
Biostimulants as a Response to the Negative Impact of Agricultural Chemicals on Vegetation Indices and Yield of Common Buckwheat (Fagopyrum esculentum Moench)

Abstract
:1. Introduction
2. Materials and Research Methods
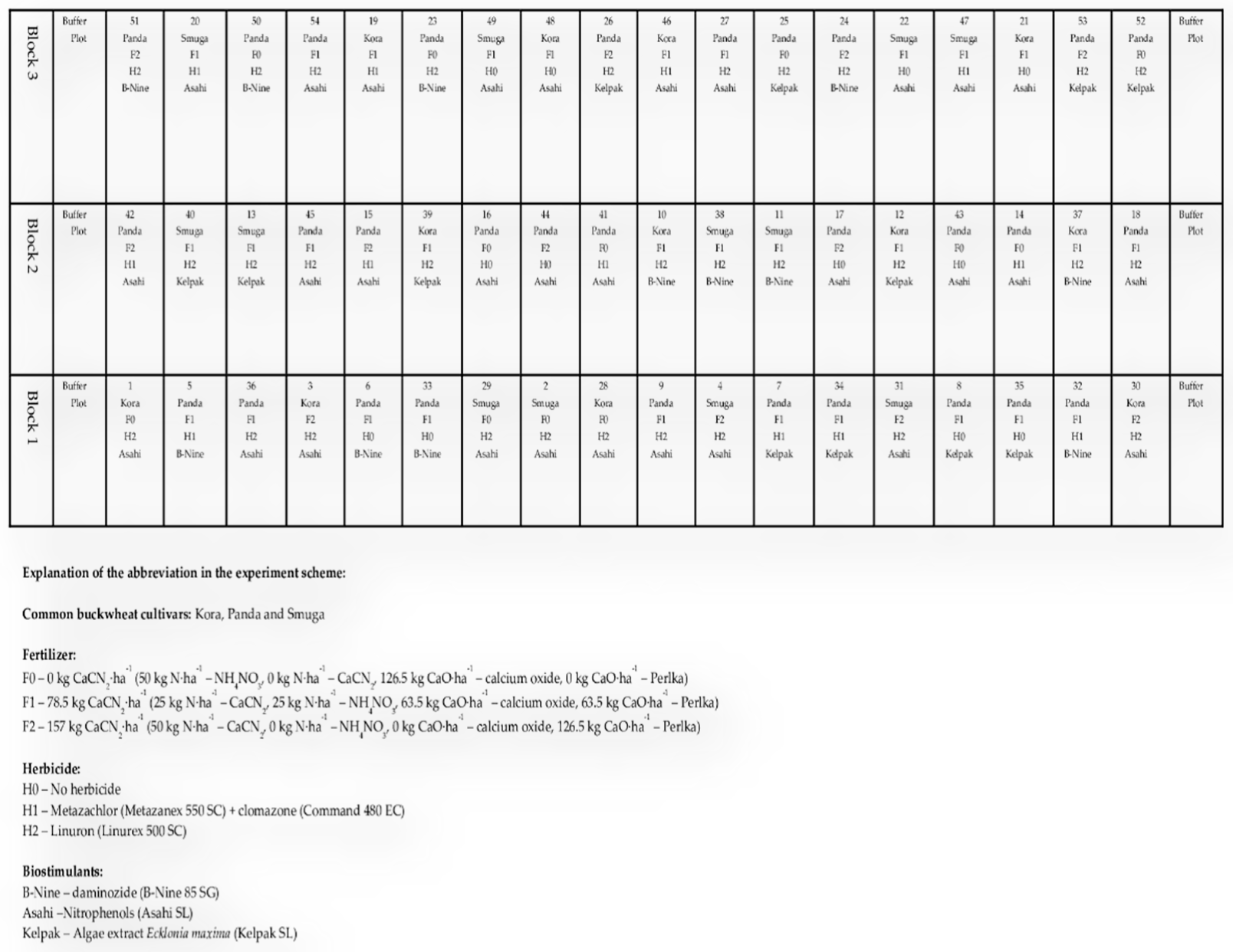
2.1. Plant Material and Field Experiment
2.2. Vegetation Indices
2.3. Soil Conditions
2.4. Weather Conditions
2.5. Statistical Analyses
3. Results
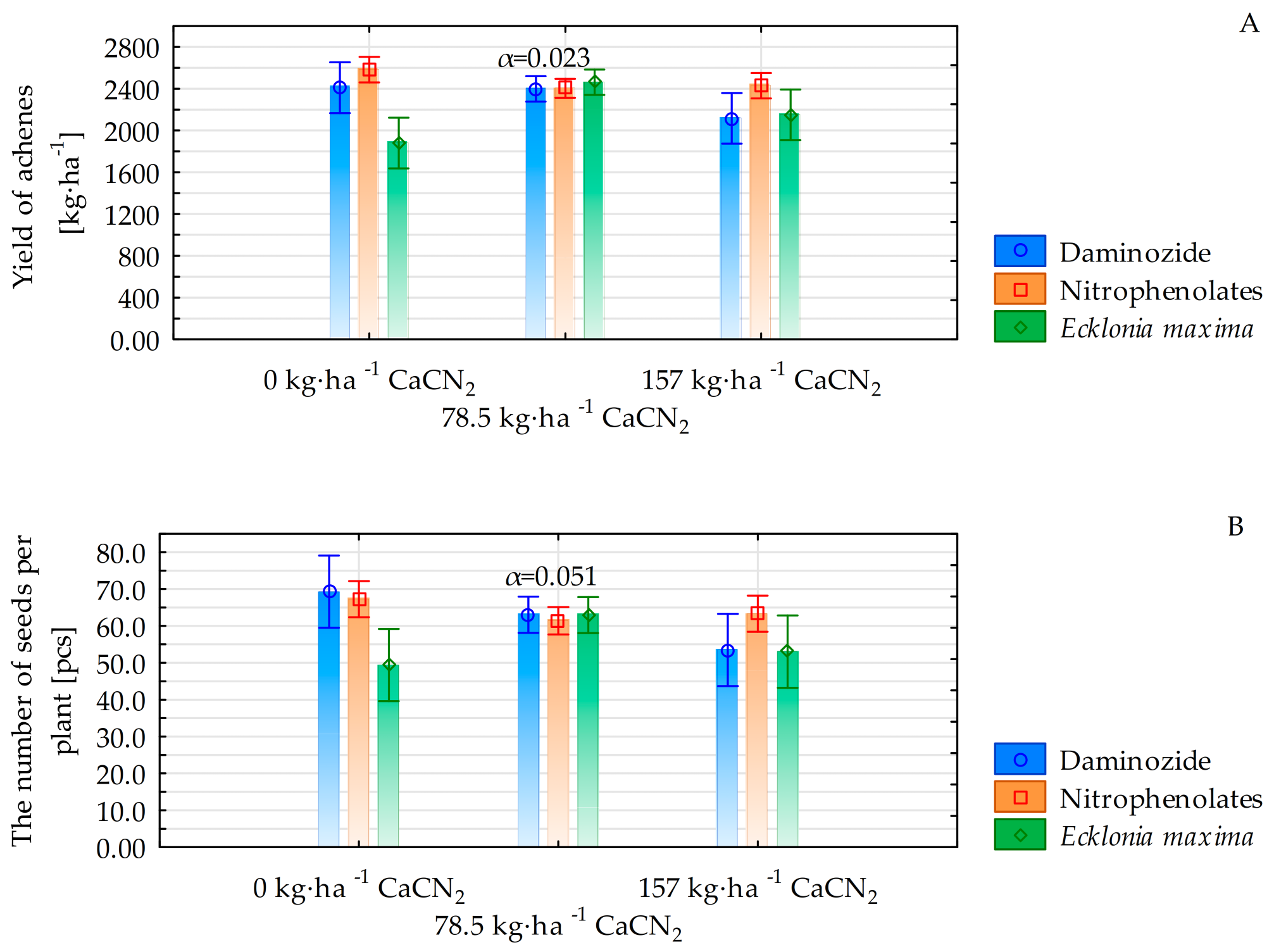
3.1. Yield and Yield Components
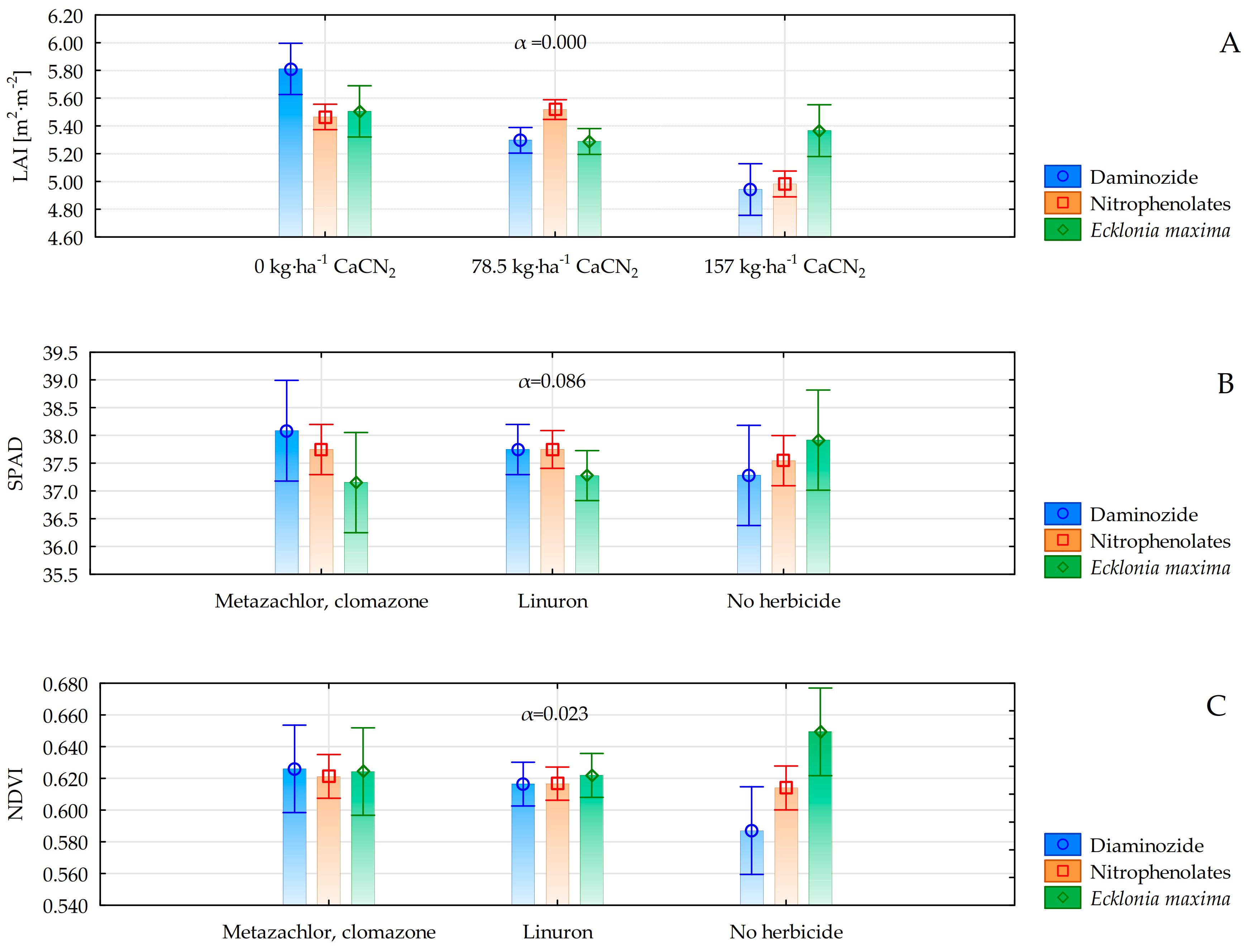
3.2. Vegetation Indices
3.3. The Segetal Flora
4. Discussion
4.1. Yield and Yield Components of Common Buckwheat
4.2. Buckwheat Vegetation Indices
4.3. The Segetal Flora of the Buckwheat Field
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Sturm, D.J.; Peteinatos, G.; Gerhards, R. Contribution of allelopathic effects to the overall weed suppression by different cover crops. Weed Res. 2018, 58, 331–337. [Google Scholar] [CrossRef]
- Wall, D.A.; Smith, M.A.H. Weed management in common buckwheat (Fagopyrum esculentum). Can. J. Plant Sci. 1999, 79, 455–461. [Google Scholar] [CrossRef]
- Sakaliene, O.; Clay, S.A.; Koskinen, W.C.; Almantas, G. Early Season Weed Suppression in Buckwheat Using Clopyralid. Weed Technol. 2008, 22, 707–712. [Google Scholar] [CrossRef] [Green Version]
- Podolska, G.; Górecka, D.; Russel, H.; Dziedzic, K.; Boguszewska, E. Abiotic stress affects the yield and nutrients of buckwheat grains. Zemdirbyste 2019, 106, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Debski, H.; Wiczkowski, W.; Szawara-Nowak, D.; Baczek, N.; Chrzanowski, G.; Horbowicz, M. Effects of glyphosate and fluazifop-P-butyl on flavonoids content and growth of common buckwheat (Fagopyrum esculentum Moench). Fresenius Environ. Bull. 2018, 27, 91–97. [Google Scholar]
- Podolska, G. Efektywność i fitotoksyczność herbicydów w zasiewach gryki zwyczajnej odmiany Kora. Pol. J. Agron. 2014, 19, 17–24. (In Polish) [Google Scholar]
- Dixon, G.R. Calcium cyanamide—A synoptic review of an environmentally benign fertiliser which enhances soil health. Acta Hortic. 2012, 938, 211–217. [Google Scholar] [CrossRef]
- Amberger, A. Cyanamide in plant metabolism. Int. J. Plant Physiol. Biochem. 2013, 5, 1–10. [Google Scholar]
- Koch, W.; Hurle, K. Grundlgen der Unkrtautbekäpfung; Verlag Eugen Ulmer: Stuttgart, Germany, 1978; p. 205. (In German) [Google Scholar]
- Börner, H. Unkrtautbekäpfung; Gustav Fisher Verlag: Jena, Germany, 1995; p. 315. (In German) [Google Scholar]
- Dobrzański, A. Rabarbar bez chwastów. Warzywa 2015, 7, 43–46. (In Polish) [Google Scholar]
- Dixon, G.R. Managing clubroot disease caused by Plasmodiophora brassicae Wor. by exploiting the interactions between calcium cyanamide fertilizer and soil microorganisms. J. Agric. Sci. 2017, 155, 527–543. [Google Scholar] [CrossRef]
- Shi, K.; Wang, L.; Zhou, V.H.; Yu, Y.L.; Yu, J.Q. Effects of calcium cyanamide on soil microbial communities and Fusarium oxysporum f. sp. Cucumberinum. Chemosphere 2009, 75, 872–877. [Google Scholar] [CrossRef]
- Lu, G.L.; Wei, W.Y.; Li, G.P.; Xia, M.; Yang, G.F.; Zhao, Z.Y. Observation on long––term field mulluscidal of Rongbao. Pract. Prev. Med. 2006, 13, 1462–1465. [Google Scholar]
- Wei, W.; Zhu, H.; Liu, Z.; Lv, G.; Ding, L.; Gu, J. Effect of calcium cyanamide on killing eggs of Schistodoma japonicum and maggots. Chin. J. Schistosomiasis Control 2009, 21, 127–129. [Google Scholar]
- Nõmmik, H. On Decomposition of Calcium Cyanamide and Dicyanamide in the Soil. Acta Agric. Scand. 1958, 8, 404–440. [Google Scholar] [CrossRef]
- Estermaier, L.M.; Sieber, A.H.; Lottspeich, F.; Matern, D.H.M.; Hartmann, G.R. Biochemical Degradation of Cyanamide and Dicyandiamide. Angew. Chem. Int. Ed. Engl. 1992, 31, 620–622. [Google Scholar] [CrossRef]
- Amoo, A.E.; Babalola, O.O. Ammonia-oxidizing microorganisms: Key players in the promotion of plant growth. J. Soil Sci. Plant Nutr. 2017, 17, 935–947. [Google Scholar] [CrossRef] [Green Version]
- Guardia, G.; Vallejo, A.; Cardenas, L.M.; Dixon, E.R.; García-Marco, S. Fate of 15N-labelled ammonium nitrate with or without the new nitrification inhibitor DMPSA in an irrigated maize crop. Soil Biol. Biochem. 2018, 116, 193–202. [Google Scholar] [CrossRef]
- du Jardin, P.; The Science of Plant Biostimulants—A Bibliographic Analysis. Ad hoc Study Report to the European Commission DG ENTR. 2012. Available online: http://ec.europa.eu/enterprise/sectors/chemicals/files/fertilizers/final_report_bio_2012_en.pdf (accessed on 12 October 2019).
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 396. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant-and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Tekindal, M.A.; Bayrak, H.; Ozkaya, B.; Genc, Y. Box-Behnken experimental design in factorial experiments: The importance of bread for nutrition and health. Turk. J. Field Crops 2012, 17, 115–123. [Google Scholar]
- Gundogdu, T.K.; Deniz, I.; Caliskan, G.; Sahin, E.S.; Azbar, N. Experimental design methods for bioengineering applications. Crit. Rev. Biotechnol. 2014, 34, 1–21. [Google Scholar]
- Peng, X.; Yang, G.; Shi, Y.; Zhou, Y.; Zhang, M.; Li, S. Box-Behnken design based statistical modelling for the extractic and physicochemical properties of pectin from sunflower heads and the comparison with commercial low-metoxyl pectin. Sci. Rep. 2020, 10, 3595. [Google Scholar]
- Harrison, H.F.; Farnham, M.W.; Michael Jackson, D. Tolerance of broccoli cultivars to pre-transplanting clomazone. Crop Prot. 2015, 69, 28–33. [Google Scholar] [CrossRef]
- Karier, P.; Kraus, G.; Kolber, I. Metazachlor traces in the main drinking water reservoir in Luxembourg: A scientific and political discussion. Environ. Sci. Eur. 2017, 29, 25. [Google Scholar] [CrossRef] [Green Version]
- Witkowicz, R.; Skrzypek, E.; Gleń-Karolczyk, K.; Krupa, M.; Biel, W.; Chłopicka, J.; Galanty, A. Effects of application of plant growth promoters, biological control agents and microbial soil additives on photosynthetic efficiency, canopy vegetation indices and yield of common buckwheat (Fagopyrum esculentum Moench). Biol. Agric. Hortic. 2021, 37, 234–251. [Google Scholar] [CrossRef]
- Craige, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- ISO 520; 2010 Specifies a Method for the Determination of the Mass of 1000 Grains of Cereals and Pulses. ISO: Geneva, Switzerland, 2022.
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources 2014. Update 2015. World Soil Resources Reports; FAO: Rome, Italy, 2015; p. 1006. ISBN 978-92-5-108370-3. [Google Scholar]
- ISO 10390; Soil Quality—Determination of pH. ISO: Geneva, Switzerland, 2005. Available online: https://www.iso.org/standard/40879.html (accessed on 8 September 2022).
- ISO 11261; Soil Quality—Determination of Total Nitrogen—Modified Kjeldahl Method. ISO: Geneva, Switzerland, 1995. Available online: https://www.iso.org/standard/19239.html (accessed on 8 September 2022).
- Mebius, L.J. A rapid method for the determination of organic carbon in soil. Anal. Chim. Acta 1960, 22, 120–124. [Google Scholar] [CrossRef]
- Selyaninov, G.T. About climate agricultural estimation. Proc. Agric. Meteorol. 1928, 20, 165–177. (In Russian) [Google Scholar]
- Liu, L.W.; Lu, C.T.; Wang, Y.M.; Lin, K.H.; Ma, X.; Lin, W.S. Rice (Oryza sativa L.) Growth Modeling Based on Growth Degree Day (GDD) and Artificial Intelligence Algorithms. Agriculture 2022, 12, 59. [Google Scholar] [CrossRef]
- Jung, G.H.; Kim, S.L.; Kim, M.J.; Kim, S.K.; Park, J.H.; Kim, C.G.; Heu, S. Effect of sowing time on buckwheat (Fagopyrum esculentum Moench) growth and yield in Central Korea. J. Crop Sci. Biotech. 2015, 4, 285–291. [Google Scholar] [CrossRef]
- Tibco. 2020. Available online: https://community.tibco.com/wiki/tibco-statistica-design-experiments#toc-5 (accessed on 17 September 2022).
- Witkowicz, R.; Biel, W.; Skrzypek, E.; Chłopicka, J.; Gleń-Karolczyk, K.; Krupa, M.; Prochownik, E.; Galanty, A. Microorganisms and biostimulants impact on the antioxidant activity of buckwheat (Fagopyrum esculentum Moench) sprouts. Antioxidants 2020, 9, 584. [Google Scholar] [CrossRef]
- Small, E. 54. Buckwheat—The World’s Most Biodiversity-Friendly Crop? Biodiversity 2017, 18, 108–123. [Google Scholar] [CrossRef]
- Luthar, Z.; Fabjan, P.; Mlinarič, K. Biotechnological Methods for Buckwheat Breeding. Plants 2021, 10, 1547. [Google Scholar] [CrossRef]
- Luthar, Z.; Germ, M.; Likar, M.; Golob, A.; Vogel-Mikuš, K.; Pongrac, P.; Kušar, A.; Pravst, I.; Kreft, I. Breeding buckwheat for increased levels of rutin, quercetin and other bioactive compounds with potential antiviral effects. Plants 2020, 9, 1638. [Google Scholar] [CrossRef]
- Zielińska, D.; Turemko, M.; Kwiatkowski, J.; Zieliński, H. Evaluation of flavonoid contents and antioxidant capacity of the aerial parts of common and tartary buckwheat plants. Molecules 2012, 17, 9668–9682. [Google Scholar] [CrossRef]
- Fang, X.; Liu, X.; Zhang, Y.; Huang, K.; Zhang, Y.; Li, Y.; Nie, J.; She, H.; Ruan, R.; Yuan, X.; et al. Effects of uniconazole or gibberellic acid application on the lignin metabolism in relation to lodging resistance of culm in common buckwheat (Fagopyrum esculentum M.). J. Agron. Crop Sci. 2018, 204, 414–423. [Google Scholar] [CrossRef]
- Amelin, A.V.; Fesenko, A.N.; Chekalin, E.I.; Fesenko, I.N.; Zaikin, V.V. Higher yielding varieties of common buckwheat (Fagopyrum esculentum Moench) with determinate growth habit (single mutation det) manifest higher photosynthesis rate at stage of grain filling. Acta Agric. Slov. 2020, 115, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Bavec, F.; Pusnik, S.; Rajcan, I. Yield performance of two buckwheat genotypes grown as a full-season and stubble-crop. Rostl. Vyrob. 2002, 48, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; D’Anna, F.; Prinzivalli, C.; Iapichino, G. Soil solarization and calcium cyanamide affect plant vigor, yield, nutritional traits, and nutraceutical compounds of strawberry grown in a protected cultivation system. Agronomy 2019, 9, 513. [Google Scholar] [CrossRef] [Green Version]
- Bletsos, F.A. Grafting and calcium cyanamide as alternatives to methyl bromide for greenhouse eggplant production. Sci. Hortic. 2006, 107, 325–331. [Google Scholar] [CrossRef]
- Tremblay, N.; Bélec, C.; Coulombe, J.; Godin, C. Evaluation of calcium cyanamide and liming for control of clubroot disease in cauliflower. Crop Prot. 2005, 24, 798–803. [Google Scholar] [CrossRef]
- Sabatino, L.; Caracciolo, G.; Consentino, B.B.; Prinzivalli, C.; D’Anna, F. Effects of plant type and calcium cyanamide dosage on yield, nutritional and nutraceutical traits of strawberry in Sicily. Acta Hortic. 2021, 1309, 979–986. [Google Scholar] [CrossRef]
- Di Gioia, F.; Gonnella, M.; Buono, V.; Ayala, O.; Cacchiarelli, J.; Santamaria, P. Calcium cyanamide effects on nitrogen use efficiency, yield, nitrate and dry matter content of lettuce. Agron. J. 2017, 109, 354–362. [Google Scholar] [CrossRef]
- Kaushal, T.; Suganuma, T.; Fujikake, H.; Ohtake, N.; Sueyoshi, K.; Takahashi, Y.; Ohyama, T. Effect of deep placement of N fertilizers and diffrent inoculation methods of Bradyrhizobia on growth, N2 fixation activity and N adsorption rate of field–grown soybean plants. J. Agron. Crop Sci. 2004, 190, 46–58. [Google Scholar]
- Hasanuddin; Hafsah, S.; Marliah, A.; Jumini; Wansurianika; Idawanni; Aryani, D.S. The effect of mixing two herbicides pendimethalin and sulfentrazone on characteristics of soybean yield. IOP Conf. Ser. Earth Environ. Sci. 2021, 644, 012067. [Google Scholar] [CrossRef]
- Karkanis, A.C.; Kontopoulou, C.K.; Lykas, C.; Kakabouki, I.; Petropoulos, S.A.; Bilalis, D. Efficacy and selectivity of pre– and post–emergence herbicides in chia (Salvia hispanica L.) under mediterranean semi–arid conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Salachna, P.; Grzeszczuk, M.; Meller, E.; Soból, M. Oligo-Alginate with low molecular mass improves growth and physiological activity of Eucomis autumnalis under salinity stress. Molecules 2018, 23, 812. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Patel, J.S. Seaweed extract: Biostimulator of plant defense and plant productivity. Int. J. Environ. Sci. Technol. 2020, 17, 553–558. [Google Scholar] [CrossRef]
- Elansary, H.O.; Mahmoud, E.A.; El-Ansary, D.O.; Mattar, M.A. Effects of water stress and modern biostimulants on growth and quality characteristics of mint. Agronomy 2020, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Efthimiadou, A.; Katsenios, N.; Chanioti, S.; Giannoglou, M.; Djordjevic, N.; Katsaros, G. Effect of foliar and soil application of plant growth promoting bacteria on growth, physiology, yield and seed quality of maize under Mediterranean conditions. Sci. Rep. 2020, 10, 21060. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Cellini, A.; Donati, I.; Pastore, C.; Onofrietti, C.; Spinelli, F. Facing climate change: Application of microbial biostimulants to mitigate stress in horticultural crops. Agronomy 2020, 10, 794. [Google Scholar] [CrossRef]
- González-Morales, S.; Solís-Gaona, S.; Valdés-Caballero, M.V.; Juárez-Maldonado, A.; Loredo–Treviño, A.; Benavides-Mendoza, A. Transcriptomics of Biostimulation of Plants Under Abiotic Stress. Front. Genet. 2021, 12, 583888. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant biostimulants: Importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress—A review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Kocira, A.; Lamorska, J.; Kornas, R.; Nowosad, N.; Tomaszewska, M.; Leszczyńska, D.; Kozłowicz, K.; Tabor, S. Changes in Biochemistry and Yield in Response to Biostimulants Applied in Bean (Phaseolus vulgaris L.). Agronomy 2020, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Maach, M.; Boudouasar, K.; Akodad, M.; Skalli, A.; Moumen, A.; Baghour, M. Application of biostimulants improves yield and fruit quality in tomato. Int. J. Veg. Sci. 2021, 27, 288–293. [Google Scholar] [CrossRef]
- Kazaz, S.; Askin, M.A.; Kilic, S.; Ersoy, N. Effects of day length and daminozide on the flowering, some quality parameters and chlorophyll content of Chrysanthemum mofifolium Ramat. Sci. Res. Essays 2010, 5, 3281–3288. [Google Scholar]
- Rose, N.R.; Woon, E.C.Y.; Tumber, A.; Walport, L.J.; Chowdhury, R.; Li, X.S.; King, O.N.F.; Lejeune, C.; Ng, S.S.; Krojer, T.; et al. Plant growth regulator daminozide is a selective inhibitor of human KDM2/7 histone demethylases. J. Med. Chem. 2012, 55, 6639–6643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemabadi, D. The effect of cycocel and daminozide on some growth and flowering characteristics of Calendula officinalis L., an ornamental and medicinal plant. J. Med. Plants Res. 2012, 6, 1752–1757. [Google Scholar] [CrossRef]
- McGraw, D.; Bostian, B. Effect of Herbicides and weeds on yield of “EXCEL” sweetpotato. HortScience 2019, 28, 261–277. [Google Scholar] [CrossRef]
- Kanissery, R.G.; Sims, G.K. Biostimulation for the Enhanced Degradation of Herbicides in Soil. Appl. Environ. Soil Sci. 2011, 2011, 843450. [Google Scholar] [CrossRef] [Green Version]
- Rangjaroen, C.; Lumyong, S.; Sloan, W.T.; Sungthong, R. Herbicide-Tolerant endophytic bacteria of rice plants as the biopriming agents for fertility recovery and disease suppression of unhealthy rice seeds. BMC Plant Biol. 2019, 19, 580. [Google Scholar] [CrossRef] [Green Version]
- Balabanova, D.A.; Paunov, M.; Goltsev, V.; Cuypers, A.; Vangronsveld, J.; Vassilev, A. Photosynthetic performance of the imidazolinone resistant sunflower exposed to single and combined treatment by the herbicide Imazamox and an amino acid extract. Front. Plant Sci. 2016, 7, 1559. [Google Scholar] [CrossRef] [Green Version]
- Neshev, N.; Balabanova–Ivanovska, D.; Yanev, M.; Mitkov, A.; Tonev, T. Recovering effect of biostimulant application on pumpkins (Cucurbita moschata Duchesne ex Poir.) treated with imazamox. Acta Hortic. 2021, 1320, 267–274. [Google Scholar] [CrossRef]
- Sadeh, Y.; Zhu, X.; Dunkerley, D.; Walker, J.P.; Zhang, Y.; Rozenstein, O.; Manivasagam, V.S.; Chenu, K. Fusion of Sentinel–2 and PlanetScope time–series data into daily 3 m surface reflectance and wheat LAI monitoring. Int. J. Appl. Earth Obs. Geoinf. 2021, 96, 102260. [Google Scholar] [CrossRef]
- Novelli, F.; Vuolo, F. Assimilation of sentinel-2 leaf area index data into a physically-based crop growth model for yield estimation. Agronomy 2019, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Munier, S.; Carrer, D.; Planque, C.; Camacho, F.; Albergel, C.; Calvet, J.C. Satellite Leaf Area Index: Global scale analysis of the tendencies per vegetation type over the last 17 years. Remote Sens. 2018, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Sytar, O.; Bruckova, K.; Kovar, M.; Zivcak, M.; Hemmerich, I.; Brestic, M. Nondestructive detection and biochemical quantification of buckwheat leaves using visible (VIS) and near–infrared (NIR) hyperspectral reflectance imaging. J. Cent. Europ. Agricul. 2017, 18, 864–878. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.R.; Singh, S.; Aggarwal, N.; Kushwah, A.; Kumar, S. Inherent variability among different lentil (Lens culinaris Medik.) genotypes against tolerance to metribuzin herbicide. Biochem. Cell. Arch. 2017, 17, 49–56. [Google Scholar]
- Leytur, M.; Vera, A.; Sala, A.; Giardina, B.; Benedetto, A. Pot growing Media Amendment with Calcium Cyanamide and Weed Control Relationships. Int. J. Plant Soil Sci. 2018, 23, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chohura, P.; Kołota, E. Suitability of some nitrogen fertilizers for the cultivation of early cabbage. J. Elem. 2014, 19, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Zellweger, H.; Feichtinger, G.; Pérez, E.; Walter, A.; Hund, A. Unkrautbekämpfung beim Buchweizen: Vergleich von fünf ausgewählten Herbiziden. Agrar. Schweiz 2017, 8, 7–8. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Treatment | |||
|---|---|---|---|---|
| Level of Experimental Factor | ||||
| I | Cultivar | Kora | Panda | MHR Smuga |
| II | Fertilizer | 0 kg CaCN2·ha−1 (50 kg N·ha−1–NH4NO3, 0 kg N·ha−1–CaCN2, 126.5 kg CaO·ha−1–calcium oxide, 0 kg CaO·ha−1–Perlka) BBCH 00 F0 | 78.5 kg CaCN2·ha−1 (25 kg N·ha−1–CaCN2, 25 kg N·ha−1–NH4NO3, 63.5 kg CaO·ha−1–calcium oxide, 63.5 kg CaO·ha−1–Perlka) BBCH 00 F1 | 157 kg CaCN2·ha−1 (50 kg N·ha−1–CaCN2, 0 kg N·ha−1–NH4NO3, 0 kg CaO·ha−1–calcium oxide, 126.5 kg CaO·ha−1–Perlka) BBCH 00 F2 |
| III | Herbicide | No herbicide | Linuron (Linurex 500 SC) 400 g·ha−1 BBCH 02 | Metazachlor (Metazanex 550 SC) 750 g·ha−1 and clomazone (Command 480 EC) 96 g·ha−1 BBCH 02 |
| IV | Biostimulants | Daminozide (Bi-Nine 85 SG) BBCH 14 | Nitrophenols (Asahi SL) BBCH 14 | Seaweed Ecklonia maxima extract (Kelpak SL) BBCH 14 |
| Month | Sum of Temperature [°C] | Rainfall [mm] | Difference between Rainfall and Rainfall over Many Year [mm] # | Hydrothermal Coefficient of Selyaninov * | Characteristic of the Month | Growing Degree Days [°D] |
|---|---|---|---|---|---|---|
| 2016 | ||||||
| May | 448.4 | 41.4 | −35.3 | 0.92 | Dry | 0–712 |
| June | 564.9 | 59.8 | −22.2 | 1.06 | Dry enough | |
| July | 566.5 | 92.8 | −3.4 | 1.64 | Moderately wet | |
| August | 573.0 | 62.0 | −9.8 | 1.08 | Quite dry | |
| 2017 | ||||||
| May | 433.2 | 83.8 | 7.2 | 1.93 | Moderate wet | 0–718 |
| June | 563.0 | 45.2 | −36.8 | 0.80 | Dry | |
| July | 593.9 | 84.4 | −11.8 | 1.42 | Optimal | |
| August | 627.8 | 83.8 | 12.0 | 1.33 | Optimal | |
| 2018 | ||||||
| May | 541.2 | 62.4 | −14.3 | 1.15 | Dry enough | 0–710 |
| June | 554.8 | 85.6 | 3.6 | 1.54 | Optimal | |
| July | 615.5 | 119.8 | 23.6 | 1.95 | Moderately wet | |
| August | 646.0 | 56.2 | −15.6 | 0.87 | Dry | |
| Parameter | Year * | ||
|---|---|---|---|
| 2016 | 2017 | 2018 | |
| Yield of the achenes [kg·ha−1] | 3177 b **A *** | 1917 B | 2113 aA |
| Plant density after emergence [pcs·m−2] | 190 aA | 201 A | 218 bA |
| Plant density before harvesting [pcs·m−2] | 161 aA | 155 B | 186 bA |
| The number of seeds per plant [pcs] | 76.9 bA | 58.7 B | 51.2 aA |
| Thousand-seed weight [g] | 24.4 aA | 23.3 B | 25.1 bA |
| Parameter | Cultivar * | Fertilization * | Herbicide * | PGP * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kora | Panda | Smuga | F0 # | F1 | F2 | No herbicide | Linuron | Metazachlor and Clomazone | Daminozide | Nitrophenols | Ecklonia maxima Extract | |
| Yield of the achenes [kg·ha−1] | 2465 a **A *** | 2269 B | 2672 bA | 2436 aA | 2418 A | 2330 aA | 2403 aA | 2403 A | 2401 aA | 2353 aA | 2313 A | 2458 aA |
| Plant density after emergence [pcs·m−2] | 206 aA | 197 B | 213 aA | 207 aA | 203 A | 199 aA | 209 aA | 199 B | 205 aA | 199 aA | 204 A | 200 aA |
| Plant density before harvesting [pcs·m−2] | 175 aA | 163 A | 170 aA | 163 aA | 169 A | 167 aA | 172 aA | 167 A | 162 aA | 163 aA | 169 A | 168 aA |
| The number of seeds per plant [pcs] | 62.9 aA | 60.6 A | 65.9 aA | 64.6 aA | 62.3 A | 60.0 aA | 59.9 aA | 62.3 A | 64.5 aA | 62.5 aA | 59.0 A | 63.5 aA |
| Thousand-seed weight [g] | 23.6 aA | 24.2 A | 25.0 bA | 24.2 aA | 24.2 A | 24.4 aA | 24.2 aA | 24.4 A | 24.0 aA | 24.8 bA | 24.2 B | 24.1 aA |
| Parameter | Year * | Plant Growth Stage * | ||||
|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | BBCH 20-29 | BBCH 50-59 | BBCH 60-69 | |
| LAI (m2·m−2) | 4.49 b **A *** | 5.87 B | 5.69 aA | 2.44 aA | 6.05 B | 7.55 bA |
| SPAD | 39.47 bA | 35.94 B | 37.48 aA | 40.45 b | - | 34.80 a |
| NDVI | 0.559 aA | 0.646 B | 0.651 aA | 0.605 aA | 0.661 B | 0.589 bA |
| Parameters | Cultivar * | Fertilization * | Herbicide * | PGP * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kora | Panda | Smuga | F0 # | F1 | F2 | No herbicide | Linuron | Metazachlor and Clomazone | Daminozide | Nitrophenols | Ecklonia maxima Extract | |
| LAI (m2·m−2) | 5.45 a **A *** | 5.28 B | 5.41 aA | 5.53 bA | 5.40 B | 5.04 aA | 5.43 bA | 5.36 A | 5.23 aA | 5.32 aA | 5.36 A | 5.34 aA |
| SPAD | 38.68 bA | 37.48 A | 36.94 aA | 38.19 bA | 37.65 aA | 37.00 aA | 37.56 aA | 37.62 A | 37.70 aA | 37.72 aA | 37.69 aA | 37.36 aA |
| NDVI | 0.633 bA | 0.618 A | 0.606 aA | 0.627 bA | 0.620 A | 0.605 aA | 0.615 aA | 0.618 A | 0.623 aA | 0.613 aA | 0.617 A | 0.627 aA |
| Parameters | Year | Cultivar * | Fertilization * | Herbicide * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | Kora | Panda | Smuga | F0 # | F1 | F2 | Control (No Herbicide) | Linuron | Metazachlor and Clomazone | |
| Number of monocotyledonous weeds [pcs·m−2] | 11.86 b **A *** | 3.87 B | 2.75 aA | 5.67 aA | 6.93 B | 4.93 aA | 6.54 aA | 6.19 A | 5.71 aA | 8.43 bA | 7.27 B | 1.43 aA |
| Number of dicotyledonous weeds [pcs·m−2] | 3.34 bA | 2.67 A | 1.26 aA | 2.38 aA | 2.51 A | 2.25 aA | 2.97 A | 2.44 A | 1.82 aA | 2.15 bA | 3.06 B | 1.28 aA |
| Dry weight of weeds [g·m−2] | - | 2.03 b | 0.97 a | 1.34 aA | 1.72 B | 1.18 A | 1.74 bA | 1.44 A | 1.39 aA | 1.73 bA | 1.58 A | 1.10 aA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krupa, M.; Witkowicz, R. Biostimulants as a Response to the Negative Impact of Agricultural Chemicals on Vegetation Indices and Yield of Common Buckwheat (Fagopyrum esculentum Moench). Agriculture 2023, 13, 825. https://doi.org/10.3390/agriculture13040825
Krupa M, Witkowicz R. Biostimulants as a Response to the Negative Impact of Agricultural Chemicals on Vegetation Indices and Yield of Common Buckwheat (Fagopyrum esculentum Moench). Agriculture. 2023; 13(4):825. https://doi.org/10.3390/agriculture13040825
Chicago/Turabian StyleKrupa, Mateusz, and Robert Witkowicz. 2023. "Biostimulants as a Response to the Negative Impact of Agricultural Chemicals on Vegetation Indices and Yield of Common Buckwheat (Fagopyrum esculentum Moench)" Agriculture 13, no. 4: 825. https://doi.org/10.3390/agriculture13040825