
Responses of Micropropagated Rhubarb (Rheum rhaponticum) Plantlets to Different Growing Media and Light Conditions in the Greenhouse
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
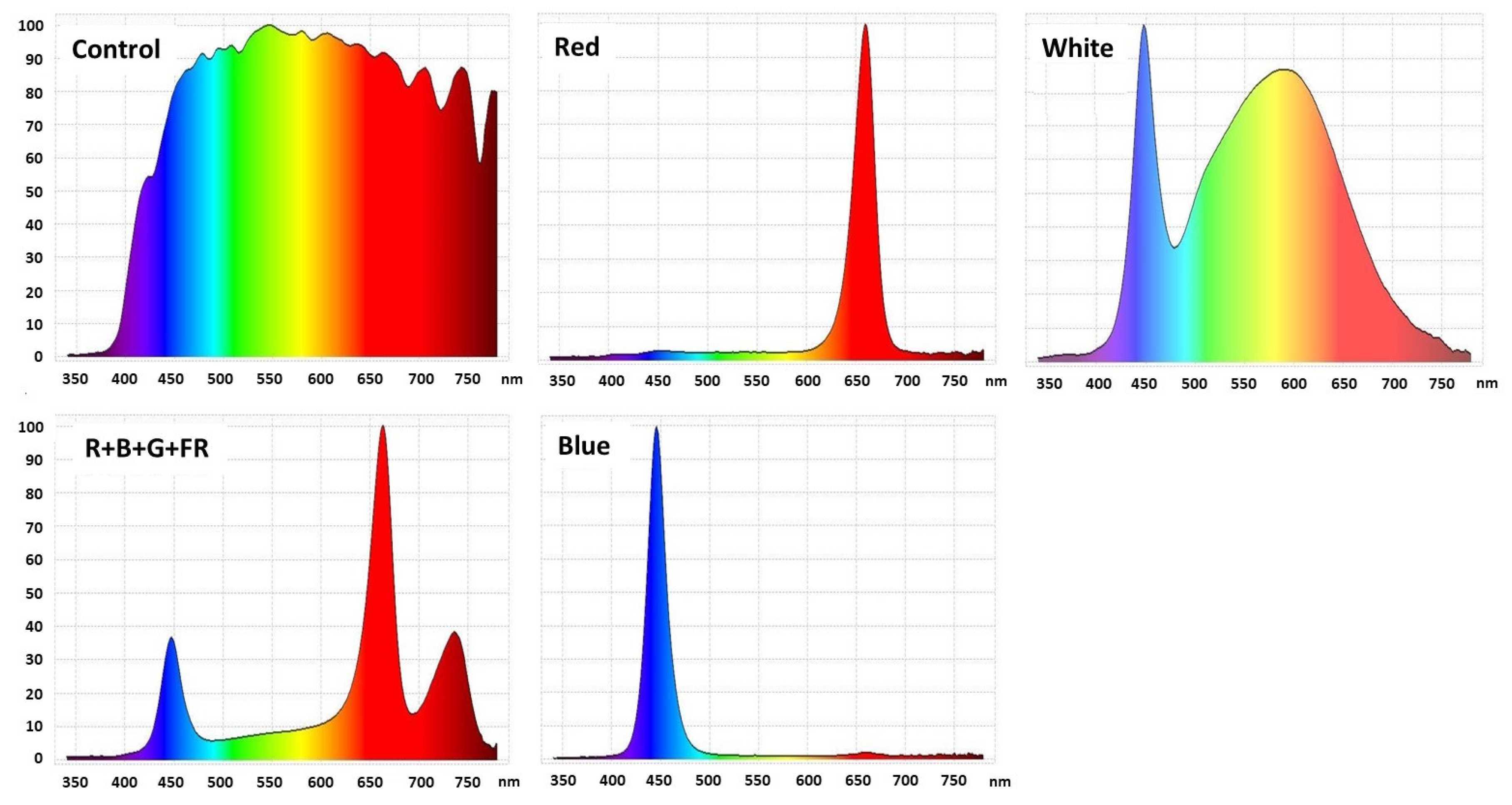
2.2. Light Treatments
2.3. Plant Growth Parameters
2.4. Phenolic Compound Analysis
2.5. Soluble Sugar Analysis
2.6. Antioxidant Activity Measurement
2.7. Statistical Analyses
3. Results and Discussion
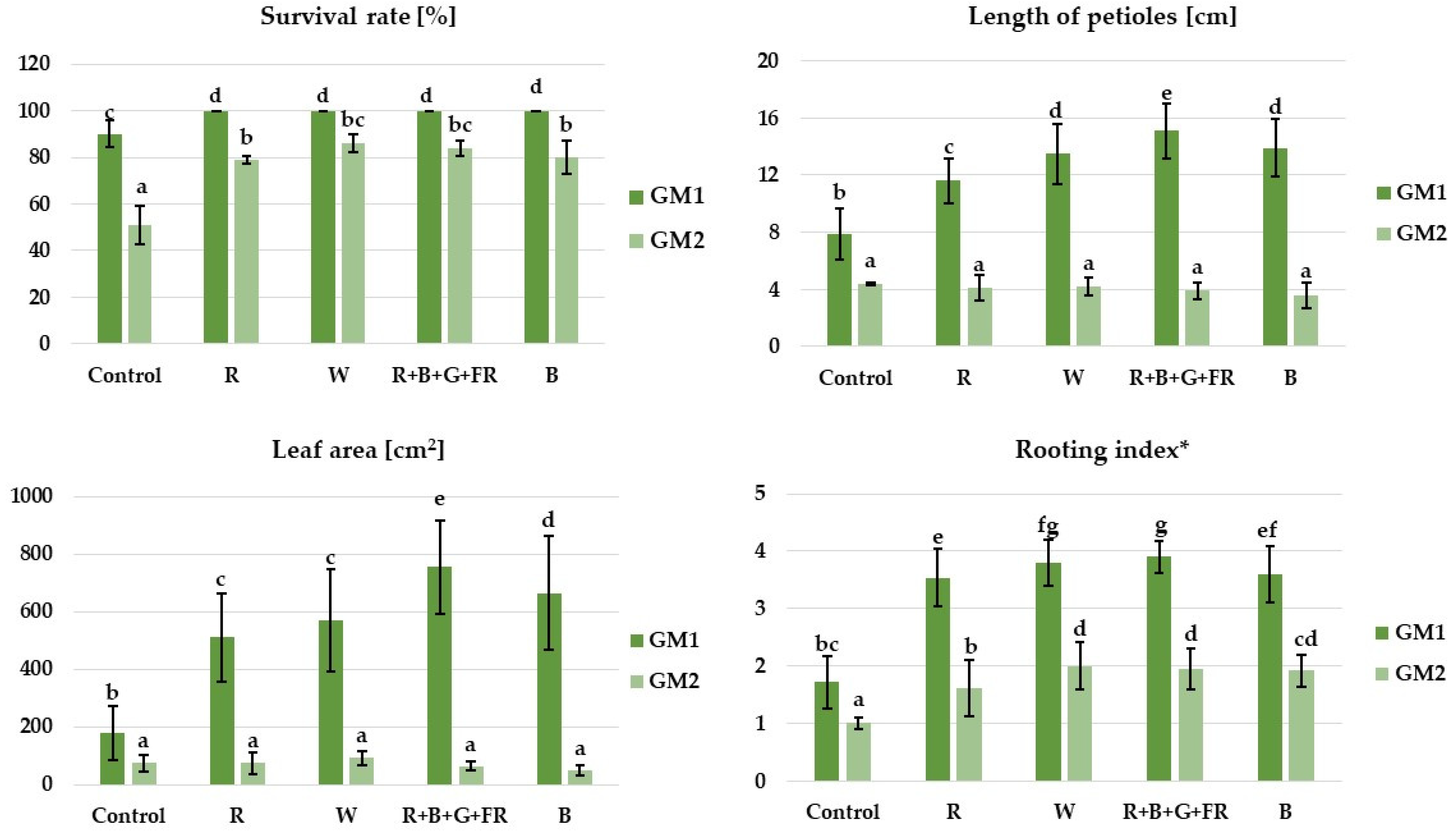
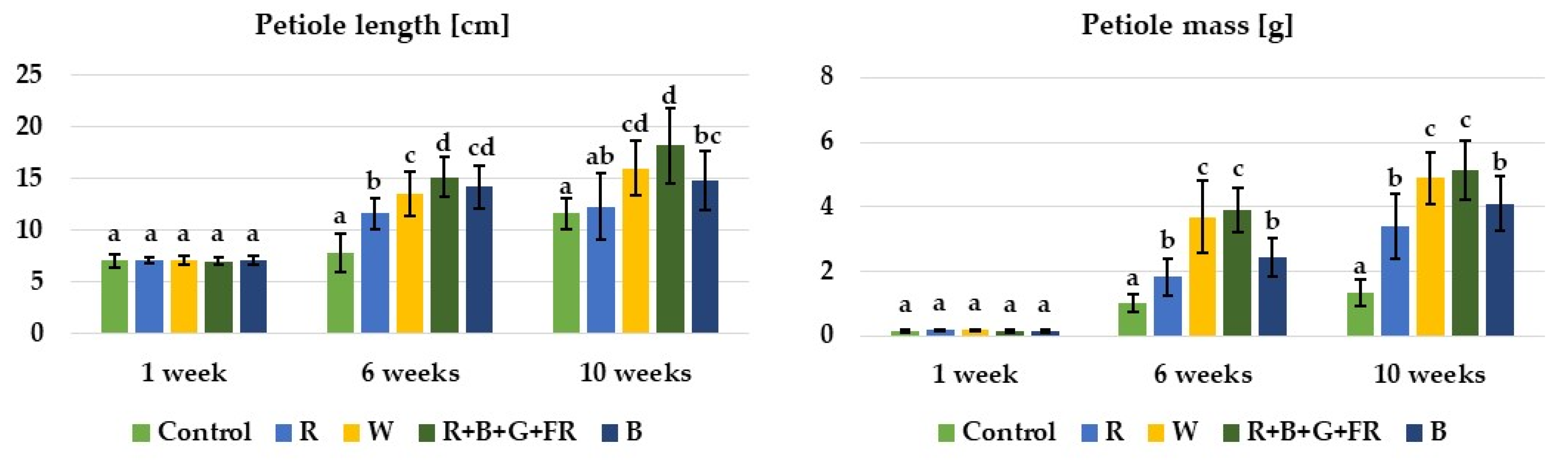
3.1. Growth and Morphological Characteristics of Plants
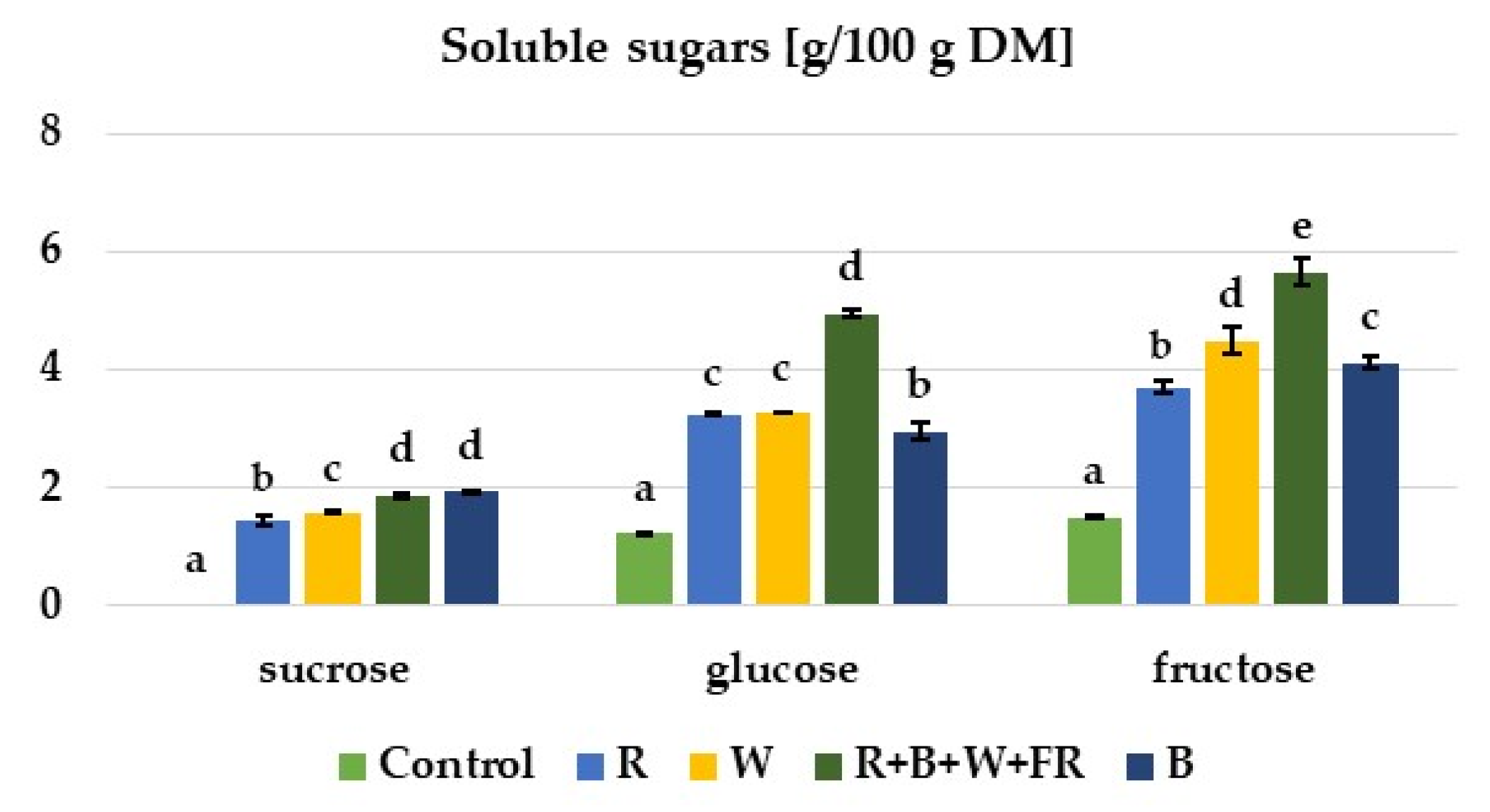
3.2. Soluble Sugars in the Leaf Petioles
3.3. Phenolic Compounds in the Leaf Petioles
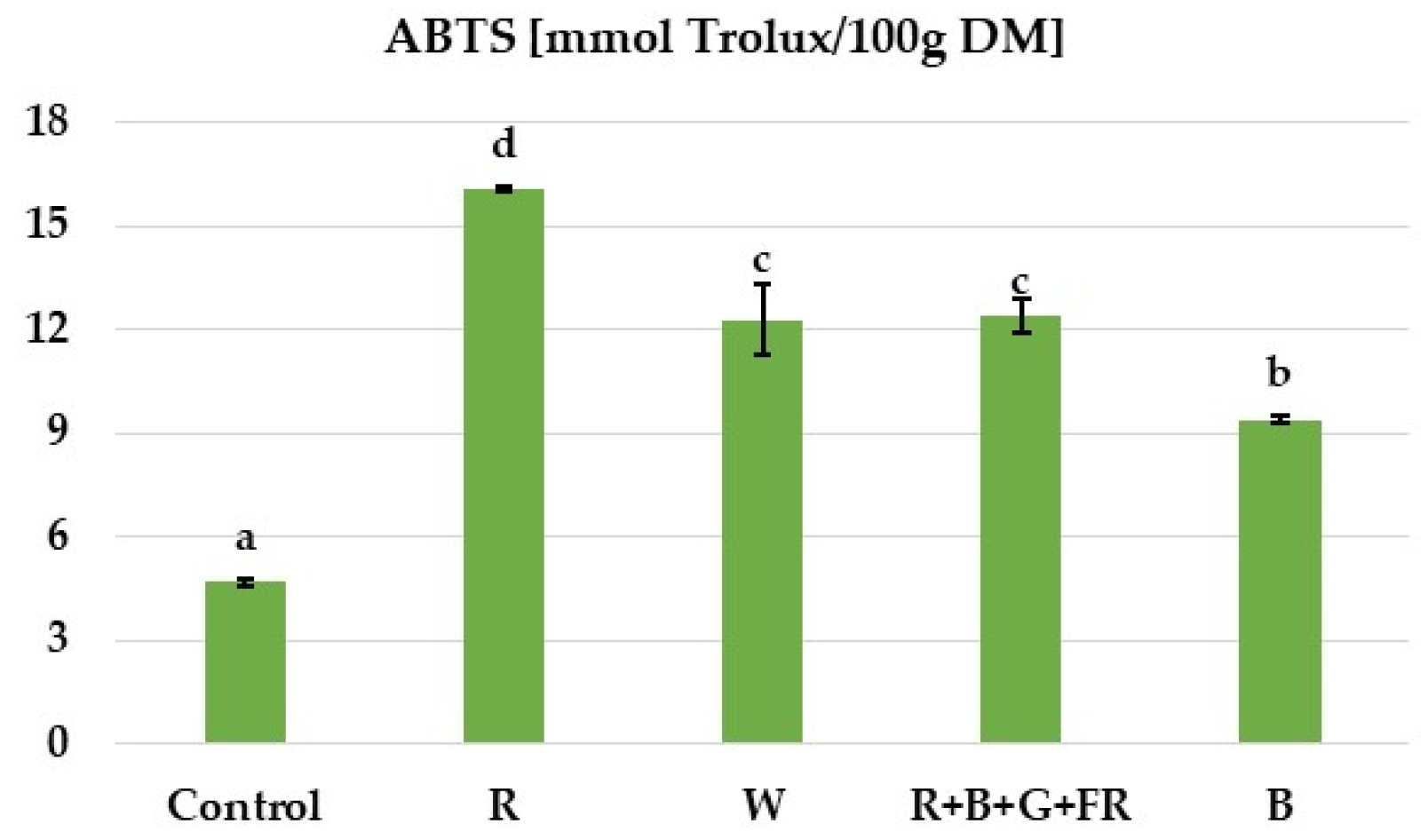
3.4. Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takeoka, G.R.; Dao, L.; Harden, L.; Pantoja, A.; Kuhl, J.C. Antioxidant activity, phenolic and anthocyanin contents of various rhubarb (Rheum spp.) varieties. Int. J. Food Sci. Technol. 2013, 48, 172–178. [Google Scholar] [CrossRef]
- Kołodziejczyk-Czepas, J.; Liudvytska, O. Rheum rhaponticum and Rheum rhabarbarum: A review of phytochemistry, biological activities and therapeutic potential. Phytochem. Rev. 2021, 20, 589–607. [Google Scholar] [CrossRef]
- Bratsch, A.; Mainville, D. Specialty crop profile: Rhubarb. In Virginia Cooperative Extension; Publication 438-110; Rudd, Ed.; VirginiaTech: Blacksburg, VA, USA, 2009; pp. 1–7. [Google Scholar]
- Kuhl, J.C.; DeBoer, V.L. Genetic diversity of rhubarb cultivars. J. Am. Soc. Hortic. Sci. 2008, 133, 587–592. [Google Scholar] [CrossRef]
- Xiang, H.; Zuo, J.; Guo, F.; Dong, D. What we already know about rhubarb: A comprehensive review. Chin. Med. 2020, 15, 88. [Google Scholar] [CrossRef]
- McDougall, G.J.; Dobson, P.; Jordan-Mahy, N. Effect of different cooking regimes on rhubarb polyphenols. Food Chem. 2010, 119, 758–764. [Google Scholar] [CrossRef]
- Kalisz, S.; Oszmiański, J.; Kolniak-Ostek, J.; Grobelna, A.; Kieliszek, M.; Cendrowski, A. Effect of a variety of polyphenols compounds and antioxidant properties of rhubarb (Rheum rhabarbarum). Food Sci. Technol. 2020, 118, 108775. [Google Scholar] [CrossRef]
- Oszmiański, J.; Wojdyło, A. Polyphenol content and antioxidative activity in apple purées with rhubarb juice supplement. Int. J. Food Sci. Technol. 2008, 43, 501–508. [Google Scholar] [CrossRef]
- Wojtania, A.; Mieszczakowska-Frąc, M. In vitro propagation method for production of phenolic-rich planting material of culinary rhubarb ‘Malinowy’. Plants 2021, 10, 1768. [Google Scholar] [CrossRef]
- Podwyszyńska, M.; Pluta, S. In vitro tetraploid induction of the blackcurrant (Ribes nigrum L.) and preliminary phenotypic observations. Zemdirb. Agric. 2019, 106, 151–158. [Google Scholar] [CrossRef]
- Pospíšilová, J.; Tichá, I.; Kadleček, P.; Haisel, D.; Plzáková, Š. Acclimatization of micropropagated plants to ex vitro conditions. Biol. Plant. 1999, 42, 481–497. [Google Scholar] [CrossRef]
- Chandra, S.; Bandopadhyay, R.; Kumar, V.; Chandra, R. Acclimatization of tissue cultured plantlets: From laboratory to land. Biotechnol. Lett. 2010, 32, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Wojtania, A.; Markiewicz, M.; Waligórski, P. Growth cessation and dormancy induction in micropropagated plantlets of Rheum rhaponticum ‘Raspberry’ influenced by photoperiod and temperature. Int. J. Mol. Sci. 2023, 24, 607. [Google Scholar] [CrossRef] [PubMed]
- Galvão, C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Massa, G.D.; Kim, H.H.; Wheeler, M.; Mitchel, C.A. Plant productivity in response to LEDs. HortScience 2008, 43, 1951–1956. [Google Scholar] [CrossRef]
- Viršilė, A.; Olle, M.; Duchovskis, P. LED lighting in horticulture. In Light Emitting Diodes for Agriculture; Gupta, S.D., Ed.; Springer: Singapore, 2017. [Google Scholar]
- Teixeira, R.T. Distinct responses to light in plants. Plants 2020, 9, 894. [Google Scholar] [CrossRef]
- Chung, G.-J.; Lee, J.-H.; Oh, M.-M. Growth and Acclimation of In Vitro-Propagated M9 Apple Rootstock Plantlets under Various Visible Light Spectrums. Agronomy 2020, 10, 1017. [Google Scholar] [CrossRef]
- Tarakanov, I.G.; Tovstyko, D.A.; Lomakin, M.P.; Shmakov, A.S.; Sleptsov, N.N.; Shmarev, A.N.; Litvinskiy, V.A.; Ivlev, A.A. Effects of light spectral quality on photosynthetic activity, biomass production, and carbon isotope fractionation in lettuce, Lactuca sativa L. plants. Plants 2022, 5, 441. [Google Scholar] [CrossRef]
- Fekry, W.A.; Wahdan, H.M. Influence of substrates on in vitro rooting and acclimatization of micropropagated strawberry (Fragaria ×Ananasa Duch.). Middle East J. Agric. 2017, 6, 682–991. [Google Scholar]
- Salgado Pirata, M.; Correia, S.; Canhoto, J. Ex vitro simultaneous acclimatization and rooting of in vitro propagated tamarillo plants (Solanum betaceum Cav.): Effect of the substrate and mineral nutrition. Agronomy 2022, 12, 1082. [Google Scholar] [CrossRef]
- Pandey, R.; Vengavasi, K.; Haekesford, M.J. Plant adaptation to nutrient stress. Plant Physiol. Rep. 2021, 26, 583–586. [Google Scholar] [CrossRef]
- Nielsen, I.L.; Haren, G.R.; Magnussen, E.L.; Dragsted, L.O.; Rasmussen, S.E. Quantification of anthocyanins in commercial black currant juices by simple high-performance liquid chromatography. Investigation of their pH stability and antioxidative potency. J. Agric. Food Chem. 2003, 51, 5861–5866. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Oszmiański, J.; Wojdyło, A. Effects of various clarification treatments on phenolic compounds and color of apple juice. Eur. Food Res. Technol. 2007, 224, 755–762. [Google Scholar] [CrossRef]
- Kipp, J.A.; Wever, G.; de Kreij, C. International Substrate Manual. Analysis, Characteristics and Recommendation; Elsevier: Amsterdam, The Netherlands, 2000; p. 94. [Google Scholar]
- Kitir, N.; Yildirim, E.; Şahin, Ü.; Turan, M.; Ekinci, M.; Ors, S.; Kul, R.; Ünlü, H.; Ünlü, H. Peat Use in Horticulture. In Open Access Peer-Reviewed Chapter in Peat; Topcuoğlu, B., Turan, M., Eds.; IntechOpen: Budapest, Hungary, 2018. [Google Scholar] [CrossRef]
- Wojtania, A.; Markiewicz, M.; Góraj-Koniarska, J. Ex vitro rooting and acclimatization of Lonicera caerulea var. kamtschatica shoots, and determination of genetic stability of plantlets using AFLP and ISSR markers. J. Hortic. Res. 2020, 28, 61–70. [Google Scholar] [CrossRef]
- Matysiak, B.; Gabryszewska, E. The effect of in vitro culture conditions on the pattern of maximum photochemical efficiency of photosystem II during acclimatisation of Helleborus niger plantlets to ex vitro conditions. Plant Cell Tiss. Organ Cult. 2016, 125, 585–593. [Google Scholar] [CrossRef]
- Lakho, M.A.; Jatoi, M.A.; Solangi, N.; Abul-Soad, A.A.; Qazi, M.A.; Abdi, G. Optimizing in vitro nutrient and ex vitro soil medium-driven resposes for multiplication, rooting, and acclimatization of pineapple. Sci. Rep. 2023, 13, 1275. [Google Scholar] [CrossRef]
- Raviv, M.; Wallach, R.; Silber, A.; Bar-Tal, A. Substrates and their analysis. In Hydroponic Production of Vegetables and Ornamentals; Sawas, D., Passam, H., Eds.; Embryo Publications: Athens, Greece, 2002; pp. 25–101. [Google Scholar]
- Wojtania, A.; Markiewicz, M.; Waligórski, P. Regulation of the bud dormancy development and release in micropropagated rhubarb ‘Malinowy’. Int. J. Mol. Sci. 2022, 23, 1480. [Google Scholar] [CrossRef]
- Seo, M.; Nambara, E.; Choi, G.; Yamaguchi, S. Interaction of light and hormone signals in germinating seeds. Plant Mol. Biol. 2009, 69, 463–472. [Google Scholar] [CrossRef]
- Lau, O.S.; Deng, X.W. Plant hormone signaling lightens up: Integrators of light and hormones. Curr. Opin. Plant Biol. 2010, 13, 571–577. [Google Scholar] [CrossRef]
- Kondo, S.; Tomiyama, H.; Rodyoung, A.; Okawa, K.; Ohara, H.; Sugaya, S.; Terahara, N.; Hirai, N. Abscisic acid metabolism and anthocyanin synthesis in grape skin are affected by light emitting diode (LED) irradiation at night. J. Plant Physiol. 2014, 171, 823–829. [Google Scholar] [CrossRef]
- Nozue, K.; Tat, A.V.; Kumar Devisetty, U.; Robinson, M.; Mumbach, M.R.; Ichihashi, Y.; Lekkala, S.; Mallof, J.N. Shade avoidance components and pathways in adult plants revealed by phenotypic profiling. PLOS Genet. 2015, 11, e1004953. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Runkle, E.S. Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemical of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Silvestri, C.; Caceres, M.E.; Ceccarelli, M.; Pica, A.L.; Rugini, E.; Cristofori, V. Influence of continuous spectrum light on morphological traits and leaf anatomy of hazelnut plantlets. Front. Plant Sci. 2019, 10, 1318. [Google Scholar] [CrossRef]
- Ciereszko, I. Regulatory roles of sugars in plant growth and development. Acta Soc. Bot. Pol. 2018, 87, 3583. [Google Scholar] [CrossRef]
- Kula-Maximenko, M.; Niewiadomska, E.; Maksymowicz, A.; Ostrowska, A.; Oklestkova, J.; Pěnčík, A.; Janeczko, A. Insight into details of the photosynthetic light reactions and selected metabolic changes in tomato seedlings growing under various light spectra. Int. J. Mol. Sci. 2021, 22, 11517. [Google Scholar] [CrossRef]
- Yang, L.Y.; Wang, L.T.; Ma, J.H.; Ma, E.D.; Li, J.Y.; Gong, M. Effects of light quality on growth and development, photosynthetic characteristics and content of carbohydrates in tobacco (Nicotiana tabacum L.) plants. Photosynthetica 2017, 55, 467–477. [Google Scholar] [CrossRef]
- Matysiak, B. The Effect of supplementary LED lighting on the morphological and physiological traits of miniature Rosa × hybrida ‘Aga’ and the development of powdery mildew (Podosphaera pannosa) under greenhouse conditions. Plants 2021, 10, 417. [Google Scholar] [CrossRef]
- Sobczak, A.; Sujkowska-Rybkowska, M.; Gajc-Wolska, J.; Kowalczyk, W.; Borucki, W.; Kalaji, H.M.; Kowalczyk, K. Photosynthetic efficiency and anatomical structure of pepper leaf (Capsicum annuum L.) transplants grown under high-pressure sodium (HPS) and light-emitting diode (LED) supplementary lighting systems. Plants 2021, 10, 1975. [Google Scholar] [CrossRef]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signal and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 274–279. [Google Scholar] [CrossRef]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Atanassova, R. The sugar-signaling hub: Overview of regulators and interaction with the hormonal and metabolic network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, F.M.; Visser, R.G.F.; Bovy, A. Antocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Ren, L.; Lian, H.; Liu, Y.; Chen, H.Y. Novel insight into the mechanism underlying light-controlled anthocyanin accumulation in eggplant (Solanum melongena L.). Plant Sci. 2016, 249, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ma, X.; Gao, X.; Wu, W.; Zhou, B. Light induced regulation pathway of anthocyanin biosynthesis in plants. Int. J. Mol. Sci. 2021, 22, 11116. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Cao, S.; Chen, W.; Yang, Z. Blue light induced anthocyanin accumulation and expression of associated genes in Chinese bayberry fruit. Scientia. Horticulturae 2014, 179, 98–102. [Google Scholar] [CrossRef]
- Azad, M.O.K.; Kjaer, K.H.; Adnan, M.; Naznin, M.T.; Lim, J.D.; Sung, I.J.; Park, C.H.; Lim, Y.S. The evaluation of growth performance, photosynthetic capacity, and primary and secondary metabolite content of leaf lettuce grown under limited irradiation of blue and red LED light in an urban plant factory. Agriculture 2020, 10, 28. [Google Scholar] [CrossRef]
- Liu, Y.; Schouten, R.E.; Tikunov, Y.; Liu, X.; Visser, R.G.F.; Tan, F.; Bovy, A.; Marcelis, L.F.M. Blue light increases anthocyanin content and delays fruit ripening in purple pepper fruit. Postharvest Biol. Technol. 2022, 192, 11202. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Sirtautas, R.; Sakalauskienė, S.; Jankauskienė, J.; Duchovskis, P.; Novičkovas, A. The impact of supplementary short-term red LED lighting on the antioxidant properties of microgreens. Acta Hortic. 2012, 956, 649–656. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Jankauskienė, J.; Novičkovas, A. The effects of supplementary short-term red LEDs lighting on nutritional quality of Perilla frutescens L. microgreens. Rural Dev. Proc. 2013, 6, 54–58. [Google Scholar]
- Lekkham, P.; Srilaong, V.; Pongprasert, N.; Kondo, S. Anthocyanin concentration and antioxidant activity in light-emitting diode (LED)-treated apples in a greenhouse environmental control system. Fruits 2016, 71, 269–274. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 13, 2452. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light. Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef] [PubMed]
- Samuolienė, G.; Brazaitytė, A.; Sirtautas, R.; Viršilė, A.; Sakalauskaitė, J.; Sakalauskienė, S. LED illumination affects bioactive compounds in romaine baby leaf lettuce. J. Sci. Food Agric. 2013, 93, 3286–3291. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, M.-H. Ultraviolet a-specific induction of anthocyanin biosynthesis and PAL expression in tomato (Solanum lycopersicum L.). Plant Growth Regul. 2010, 62, 1–8. [Google Scholar] [CrossRef]
- Xu, Z.; Mahmood, K.; Rothstein, S.J. ROS induces anthocyanin production via late biosynthetic genes and anthocyanin deficiency confers the hypersensitivity to ROS-generating stresses in Arabidopsis. Plant Cell Physiol. 2017, 58, 1364–1377. [Google Scholar] [CrossRef]
- Naig, A.H.; Kim, C.K. Abiotic stress-induced anthocyanins in plants: Their role in tolerance to abiotic stresses. Physiol. Plant. 2021, 172, 1711–1723. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Floragard Substrate (GM1) | Neutralised Sphagnum Peat (GM2) |
|---|---|---|
| pH (H2O; 1:2) | 6.2 | 6.2 |
| EC (mS cm−1) | 1.14 | 0.47 |
| N-NO3 (mg·dm−3) | 93.0 | 19.0 |
| N-NH4 (mg·dm−3) | 87.5 | <1.0 |
| P-PO4 (mg·dm−3) | 85.0 | 39.0 |
| K (mg·dm−3) | 193.0 | 68.0 |
| Mg (mg·dm−3) | 111.0 | 111.0 |
| Ca (mg·dm−3) | 1151.0 | 1915.0 |
| Na (mg·dm−3) | 10.0 | 10.0 |
| Cl− (mg·dm−3) | 38.0 | 44.0 |
| S-SO4 (mg·dm−3) | 84.0 | 27.0 |
| Fe (mg·dm−3) | 8.05 | 10.1 |
| Mn(mg·dm−3) | 4.7 | 2.24 |
| Cu (mg·dm−3) | 1.2 | 0.83 |
| Zn (mg·dm−3) | 2.2 | 1.9 |
| Organic matter (%) | 91.9 | 90.0 |
| Total pore space (%) | 94.7 | 95.3 |
| Bulk density (g·cm−3) | 85.7 | 76.7 |
| Water volume at −10 cm H2O (%) | 68.3 | 60.0 |
| Air volume at −10 cm H2O (%) | 26.4 | 35.3 |
| LED Light Treatments | UV-A 340–400 nm | Blue 400–500 nm | Green-Yellow 500–600 nm | Red 600–700 nm | Far-Red 700–780 nm | R:FR Ratio |
|---|---|---|---|---|---|---|
| Control | 3.9 | 22.1 | 28.0 | 27.4 | 16.8 | 1.6 |
| R-Red | - | - | - | 100.0 | - | - |
| W-White | 0.1 | 24.4 | 44.4 | 28.9 | 2.2 | 11.3 |
| R+B+G+FR | 0.2 | 16.3 | 10.3 | 49.4 | 23.8 | 2.1 |
| B-Blue | - | 100.0 | - | - | - | - |
| Trait/Interaction | Leaf Petiole Length | Leaf Area [cm2] | Rooting Index | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Light | 45.8 | <0.000 *** | 37.0 | <0.000 *** | 134.2 | <0.000 *** |
| Substrate | 1352.0 | <0.000 *** | 845.1 | <0.000 *** | 998.0 | <0.000 *** |
| Light × Substrate | 27.2 | <0.000 *** | 42.2 | <0.000 *** | 20.3 | <0.000 *** |
| Phenolic Compounds [mg/100 g DM] | Light Treatments | ||||
|---|---|---|---|---|---|
| Control | Red | White | R+B+G+FR | Blue | |
| Anthocyanins: | 216.4 ± 0.79 a | 561.0 ± 6.3 d | 585.0 ± 0.69 e | 374.1 ± 0.42 b | 435.4 ± 4.0 c |
| Cyanidin-3-O-rutinoside | 200.8 ± 6.3 a | 488.4 ± 6.7 d | 503.2 ± 3.4 e | 332.2 ± 2.3 b | 387.7 ± 3.6 c |
| Cyanidin-3-O-glucoside | 10.8 ± 0.11 a | 59.9 ± 0.17 d | 68.8 ± 0.63 e | 33.0 ± 0.11 b | 38.4 ± 0.17 c |
| Delphinidin-3-O-rutonoside | 4.8 ± 0.04 a | 12.7 ± 0.25 c | 13.0 ± 0.28 c | 8.9 ± 0.08 b | 9.2 ± 0.22 b |
| Flavonols | 3.1 ± 0.17 a | 4.4 ± 0.20 d | 4.1 ± 0.13 c | 3.6 ± 0.02 b | 3.4 ± 0.03 b |
| Flavan-3-ols (Catechin) | 12.7 ± 1.3 a | 32.3 ± 0.57 c | 30.8 ± 0.03 c | 36.5 ± 0.60 d | 23.4 ± 0.06 b |
| Phenolic acids | 1.9 ± 0.03 a | 4.1 ± 0.09 d | 3.0 ± 0.05 c | 2.1 ± 0.12 b | 2.1 ± 0.05 b |
| Total phenolic compounds | 234.0 ± 19.2 a | 601.8 ± 6.8 d | 622.8 ± 0.58 e | 416.4 ± 11.8 b | 464.4 ± 3.9 c |
| Petiole Mass | Petiole Length | Leaf Area | Sugars | Anthocyanins | Phenolic Compounds | |
|---|---|---|---|---|---|---|
| Petiole length | 0.800 | |||||
| Leaf area | 0.872 | 0.666 | ||||
| Sugars | 0.891 | 0.718 | 0.934 | |||
| Anthocyanins | 0.605 | 0.195 | 0.520 | 0.528 | ||
| Phenolic compounds | 0.625 | 0.219 | 0.543 | 0.560 | 0.999 | |
| ABTS | 0.607 | 0.257 | 0.616 | 0.708 | 0.832 | 0.852 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtania, A.; Matysiak, B.; Mieszczakowska-Frąc, M.; Nowak, J.S.; Szwejda-Grzybowska, J. Responses of Micropropagated Rhubarb (Rheum rhaponticum) Plantlets to Different Growing Media and Light Conditions in the Greenhouse. Agriculture 2023, 13, 890. https://doi.org/10.3390/agriculture13040890
Wojtania A, Matysiak B, Mieszczakowska-Frąc M, Nowak JS, Szwejda-Grzybowska J. Responses of Micropropagated Rhubarb (Rheum rhaponticum) Plantlets to Different Growing Media and Light Conditions in the Greenhouse. Agriculture. 2023; 13(4):890. https://doi.org/10.3390/agriculture13040890
Chicago/Turabian StyleWojtania, Agnieszka, Bożena Matysiak, Monika Mieszczakowska-Frąc, Jacek S. Nowak, and Justyna Szwejda-Grzybowska. 2023. "Responses of Micropropagated Rhubarb (Rheum rhaponticum) Plantlets to Different Growing Media and Light Conditions in the Greenhouse" Agriculture 13, no. 4: 890. https://doi.org/10.3390/agriculture13040890