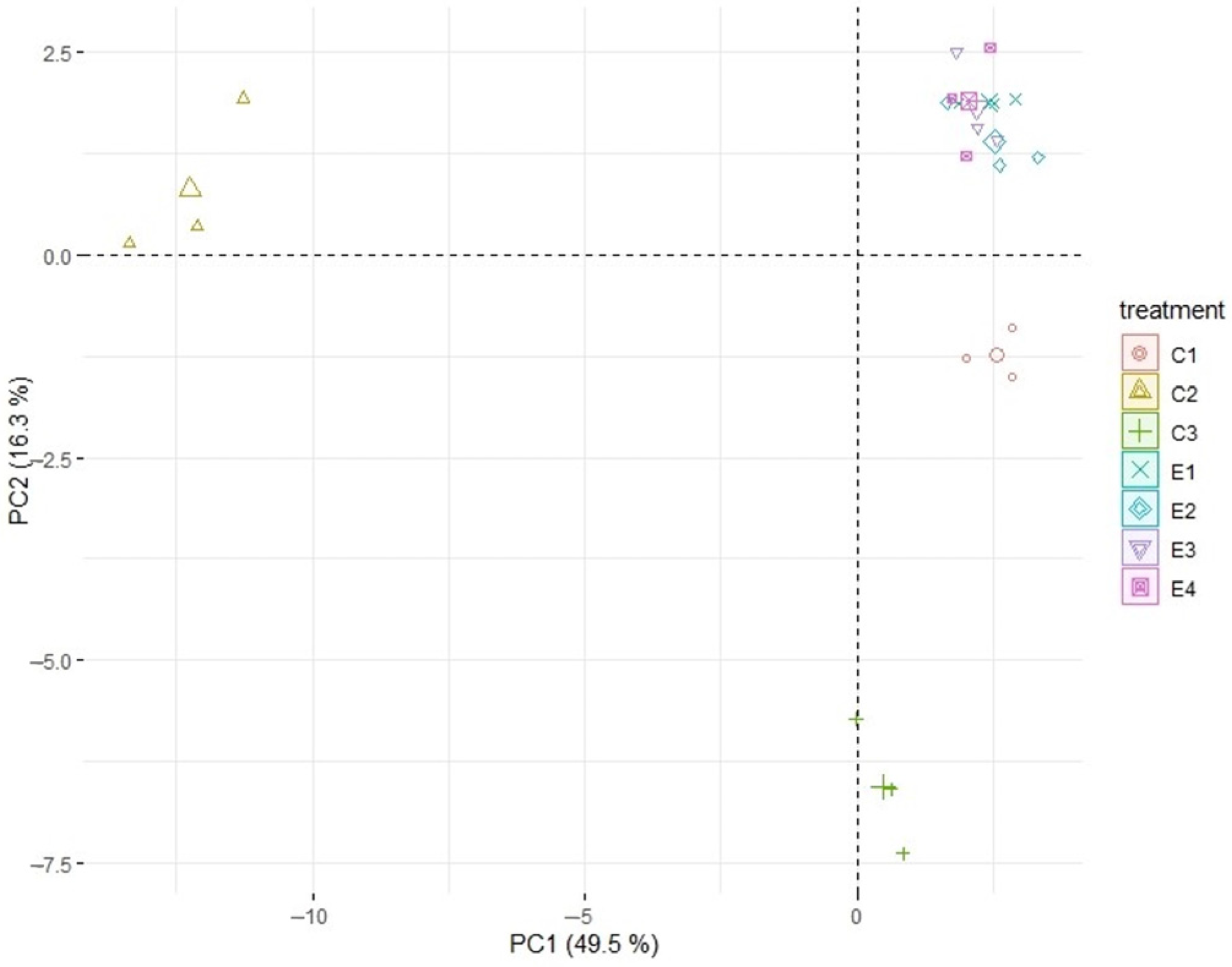
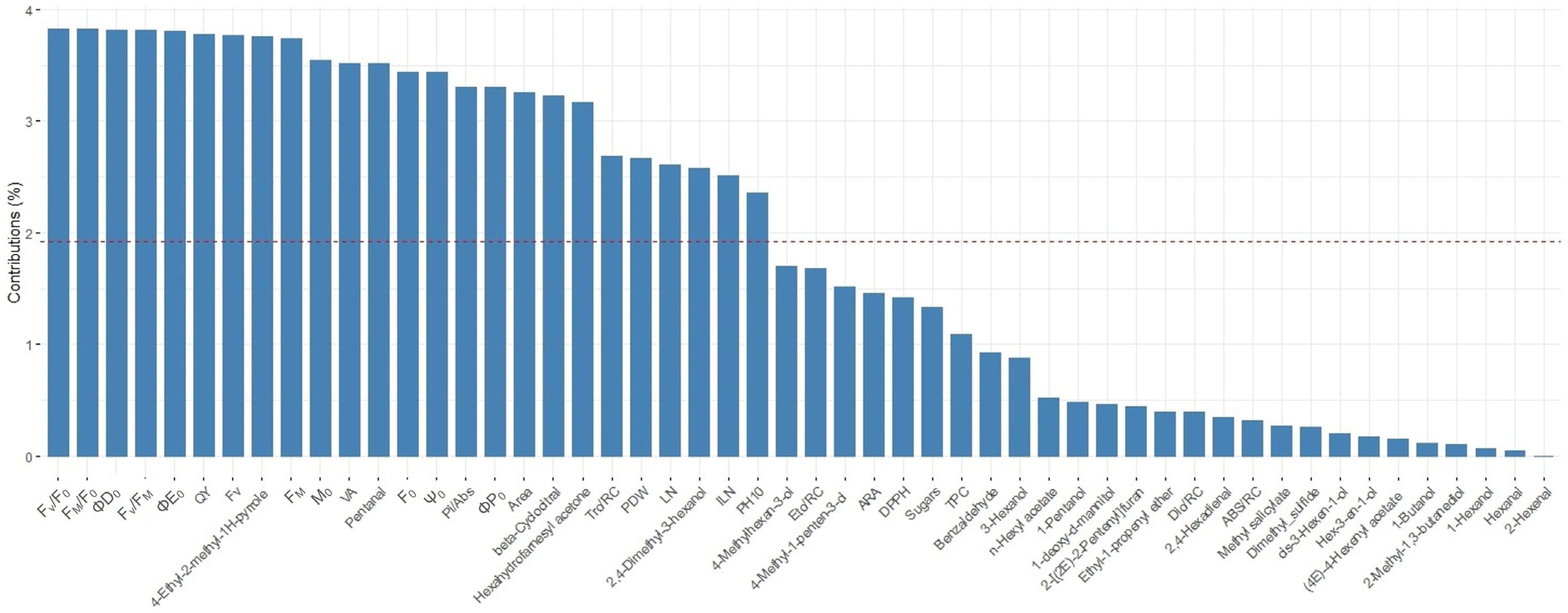
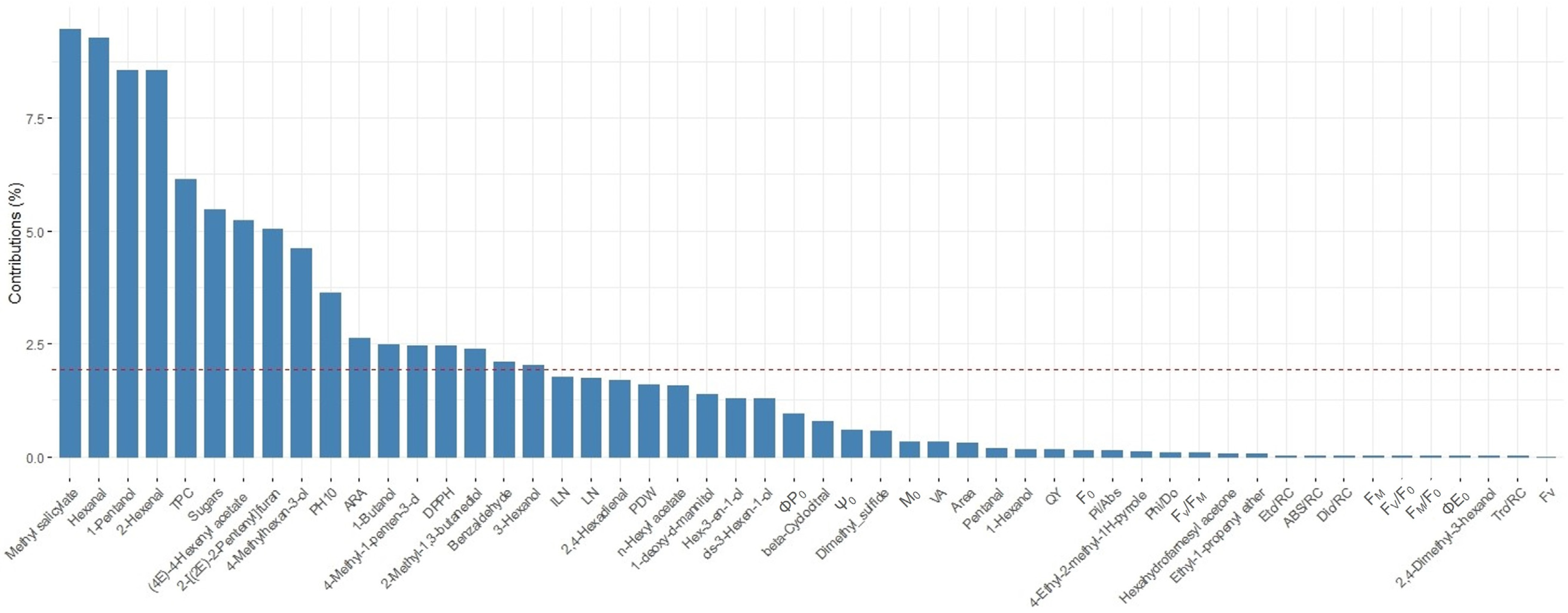
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acheuk, F.; Basiouni, S.; Shehata, A.A.; Dick, K.; Hajri, H.; Lasram, S.; Yilmaz, M.; Emekci, M.; Tsiamis, G.; Spona-Friedl, M. Status and Prospects of Botanical Biopesticides in Europe and Mediterranean Countries. Biomolecules 2022, 12, 311. [Google Scholar] [CrossRef]
- Tudi, M.; Ruan, H.D.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture Development, Pesticide Application and Its Impact on the Environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef]
- Tlak Gajger, I.; Dar, S.A. Plant Allelochemicals as Sources of Insecticides. Insects 2021, 12, 189. [Google Scholar] [CrossRef]
- Werrie, P.-Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.-L. Phytotoxicity of Essential Oils: Opportunities and Constraints for the Development of Biopesticides. A Review. Foods 2020, 9, 1291. [Google Scholar] [CrossRef] [PubMed]
- Eichenberg, D.; Ristok, C.; Kroeber, W.; Bruelheide, H. Plant Polyphenols–Implications of Different Sampling, Storage and Sample Processing in Biodiversity-Ecosystem Functioning Experiments. Chem. Ecol. 2014, 30, 676–692. [Google Scholar] [CrossRef]
- Qari, S.H.; Khalil, A.H.; Abdelfattah, N.A.H.; Shehawy, A.A. Effect of Different Plant Extracts and Nanoparticles on Thrips Tabaci (Lind.)(Thysanoptera: Thripidae) under Field Conditions and Their Allelopathic Potential on the Onion, Allium cepa L. Using Bioassays and RAPD Analysis. Egypt. J. Biol. Pest Control 2020, 30, 1–8. [Google Scholar] [CrossRef]
- Schnarr, L.; Segatto, M.L.; Olsson, O.; Zuin, V.G.; Kümmerer, K. Flavonoids as Biopesticides—Systematic Assessment of Sources, Structures, Activities and Environmental Fate. Sci. Total Environ. 2022, 824, 153781. [Google Scholar] [CrossRef]
- Rao, P.; Rathod, V. Valorization of Food and Agricultural Waste: A Step towards Greener Future. Chem. Rec. 2019, 19, 1858–1871. [Google Scholar] [CrossRef] [PubMed]
- Jensch, C.; Strube, J. Proposal of a New Green Process for Waste Valorization and Cascade Utilization of Essential Oil Plants. Sustainability 2022, 14, 3227. [Google Scholar] [CrossRef]
- Navarrete, A.; Herrero, M.; Martín, A.; Cocero, M.J.; Ibáñez, E. Valorization of Solid Wastes from Essential Oil Industry. J. Food Eng. 2011, 104, 196–201. [Google Scholar] [CrossRef]
- Saha, A.; Basak, B.B.; Manivel, P.; Kumar, J. Valorization of Java Citronella (Cymbopogon Winterianus Jowitt) Distillation Waste as a Potential Source of Phenolics/Antioxidant: Influence of Extraction Solvents. J. Food Sci. Technol. 2021, 58, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Truzzi, E.; Chaouch, M.A.; Rossi, G.; Tagliazucchi, L.; Bertelli, D.; Benvenuti, S. Characterization and Valorization of the Agricultural Waste Obtained from Lavandula Steam Distillation for Its Reuse in the Food and Pharmaceutical Fields. Molecules 2022, 27, 1613. [Google Scholar] [CrossRef] [PubMed]
- Slavov, A.; Vasileva, I.; Stefanov, L.; Stoyanova, A. Valorization of Wastes from the Rose Oil Industry. Rev. Environ. Sci. Bio. Technol. 2017, 16, 309–325. [Google Scholar] [CrossRef]
- Puglia, D.; Pezzolla, D.; Gigliotti, G.; Torre, L.; Bartucca, M.L.; Del Buono, D. The Opportunity of Valorizing Agricultural Waste, Through Its Conversion into Biostimulants, Biofertilizers, and Biopolymers. Sustainability 2021, 13, 2710. [Google Scholar] [CrossRef]
- Schroeder, P.; Pereira, V.B.; Ventura, A.; Andrade, J.; Figueiredo, M.K.-K.; das Chagas, E.F.; de Almeida Azevedo, D.; Romeiro, G.A.; da Silva, R.V.S. Chemical Evaluation via High-Resolution Techniques and Biopesticide Potential of Bio-Oil and Aqueous Phase from Slow Pyrolysis of Passion Fruit Seed Cake. Waste Biomass Valorization 2022, 1–22. [Google Scholar] [CrossRef]
- Preedy, V.R. Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2015; ISBN 012416644X. [Google Scholar]
- López, M.D.; Jordán, M.J.; Pascual-Villalobos, M.J. Toxic Compounds in Essential Oils of Coriander, Caraway and Basil Active against Stored Rice Pests. J. Stored Prod. Res. 2008, 44, 273–278. [Google Scholar] [CrossRef]
- Johri, R. Cuminum Cyminum and Carum Carvi: An Update. Pharmacogn. Rev. 2011, 5, 63. [Google Scholar] [CrossRef]
- Synowiec, A.; Możdżeń, K.; Krajewska, A.; Landi, M.; Araniti, F. Carum Carvi L. Essential Oil: A Promising Candidate for Botanical Herbicide against Echinochloa Crus-Galli (L.) P. beauv. in Maize Cultivation. Ind. Crops Prod. 2019, 140, 111652. [Google Scholar] [CrossRef]
- Füzy, A.; Kovács, R.; Cseresnyés, I.; Parádi, I.; Szili-Kovács, T.; Kelemen, B.; Rajkai, K.; Takács, T. Selection of Plant Physiological Parameters to Detect Stress Effects in Pot Experiments Using Principal Component Analysis. Acta Physiol. Plant. 2019, 41, 56. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T.; An, M. Screening Methods for the Evaluation of Crop Allelopathic Potential. Bot. Rev. 2001, 67, 403–415. [Google Scholar] [CrossRef]
- Yuan, B.-Z.; Bie, Z.-L.; Sun, J. Bibliometric Analysis of Cucumber (Cucumis sativus L.) Research Publications from Horticulture Category Based on the Web of Science. HortScience 2021, 56, 1304–1314. [Google Scholar] [CrossRef]
- Hanelt, D. Photosynthesis Assessed by Chlorophyll Fluorescence. In Bioassays; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 169–198. ISBN 9780128118610. [Google Scholar]
- Takayama, K.; Konishi, A.; Omasa, K. No Diagnosis of Invisible Photosynthetic Injury Caused by a Herbicide (Basta) with Chlorophyll Fluorescence Imaging System. Agric. Eng. Int. CIGR J. Sci. Res. Dev. 2003, 4, 147–173. [Google Scholar]
- Li, H.; Wang, P.; Weber, J.F.; Gerhards, R. Early Identification of Herbicide Stress in Soybean (Glycine max (L.) Merr.) Using Chlorophyll Fluorescence Imaging Technology. Sensors 2017, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.L.; Tian, Y.; Li, L.; Yu, M.; Hou, R.P.; Ren, X.M. H2S Alleviates Salinity Stress in Cucumber by Maintaining the Na+/K+ Balance and Regulating H2S Metabolism and Oxidative Stress Response. Front. Plant Sci. 2019, 10, 678. [Google Scholar] [CrossRef]
- Estaji, A.; Kalaji, H.M.; Karimi, H.R.; Roosta, H.R.; Moosavi-Nezhad, S.M. How Glycine Betaine Induces Tolerance of Cucumber Plants to Salinity Stress? Photosynthetica 2019, 57, 753–761. [Google Scholar] [CrossRef]
- Sun, L.; Xu, H.; Hao, H.; An, S.; Lu, C.; Wu, R.; Su, W. Effects of Bensulfuron-Methyl Residue on Photosynthesis and Chlorophyll Fluorescence in Leaves of Cucumber Seedlings. PLoS ONE 2019, 14, e0215486. [Google Scholar] [CrossRef]
- Wang, L.; Qin, Z.; Li, X.; Yang, J.; Xin, M. Persistence Behavior of Chlorpyrifos and Biological Toxicity Mechanism to Cucumbers under Greenhouse Conditions. Ecotoxicol. Environ. Saf. 2022, 242, 113894. [Google Scholar] [CrossRef]
- Dunan, L.; Malanga, T.; Bearez, P.; Benhamou, S.; Monticelli, L.S.; Desneux, N.; Michel, T.; Lavoir, A.-V. Biopesticide Evaluation from Lab to Greenhouse Scale of Essential Oils Used against Macrosiphum Euphorbiae. Agriculture 2021, 11, 867. [Google Scholar] [CrossRef]
- Matsui, K.; Ujita, C.; Fujimoto, S.; Wilkinson, J.; Hiatt, B.; Knauf, V.; Kajiwara, T.; Feussner, I. Fatty Acid 9- and 13-Hydroperoxide Lyases from Cucumber1. FEBS Lett. 2000, 481, 183–188. [Google Scholar] [CrossRef]
- Hao, L.; Chen, S.; Wang, C.; Chen, Q.; Wan, X.; Shen, X.; Cheng, Z.; Meng, H. Aroma Components and Their Contents in Cucumbers from Different Genotypes. J. Northwest A F Univ. Sci. Ed. 2013, 41, 139–146. [Google Scholar]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and Distribution of Floral Scent. Bot. Rev. 2006, 72, 1. [Google Scholar] [CrossRef]
- Zhang, J.; Gu, X.; Yan, W.; Lou, L.; Xu, X.; Chen, X. Characterization of Differences in the Composition and Content of Volatile Compounds in Cucumber Fruit. Foods 2022, 11, 1101. [Google Scholar] [CrossRef]
- Ramos, S.E.; Bakhtiari, M.; Castañeda-Zárate, M.; Iriart, V.; Ashman, T.-L. Herbicides and Their Potential to Disrupt Plant–Insect Chemical Communication. J. Syst. Evol. 2022, 1–11. [Google Scholar] [CrossRef]
- Fuchs, B.; Saikkonen, K.; Helander, M. Glyphosate-Modulated Biosynthesis Driving Plant Defense and Species Interactions. Trends Plant Sci. 2021, 26, 312–323. [Google Scholar] [CrossRef]
- Kamou, N.N.; Kalogiouri, N.P.; Tryfon, P.; Papadopoulou, A.; Karamanoli, K.; Dendrinou-Samara, C.; Menkissoglu-Spiroudi, U. Impact of Geraniol and Geraniol Nanoemulsions on Botrytis Cinerea and Effect of Geraniol on Cucumber Plants’ Metabolic Profile Analyzed by LC-QTOF-MS. Plants 2022, 11, 2513. [Google Scholar] [PubMed]
- Chen, S.; Zhang, R.; Hao, L.; Chen, W.; Cheng, S. Profiling of Volatile Compounds and Associated Gene Expression and Enzyme Activity during Fruit Development in Two Cucumber Cultivars. PLoS ONE 2015, 10, e0119444. [Google Scholar] [CrossRef]
- Deng, J.; Yu, H.-J.; Li, Y.-Y.; Zhang, X.-M.; Liu, P.; Li, Q.; Jiang, W.-J. Leaf Volatile Compounds and Associated Gene Expression during Short-Term Nitrogen Deficient Treatments in Cucumis Seedlings. Int. J. Mol. Sci. 2016, 17, 1713. [Google Scholar] [CrossRef]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, I. The Role of Sugars in Plant Responses to Stress and Their Regulatory Function during Development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed]
- Feller, C.; Bleiholder, H.; Buhr, L.; Hack, H.; Hess, M.; Klose, R.; Meier, U.; Stauss, R.; Boom, T.; van den Weber, E. Phanologische Entwicklungsstadien von Gemusepflanzen II. Fruchtgemuse Und Hulsenfruchte. Nachr. Dtsch. Pflanzenschutzd. 1995, 47, 217–232. [Google Scholar]
- Athar, H.-R.; Ambreen, S.; Javed, M.; Hina, M.; Rasul, S.; Zafar, Z.U.; Manzoor, H.; Ogbaga, C.C.; Afzal, M.; Al-Qurainy, F.; et al. Influence of Sub-Lethal Crude Oil Concentration on Growth, Water Relations and Photosynthetic Capacity of Maize (Zea mays L.) Plants. Environ. Sci. Pollut. Res. 2016, 23, 18320–18331. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Taylor and Francis: London, UK; New York, NY, USA, 2000; pp. 445–483. [Google Scholar]
- Nakurte, I.; Berga, M.; Pastare, L.; Kienkas, L.; Senkovs, M.; Boroduskis, M.; Ramata-Stunda, A. Valorization of Bioactive Compounds from By-Products of Matricaria Recutita White Ray Florets. Plants 2023, 12, 396. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Package “Factoextra” for R: Extract and Visualize the Results of Multivariate Data Analyses; R Package Version; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 2012, 1, 726. [Google Scholar]
- Marichali, A.; Hosni, K.; Dallali, S.; Ouerghemmi, S.; Ltaief, H.B.H.; Benzarti, S.; Kerkeni, A.; Sebei, H. Allelopathic Effects of Carum carvi L. Essential Oil on Germination and Seedling Growth of Wheat, Maize, Flax and Canary Grass. Allelopath. J. 2014, 34, 81. [Google Scholar]
- Campos, E.V.R.; de Oliveira, J.L.; Pascoli, M.; de Lima, R.; Fraceto, L.F. Neem Oil and Crop Protection: From Now to the Future. Front. Plant Sci. 2016, 7, 1494. [Google Scholar] [CrossRef]
- Nebo, L.; Varela, R.M.; Molinillo, J.M.G.; Severino, V.G.P.; Sarria, A.L.F.; Cazal, C.M.; das Fernandes, M.F.G.; Fernandes, J.B.; Macías, F.A. Phytotoxicity of Triterpenes and Limonoids from the Rutaceae and Meliaceae. 5α,6β,8α,12α-Tetrahydro-28-Norisotoonafolin--a Potent Phytotoxin from Toona Ciliata. Nat. Prod. Commun. 2015, 10, 17–20. [Google Scholar] [CrossRef]
- Khanam, H.; Shazadur Rahman, M.; Jahidul Islam, M.; Fancy, R.; Belal Hosain, M.; Shamima Nasrin Rima, R. Allelopathic Effect of Neem (Azadirachta Indica) Leaf Extracts on Germination and Seedling Growth of Some Vegetable Crops. Asian J. Res. Bot. 2020, 4, 113–121. [Google Scholar]
- Gomes, M.P.; Smedbol, E.; Chalifour, A.; Hénault-Ethier, L.; Labrecque, M.; Lepage, L.; Lucotte, M.; Juneau, P. Alteration of Plant Physiology by Glyphosate and Its By-Product Aminomethylphosphonic Acid: An Overview. J. Exp. Bot. 2014, 65, 4691–4703. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Fayez, K.A. Photosynthesis, Antioxidant Status and Gas-Exchange Are Altered by Glyphosate Application in Peanut Leaves. Photosynthetica 2016, 54, 307–316. [Google Scholar] [CrossRef]
- Jankevica, L.; Polis, O.; Korica, A.; Samsone, I.; Laugale, V.; Daugavietis, M. Environmental Risk Assessment Studies on New Plant Protection Products Which Have Been Elaborated from Coniferous Tree Bark. Agron. Res. 2018, 16, 2056–2067. [Google Scholar] [CrossRef]
- Luo, Y.; Hu, T.; Huo, Y.; Wang, L.; Zhang, L.; Yan, R. Effects of Exogenous Melatonin on Chrysanthemum Physiological Characteristics and Photosynthesis under Drought Stress. Horticulturae 2023, 9, 106. [Google Scholar] [CrossRef]
- Nasrollahi, P.; Razavi, S.M.; Ghasemian, A.; Zahri, S. Physiological and Biochemical Responses of Lettuce to Thymol, as Allelochemical. Russ. J. Plant Physiol. 2018, 65, 598–603. [Google Scholar] [CrossRef]
- Ye, J.; Hua, S.; Liu, S.; Tian, F.; Ji, X.; Li, Y.; Hou, M.; Xu, W.; Meng, L.; Sun, L. Enantioselective Effects of Chiral Fragrance Carvone (L- and D-Carvone) on the Physiology, Oxidative Damage, Synthesis, and Release of Microcystin-LR in Microcystis Aeruginosa. Sci. Total Environ. 2022, 853, 158631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tan, Y.; McClements, D.J. Investigate the Adverse Effects of Foliarly Applied Antimicrobial Nanoemulsion (Carvacrol) on Spinach. LWT 2021, 141, 110936. [Google Scholar] [CrossRef]
- Zobiole, L.H.S.; de Oliveira, R.S.; Kremer, R.J.; Constantin, J.; Bonato, C.M.; Muniz, A.S. Water Use Efficiency and Photosynthesis of Glyphosate-Resistant Soybean as Affected by Glyphosate. Pestic. Biochem. Physiol. 2010, 97, 182–193. [Google Scholar] [CrossRef]
- Yanniccari, M.; Tambussi, E.; Istilart, C.; Castro, A.M. Glyphosate Effects on Gas Exchange and Chlorophyll Fluorescence Responses of Two Lolium perenne L. Biotypes with Differential Herbicide Sensitivity. Plant Physiol. Biochem. 2012, 57, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Naranjo, E.; Redondo-Gómez, S.; Cox, L.; Cornejo, J.; Figueroa, M.E. Effectiveness of Glyphosate and Imazamox on the Control of the Invasive Cordgrass Spartina Densiflora. Ecotoxicol. Environ. Saf. 2009, 72, 1694–1700. [Google Scholar] [CrossRef]
- Midzi, J.; Jeffery, D.W.; Baumann, U.; Rogiers, S.; Tyerman, S.D.; Pagay, V. Stress-Induced Volatile Emissions and Signalling in Inter-Plant Communication. Plants 2022, 11, 2566. [Google Scholar] [CrossRef]
- Li, X.; Sun, Y.; Wang, X.; Dong, X.; Zhang, T.; Yang, Y.; Chen, S. Relationship between Key Environmental Factors and Profiling of Volatile Compounds during Cucumber Fruit Development under Protected Cultivation. Food Chem. 2019, 290, 308–315. [Google Scholar] [CrossRef]
- Zhou, Q.; Cheng, X.; Wang, S.; Liu, S.; Wei, C. Effects of Chemical Insecticide Imidacloprid on the Release of C6 Green Leaf Volatiles in Tea Plants (Camellia sinensis). Sci. Rep. 2019, 9, 625. [Google Scholar] [CrossRef]
- Li, Y.; Wu, C.; Wu, T.; Wang, L.; Chen, S.; Ding, T.; Hu, Y. Preparation and Characterization of Citrus Essential Oils Loaded in Chitosan Microcapsules by Using Different Emulsifiers. J. Food Eng. 2018, 217, 108–114. [Google Scholar] [CrossRef]
- Arbid, Y.; Sleiman, M.; Richard, C. Photochemical Interactions between Pesticides and Plant Volatiles. Sci. Total Environ. 2022, 807, 150716. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Tonny, S.H.; Afzal, S.; Farooqui, Z.; Alam, P.; Ahmed, S.M.; Yu, F.; Hayat, S. β-Cyclocitral: Emerging Bioactive Compound in Plants. Molecules 2022, 27, 6845. [Google Scholar] [CrossRef] [PubMed]
- Gondor, O.K.; Pál, M.; Janda, T.; Szalai, G. The Role of Methyl Salicylate in Plant Growth under Stress Conditions. J. Plant Physiol. 2022, 277, 153809. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Extract Concentration, mg mL−1 | Chemical Composition | ||
|---|---|---|---|---|
| Carvone, % | Limonene, % | Other Compounds, % | ||
| E1 | 1.26 | 77.42% | 21.98% | 0.60% |
| E2 | 2.37 | 97.66% | 0% | 2.34% |
| E3 | 1.87 | 98.88% | 0% | 1.12% |
| E4 | 2.55 | 98.62% | 0% | 1.38% |
| Abbreviation | Measurement Specification |
|---|---|
| F0 | Minimal fluorescence with all PSII reaction centers (RC) assumed to be open |
| Fv | Maximal variable fluorescence |
| FM | Maximal fluorescence, when all PSII RCs are closed |
| Fm/F0 | Ratio of maximal to minimal fluorescence |
| Fv/F0 | Ratio of variable to minimal fluorescence |
| Fv/FM | Photosynthetic efficiency of dark-adapted reaction centers, i.e., which number of absorbed photons can be converted into electron transport |
| M0 | Approximated initial slope (in ms−1) of fluorescence transient V = f(t) |
| QY | Quantum yield |
| Area | Total complimentary area between the fluorescence induction OJIP curve and Fm line |
| ΦP0 | Maximum quantum yield of primary PS II photochemistry |
| Ψ0 | Probability at t = 0 that a trapped excitation moves on electron into the electron transport chain beyond QA− |
| ΦE0 | Maximum quantum yield of primary PS II photochemistry |
| ΦD0 | Quantum yield of energy dissipation |
| PIABS | Performance index on absorption basis |
| ABS/RC | Absorbed photon flux per reaction center (RC) |
| TR0/RC | Maximum trapped exciton flux per PSII |
| ET0/RC | Electron transport flux from QA to QB per reaction center |
| DI0/RC | Heat dissipation per reaction center |
| Parameter | C2 | C3 | E1 | E2 | E3 | E4 | C1 |
|---|---|---|---|---|---|---|---|
| PH10, cm | 21.67 a | 27.67 a | 68.00 b | 64.67 b | 61.00 b | 67.00 b | 64.33 b |
| PDW, g | 1.76 a | 3.10 b | 4.93 c | 4.60 c | 4.86 c | 6.14 d | 5.32 cd |
| LN | 7.33 a | 9.67 a | 14.33 b | 14.67 b | 13.67 b | 14.33 b | 14.67 b |
| ILN | 1.00 a | 3.00 a | 7.33 b | 7.67 b | 6.67 b | 6.67 b | 7.33 b |
| VA, scores | 1.00 a | 3.50 b | 4.67 c | 4.50 c | 4.33 bc | 4.83 c | 4.50 c |
| No. | Compound | Class | Formula | Relative Amount Range, % |
|---|---|---|---|---|
| 1 | 1-Deoxy-d-mannitol | Sugar alcohols | C6H14O5 | 0–4.1 |
| 2 | Dimethyl sulfide | Sulfides | C2H6S | 6.5–70.6 |
| 3 | Pentanal | Aldehydes | C5H10O | 0–21.1 |
| 4 | 2,4-Hexadienal | Aldehydes | C6H8O | 0–15.7 |
| 5 | Ethyl-1-propenyl ether | Ethers | C5H10O | 0–22.0 |
| 6 | 2,4-Dimethyl-3-hexanol | Alcohols | C8H18O | 0–1.6 |
| 7 | 4-Methylhexan-3-ol | Alcohols | C7H16O | 0–90.6 |
| 8 | 2-Methyl-1,3-butanediol | Alcohols | C5H12O2 | 0–12.0 |
| 9 | Hexanal | Aldehydes | C6H12O | 0–6.2 |
| 10 | 1-Butanol | Alcohols | C4H10O | 0–2.9 |
| 11 | 4-Methyl-1-penten-3-ol | Alcohols | C6H12O | 0–2.7 |
| 12 | 1-Pentanol | Alcohols | C5H12O | 0–1.2 |
| 13 | 4-Ethyl-2-methyl-1H-pyrrole | Pyrroles | C7H11N | 0–2.1 |
| 14 | 2-Hexenal | Aldehydes | C6H10O | 0–2.7 |
| 15 | n-Hexyl acetate | Esters | C8H16O2 | 0–0.7 |
| 16 | 2-[(2E)-2-Pentenyl]furan | Furans | C9H12O | 0–1.5 |
| 17 | (4E)-4-Hexenyl acetate | Esters | C8H14O2 | 0–26.8 |
| 18 | 1-Hexanol | Alcohols | C6H14O | 0–2.0 |
| 19 | Hex-3-en-1-ol | Alcohols | C6H12O | 0–0.8 |
| 20 | cis-3-Hexen-1-ol | Alcohols | C6H12O | 0–23.4 |
| 21 | Methyl salicylate | Esters | C8H8O3 | 0–1.7 |
| 22 | Benzaldehyde | Aldehydes | C7H6O | 0–2.2 |
| 23 | 3-Hexanol | Alcohols | C6H14O | 0–6.2 |
| 24 | β-Cyclocitral | Aldehydes | C10H16O | 0–1.6 |
| 25 | Hexahydrofarnesyl acetone | Sesquiterpenes | C18H360 | 0–4.6 |
| Parameter | C2 | C3 | E1 | E2 | E3 | E4 | C1 |
|---|---|---|---|---|---|---|---|
| TPC a mg (GAE/g) | 8.22 a * | 4.64 b | 6.26 ac | 6.69 bc | 6.97 ac | 6.91 ac | 6.42 c |
| Sugars b, mg (GLE/g) | 32.70 a | 13.97 b | 26.86 c | 19.75 bde | 23.57 acd | 26.53 cde | 17.37 be |
| ARA c, mg (ASE /g) | 6.95 a | 2.86 b | 4.19 ab | 4.34 ab | 4.49 ab | 4.43 ab | 4.80 ab |
| DPPH Quenched, % | 40.67 a | 17.39 b | 25.03 ab | 25.87 ab | 25.74 ab | 26.25 ab | 28.74 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mežaka, I.; Kronberga, A.; Berga, M.; Kaļāne, L.; Pastare, L.; Skudriņš, G.; Nakurte, I. Biochemical and Physiological Responses of Cucumis sativus L. to Application of Potential Bioinsecticides—Aqueous Carum carvi L. Seed Distillation By-Product Based Extracts. Agriculture 2023, 13, 1019. https://doi.org/10.3390/agriculture13051019
Mežaka I, Kronberga A, Berga M, Kaļāne L, Pastare L, Skudriņš G, Nakurte I. Biochemical and Physiological Responses of Cucumis sativus L. to Application of Potential Bioinsecticides—Aqueous Carum carvi L. Seed Distillation By-Product Based Extracts. Agriculture. 2023; 13(5):1019. https://doi.org/10.3390/agriculture13051019
Chicago/Turabian StyleMežaka, Ieva, Arta Kronberga, Marta Berga, Laura Kaļāne, Laura Pastare, Gundars Skudriņš, and Ilva Nakurte. 2023. "Biochemical and Physiological Responses of Cucumis sativus L. to Application of Potential Bioinsecticides—Aqueous Carum carvi L. Seed Distillation By-Product Based Extracts" Agriculture 13, no. 5: 1019. https://doi.org/10.3390/agriculture13051019