
The Impact of Auxin and Cytokinin on the Growth and Development of Selected Crops


Abstract
:1. Development of the Research on Plant Growth and Development Regulators
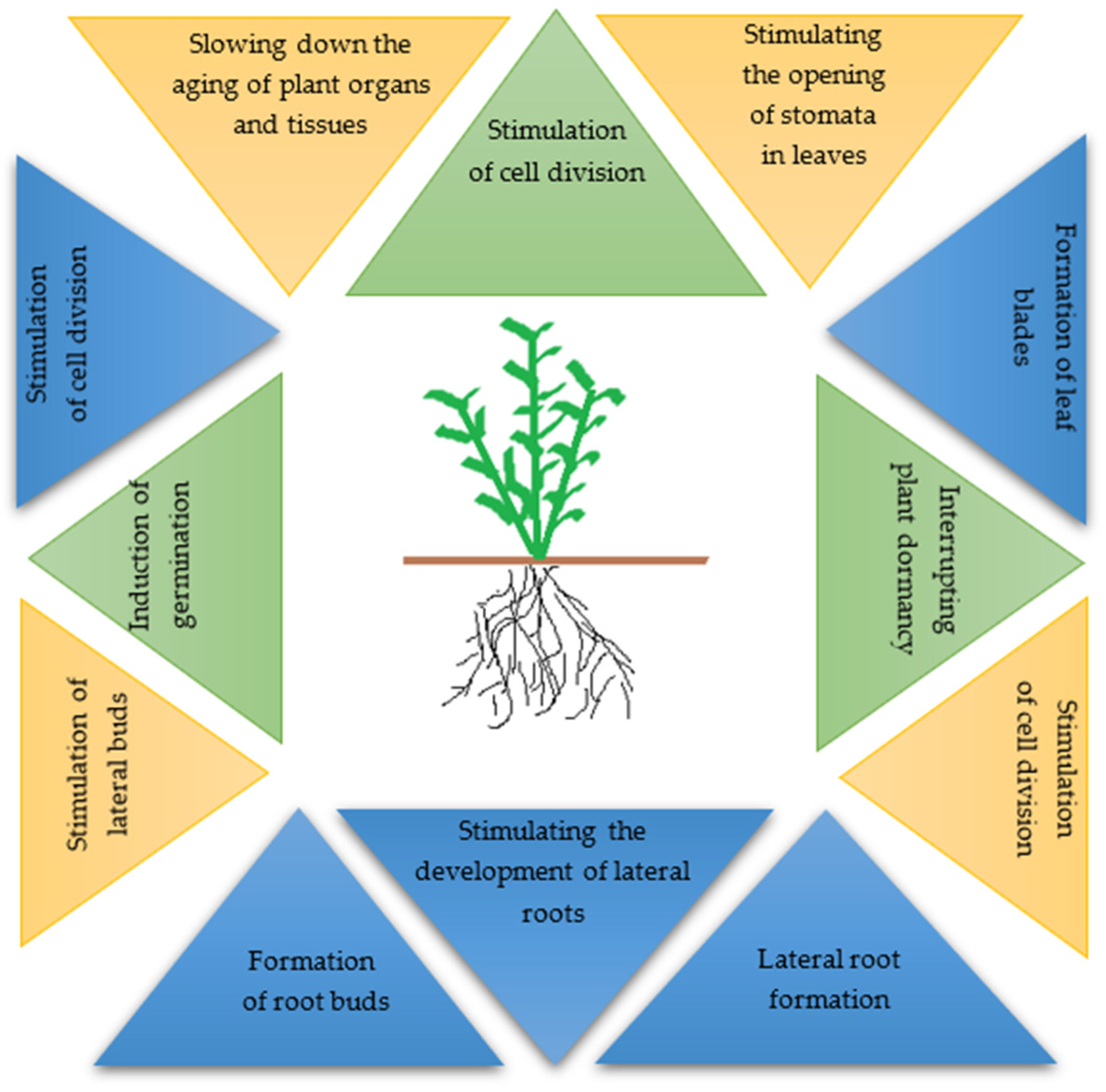
2. The Function and Role of Hormones in Plant Growth and Development Regulation
2.1. The Importance of Cytokinins in Plant Growth and Development Processes
2.2. The Importance of Auxins in Plant Growth and Development Processes
2.3. Effect of Auxins and Cytokinins on Selected Physiological Parameters of Plants

{kind=link}
{kind=link}
{kind=link}
| Plant Physiological Precesses | Crops | |||||
|---|---|---|---|---|---|---|
| Barley Leaves and Seedlings | Soybean Leaves | Cotton Leaves | Wheat Leaves | Alfalfa Leaves | Cabbage Leaves | |
| Increased assimilation CO2 | x [88,89] | x [81] o [81] | x [91,93] o [91,93] | |||
| Increase in total chlorophyll content | x [95] | x [89] | o [17] | |||
| Increase in the content of chlorophyll “a” | o [94,96] | |||||
3. The Use of Phytohormones as Growth Regulators in Plant Production
3.1. Effect of Auxins and Cytokinins on Crop Yield and Morphometry
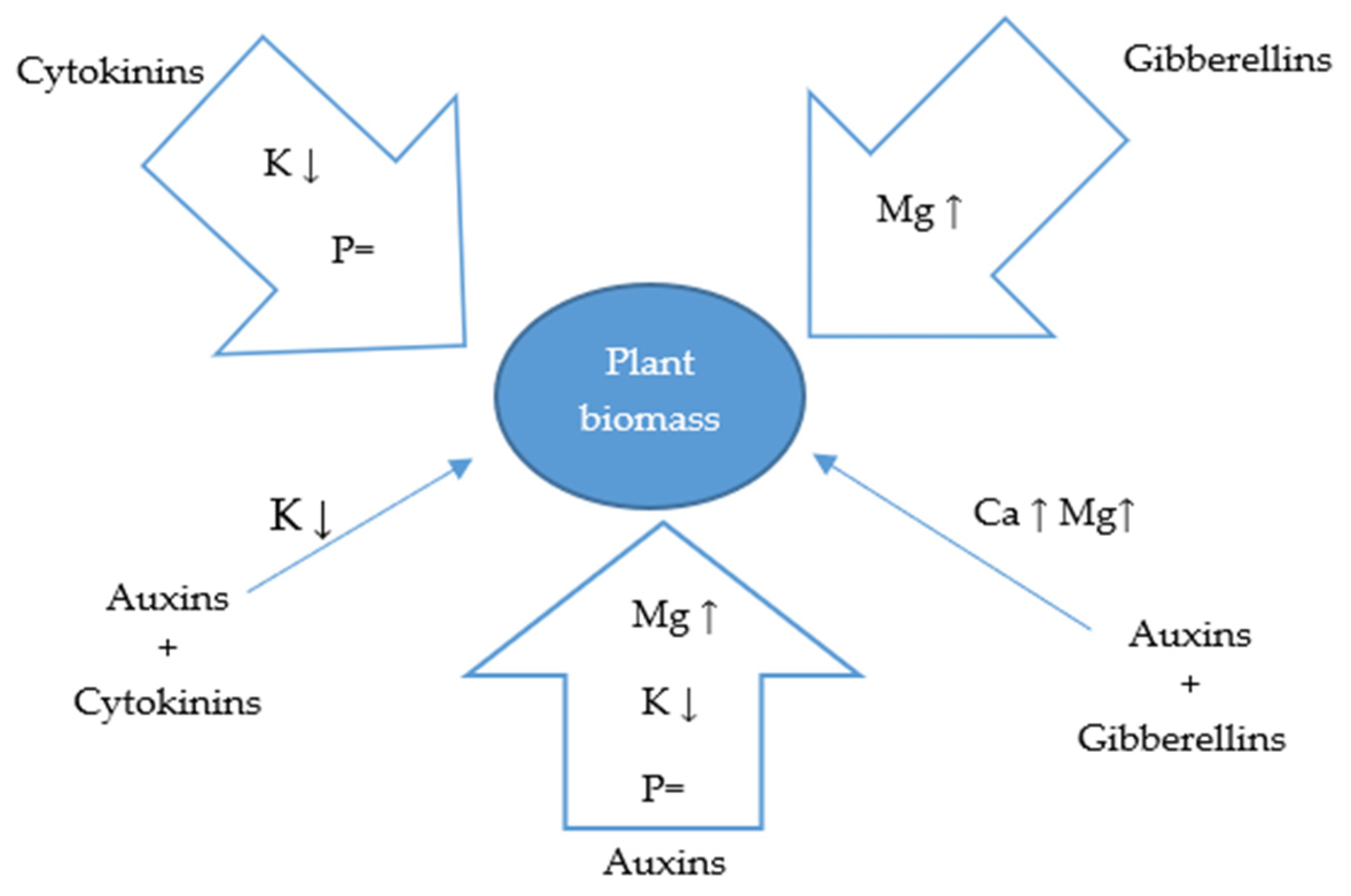
3.2. Effect of Auxins and Cytokinins on Chemical Composition of Plant Biomass
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pospišilová, J.; Synkova, H.; Rulcová, J. Cytokinins and water stress. Biol. Plant 2000, 43, 321–328. [Google Scholar] [CrossRef]
- Jakubowska, A.; Zielińska, E.; Kowalczyk, S. Metabolism and transport of auxin in plants. Post. Bioch. 2001, 47, 169–182. (In Polish) [Google Scholar]
- Baluška, F.; Šamaj, J.; Menzel, D. Polar transport of auxin: Carrier -mediated flux across the plasma membrane or neurotransmitter–like secretion? Trends Cell Biol. 2003, 13, 282–285. [Google Scholar] [CrossRef] [PubMed]
- Jakubowska, A. Synthesis and hydrolysis of plant hormone conjugates in the regulation of active hormone levels. Postępy Biol. Komórki 2003, 30, 565–587. (In Polish) [Google Scholar]
- Leyser, O. Dynamic integration of auxin transport and signaling. Curr. Biol. 2006, 16, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Czapla, J.; Nogalska, A.; Stasiulewicz, L. Synthetic auxin effect on the yield and the mineral soybeans. Acta Sci. Pol. Agric. 2003, 2, 123–131. (In Polish) [Google Scholar]
- Flasiński, M.; Hąc-Wydro, K. Natural vs synthetic auxin: Studies on the interactions between plant hormones and biological membrane lipids. Environ. Res. 2014, 133, 123–134. [Google Scholar] [CrossRef]
- Act of 10 July 2007 on Fertilizers and Fertilization. 2007. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU20071471033 (accessed on 17 March 2023).
- Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market; EU: Maastricht, The Netherlands, 2009.
- Nogalska, A.; Czapla, J. Yielding of spring barley depending on the application of growth regulators, and mixtures thereof with magnesium sulfate. Pol. J. Nat. Sci. 2003, 12, 37–51. (In Polish) [Google Scholar]
- Wybouw, B.; De Rybel, B. Cytokinin–a developing story. Trends Plant Sci. 2019, 24, 177–185. [Google Scholar] [CrossRef]
- Ortiz-Castro, R.; Campos-García, J.; López-Bucio, J. Pseudomonas putida and Pseudomonas fluorescens Influence Arabidopsis Root System Architecture Through an Auxin Response Mediated by Bioactive Cyclodipeptides. J. Plant Growth Regul. 2020, 39, 254–265. [Google Scholar] [CrossRef]
- Holalu, S.V.; Reddy, S.K.; Finlayson, S.A. Low Red Light: Far Red Light Inhibits Branching by Promoting Auxin Signaling. J. Plant Growth Regul. 2021, 40, 2028–2036. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, Y.; Zhang, X.; Du, H.; Xu, B.; Huang, B. Melatonin suppression of heat-induced leaf senescence involves changes in abscisic acid and cytokinin biosynthesis and signaling pathways in perennial ryegrass (Lolium perenne L.). Environ. Exp. Bot. 2017, 138, 36–45. [Google Scholar] [CrossRef]
- Downs, C.G.; Somerfield, S.D.; Davey, M.C. Cytokinin treatment senescence but not sucrose loss in harvested braccoli. Post. Biol. Technol. 1997, 11, 93–100. [Google Scholar] [CrossRef]
- Skutnik, E.; Rabiza-Świder, J.; Wachowicz, M.; Łukaszewska, A.J. Senescence of cut leaves of Zentedeschia aethiopica and Z. elliottiana. Part I. Chlorophyll degradation. Acta Sci. Pol. Hortorum Cultus 2004, 3, 57–65. [Google Scholar]
- Costa, M.L.; Civello, P.M.; Chaves, A.R.; Martinez, G.A. Effect of ethephon and 6-benzylaminopuryne on chlorophyll degrading enzymes and peroxidase-linked chlorophyll bleaching during post-harvest senescence of braccoli (Brassica oleracea L.) at 20 °C. Postharvest Biol. Technol. 2005, 35, 191–199. [Google Scholar] [CrossRef]
- Haiser, M.G.; Jönsson, H. Modeling auxin transport and plant development. J. Plant Growth Regul. 2006, 25, 302–312. [Google Scholar]
- Kramer, M.E.; Bennett, J. Auxin transport: A field in flux. Trends Plant Sci. 2006, 11, 383–386. [Google Scholar] [CrossRef]
- Zhao, Y. The role of local biosynthesis of auxin and cytokinin on plant development. Curr. Opi. Plant Biol. 2008, 11, 16–22. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Márquez, G.; Alarcón, M.V.; Salguero, J. Cytokinin Inhibits Lateral Root Development at the Earliest Stages of Lateral Root Primordium Initiation in Maize Primary Root. J. Plant Growth Regul. 2019, 38, 83–92. [Google Scholar] [CrossRef]
- Fagerness, M.J.; Penner, D. Spray application parameters that influence the growth inhibiting effects of trinexapac-ethyl. Crop Sci. 1998, 38, 1028–1035. [Google Scholar] [CrossRef]
- Qian, Y.L.; Engelke, M.C. Influence of Trinexapac-Ethyl on Diamond Zoysiagrass in Shade Environment. Crop Sci. 1999, 39, 202–208. [Google Scholar] [CrossRef]
- Sawicka, B. Effects of growth regulators Mival and Potejtin application in potato cultivation. Part I “Influence of growth regulators on incidense of common scab (Streptomyces sp.)”. Ann. Agric. Sci. Ser. E-Plant Prot. 1999, 28, 43–54. [Google Scholar]
- Sawicka, B. Effects of growth regulators Mival and Potejtin application in potato cultivation. Part II “The influence of growth regulators on incidence of Rhisoctonia solani sclerotia bearing tubers”. Ann. Agric. Sci. Ser. E-Plant Prot. 1999, 28, 55–66. [Google Scholar]
- Sawicka, B. Effects of growth regulators Mival and Potejtin application in potato cultivation. Part III. The influence of growth regulators on storage losses of tubers. Ann. Agric. Sci. Ser. E-Plant Prot. 1999, 28, 67–79. [Google Scholar]
- Mikos-Bielak, M. Exogenous growth regulators in potato cultivation. Ann. Univ. Mariae Curie-Sklodowska Sect. A 2005, 60, 281–292. [Google Scholar]
- Abas, L.; Benjamins, R.; Malenica, N.; Paciorek, T.; Wiśniewska, J.; Moulinier-Anzola, J.C.; Sieberer, T.; Frimal, J.; Luchning, C. Intrecellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved root gravitropism. Nat. Cell Biol. 2006, 8, 249–256. [Google Scholar] [CrossRef]
- Heisler, M.G.; Ohno, C.; Das, P.; Sieber, P.; Reddy, G.V.; Long, J.A.; Meyerowitz, E.M. Patterens of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr. Biol. 2005, 15, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Xiao-Ping, S.; Xi-Gui, S. Cytokinin and auxin-induced stomatal opening is related to the change of nitric oxide levels in guard cells in broad bean. Physiol. Plant 2006, 128, 569–5579. [Google Scholar] [CrossRef]
- Werner, T.; Schmülling, T. Cytokinin action in plant development. Curr. Opin. Plant Biol. 2009, 12, 527–538. [Google Scholar] [CrossRef]
- Gan, L.; Song, M.; Wang, X.; Yang, N.; Li, H.; Liu, X.; Li, Y. Cytokinins is involved in regulation of tomato pericarp thickness and fruit size. Hortic. Res. 2022, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- Raspor, M.; Motyka, V.; Ninković, S.; Malbeck, J.; Dobrev, P.I.; Zdravkovic-Korac, S.; Simonovic, A.; Cosic, R.; Cingel, A.; Savic, J.; et al. Overexpressing AtCKX1 in Potato Plants grown In Vitro: The Effects on Cytokinin Composition and Tuberization. J. Plant Growth Regul. 2021, 40, 37–47. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Yang, L.; Chan, Z. Melatonin Antagonizes Cytokinin Responses to Stimulate Root Growth in Arabidopsis. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Šmehilová, M.; Galuszka, P.; Bilyru, K.D.; Jaworek, P.; Kowalska, M.; Sebela, M.; Sedlářová, M.; English, J.T.; Fébort, I. Subcellular lokalization and biochemical comparison of cytosolic and secreted cytokinin dehydrogenese enzymes from maize. J. Exp. Bot. 2009, 60, 2701–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubo, Y.O.; Blakley, I.C.; Yamburenko, M.V.; Worthen, J.M.; Street, I.H.; Franco-Zorrilla, J.M.; Zhang, W.; Hill, K.; Raines, T.; Solano, R.; et al. Cytokinin induces genome-wide binding of the type-B response regulator ARR10 to regulate growth and development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, E5995–E6004. [Google Scholar] [CrossRef] [Green Version]
- Galuszka, P.; Fébort, I.; Sebela, M.; Sauer, P.; Jacobsen, S.; Pec, P. Cytokinin oxidase or dehydrogenase? Mechanism of cytokinin degradation in cereals. Eur. J. Biochem. 2001, 268, 450–461. [Google Scholar] [CrossRef]
- Nordström, A.; Tarkowski, P.; Tarkowsks, D.; Norbeak, R.; Astot, C.; Dolezal, K.; Sandberg, G. Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: A factor of potential importance for auxin-cytokinin-regulated development. Proc. Natl. Acad. Sci. USA 2004, 101, 8039–8044. [Google Scholar] [CrossRef] [Green Version]
- Dierck, R.; De Keyser, E.; De Riek, J.; Dhooghe, E.; Van Huylenbroeck, J.; Prinsen, E. Change in auxin and cytokinin levels coincides with altered expression of branching genes during axillary bud outgrowth in chrysanthemum. PLoS ONE 2016, 11, e0161732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudo, T.; Kiba, T.; Sakakibara, H. Metabolism and long-distance translocation of cytokinins. J. Integr. Plant Biol. 2010, 52, 53–60. [Google Scholar] [CrossRef]
- Camas-Reyes, A.; Vuelvas-Nolasco, R.; Cabrera-Ponce, J.L.; Pereyra-Alférez, B.; Molina-Torres, J.; Martínez-Antonio, A. Effect of different cytokinins on shoot outgrowth and bioactive compounds profile of lemograss essential oil. Int. J. Plant Biol. 2022, 13, 25. [Google Scholar] [CrossRef]
- Janečková, H.; Husičková, A.; Lazár, D.; Ferretti, U.; Pospíšil, P.; Špundová, M. Exogenous application of cytokinin during dark senescence eliminates the acceleration of photosystem II impairment caused by chlorophyll b deficiency in barley. Plant Physiol. Biochem. 2019, 136, 43–51. [Google Scholar] [CrossRef]
- Pons, T.L.; Jordi, W.; Kuiper, D. Acclimation of plants to light gradients in leaf canopies: Evidence for a possible role for cytokinins transported in the transpiration stream. J. Exp. Bot. 2001, 52, 1563–1574. [Google Scholar] [CrossRef]
- Roman, H.; Girault, T.; Le Gourrierec, J.; Leduc, N. In silico analysis of 3 expansin gene promoters reveals 2 hubs controlling light and cytokinins response during bud outgrowth. Plant Signal Behav. 2017, 12, e-1284725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaller, G.E.; Street, I.H.; Kieber, J.J. Cytokinin and the cell cycle. Curr. Opin. Plant Biol. 2014, 21, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Talla, S.K.; Panigrahy, M.; Kappara, S.; Nirosha, P.; Neelamraju, S.; Ramanan, R. Cytokinin delays dark-induced senescence in rice by maintaining the chlorophyll cycle and photosynthetic complexes. J. Exp. Bot. 2016, 67, 1839–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Giordano, W.; Hirsch, A.M. Cytokinin induces expansin gene expression in Melilotus alba Desr. wild-type and the non-nodulating, non-mycorrhizal (Nod-Myc-) mutant Masym3. Plant Signal Behav. 2008, 3, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusaba, M.; Tanaka, A.; Tanaka, R. Stay-green plants: What do they tell us about the molecular mechanism of leaf senescence. Photosynth. Res. 2013, 117, 221–234. [Google Scholar] [CrossRef]
- Wu, W.; Du, K.; Kang, X.; Wei, H. The diverse roles of cytokinins in regulating leaf development. Hortic. Res. 2021, 8, 118. [Google Scholar] [CrossRef]
- Buchman-Vollaston, V.; Pagr, T.; Harrison, E.; Breeze, E.; Lim, P.O.; Nam, H.G.; Lin, J.F.; Swidzinski, J.; Ishizaki, K.; Leaver, C.J. Comparative transcriptome analysis reveals significant differences in gene exspresion and signaling pathways between development and dark/starvation-induced senescence in Arabidopsis. Plant J. 2005, 42, 567–585. [Google Scholar] [CrossRef]
- Burr, C.A.; Sun, J.; Yamburenko, M.V.; Willoughby, A.; Hodgens, C.; Boeshore, S.L.; Elmore, A.; Atkinson, J.; Nimchuk, Z.L.; Bishopp, A.; et al. The HK5 and HK6 cytokinin receptors mediate diverse developmental pathways in rice. Development 2020, 147, 191734. [Google Scholar] [CrossRef]
- Frimal, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jürgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Frimal, J.; Wiśniewska, J.; Benková, E.; Mendagen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 41, 806–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanneste, S.; Friml, J. Auxin: A Trigger for Change in Plant Development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Kawamura, A.; Suzuki, T.; Segami, S.; Maeshima, M.; Polyn, S.; De Veylder, L.; Sugimoto, K. Transcriptional activation of auxin biosynthesis drives developmental reprogramming of differentiated cells. Plant Cell 2022, 34, 4348–4365. [Google Scholar] [CrossRef]
- Herrera-Ubaldo, H. A case of identity: Activation of auxin biosynthesis drives cell reprogramming. Plant Cell 2022, 34, 4124–4125. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.A.; Desind, S.; Hallak, A.; Alhaddad, A.; Smalley, J.V.; Campanella, J.J. Correction to: Examination of the M20D Auxin Conjugate Peptidase Family from Hornwort and Implications on the Evolution of the Tracheophytes. J. Plant Growth Regul. 2022, 41, 2707. [Google Scholar] [CrossRef]
- Morris, D.A.; Frimal, J.; Zažímalová, E. The transport of auxins. In Plant Hormones: Biosynthesis, Signal Transduction, Action; Davies, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 437–470. [Google Scholar]
- Márquez, G.; Alarcón, M.V.; Salguero, J. Differential responses of primary and lateral roots to indole-3-acetic acid, indole-3-butyric acid, and 1-naphthaleneacetic acid in maize seedlings. Biol. Plant 2016, 60, 367–375. [Google Scholar] [CrossRef]
- Bartel, B.; LeClere, S.; Magidion, M.; Zolman, B.K. Inputs to the active indole-acetic acid pool: De novo synthesis, conjugate hydrolysis and indole-3-butyric acid β-oxidation. J. Plant Growth Regul. 2001, 20, 198–216. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [Green Version]
- Novitskaya, L.L.; Tarelkina, T.V.; Galibina, N.A.; Moshchenskaya, Y.L.; Nikolaeva, N.N.; Nikerova, K.M.; Podgornaya, M.N.; Sofronova, I.N.; Semenova, L.I. The Formation of Structural Abnormalities in Karelian Birch Wood is Associated with Auxin Inactivation and Disrupted Basipetal Auxin Transport. J. Plant Growth Regul. 2020, 39, 378–394. [Google Scholar] [CrossRef]
- Ljung, K.; Hull, A.K.; Celenza, J.; Yamada, M.; Estelle, M.; Normalny, J.; Sandberg, G. Sites and regulation of auxin biosynthesis in Arabidopsis roots. Plant Cell 2005, 17, 1090–1104. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Pan, J.; Wang, H.; Reiter, R.J.; Li, X.; Mou, Z.; Zhang, J.; Yao, Z.; Zhao, D.; Yu, D. Melatonin inhibits seed germination by crosstalk with abscisic acid, gibberellin, and auxin in Arabidopsis. J. Pineal Res. 2021, 70, e12736. [Google Scholar] [CrossRef] [PubMed]
- Lobo, L.L.B.; da Silva, M.S.R.d.; Carvalho, R.F.; Rigobelo, E.C. The Negative Effect of Coinoculation of Plant Growth-Promoting Bacteria Is Not Related to Indole-3-Acetic Acid Synthesis. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Shin, S.Y.; Choi, Y.; Kim, S.-G.; Park, S.-J.; Park, J.-S.; Moon, K.-B.; Kim, H.-S.; Jeon, J.H.; Cho, H.S.; Lee, H.-J. Submergence promotes auxin-induced callus formation through ethylene-mediated post-transcriptional control of auxin receptors. Mol. Plant 2022, 15, 1947–1961. [Google Scholar] [CrossRef]
- Rahman, A. Auxin: A regulator of cold stress response. Physiol. Plant 2013, 147, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kreiser, M.; Giblin, C.; Murphy, R.; Fiesel, P.; Braun, L.; Johnson, G.; Wyse, D.; Cohen, J.D. Conversion of Indole-3-Butyric Acid to Indole-3-Acetic Acid in Shoot Tissue of Hazelnut (Corylus) and Elm (Ulmus). J. Plant Growth Regul. 2016, 35, 710–721. [Google Scholar] [CrossRef]
- Yang, L.; You, J.; Li, J.; Wang, Y.; Chan, Z. Melatonin promotes Arabidopsis primary root growth in an IAA dependent manner. J. Exp. Bot. 2021, 72, 5599–5611. [Google Scholar] [CrossRef]
- Hagen, G.; Guilfoyle, T. Auxin responsive gene expression: Genes promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef]
- Xu, C.; Cao, H.; Zhang, Q.; Wang, H.; Xin, W.; Xu, E.; Zhang, S.; Yu, R.; Yu, D.; Hu, Y. Control of auxin-induced callus formation by bZIP59–LBD complex in Arabidopsis regeneration. Nat. Plants 2018, 4, 108–115. [Google Scholar]
- Zhou, J.; Sittmann, J.; Guo, L.; Xiao, Y.; Huang, X.; Pulapaka, A.; Liu, Z. Gibberellin and auxin signaling genes RGA1 and ARF8 repress accessory fruit initiation in diploid strawberry. Plant Physiol. 2021, 185, 1059–1075. [Google Scholar] [CrossRef]
- Sharma, A.; Wang, J.; Xu, D.; Tao, S.; Chong, S.; Yan, D.; Li, Z.; Yuan, H.; Zheng, B. Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Carya cathayensis plants. Sci. Total Environ. 2020, 713, 136675. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Gong, M.; Xu, X.; Li, H.; Deng, W. Roles of auxin in the growth, development, and stress tolerance of horticultural plants. Cells 2022, 11, 2761. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.; Kuhn, N.; Restovic, F.; Meyer-Regueiro, C.; Madariaga, M.; Arce-Johnson, P. The Glucose-Related Decrease in Polar Auxin Transport During Ripening and its Possible Role in Grapevine Berry Coloring. J. Plant Growth Regul. 2023, 42, 365–375. [Google Scholar] [CrossRef]
- Quittenden, L.J.; McAdam, E.L.; Davies, N.W.; Ross, J.J. Evidence that Indole-3-Acetic Acid is Not Synthesized Via the Indole-3-Acetamide Pathway in Pea Roots. J. Plant Growth Regul. 2014, 33, 831–836. [Google Scholar] [CrossRef]
- Simon, M.R. Gene action and heritability for photosynthetic activity in two wheat crosses. Euphytica 1994, 76, 235–238. [Google Scholar] [CrossRef]
- Michałek, S.; Borowski, E. Response of selected soybean (Glycine max L.) cultivars to drought. Zesz. Nauk. AR Kraków 1998, 333, 905–907. (In Polish) [Google Scholar]
- Fu, J.; Huang, B.; Zhang, G. Physiological and biochemical changes during seed filling in relation to leaf senescence in soybean. Biol. Plant. 2000, 43, 545–548. [Google Scholar] [CrossRef]
- Subrahmanyam, D. Interrelationship between leaf gas-exchange characteristics, area leaf mass, and yield in soybean (Glycine max L. Merr) genotypes. Photosyntetica 2002, 40, 441–444. [Google Scholar] [CrossRef]
- Luquez, V.M.; Giuamet, J.J.; Montaldi, E.R. Net photosynthetic and transpiration rates in chlorophyll-deficient isoline of soybean under well-watered and drought conditions. Photosynthetica 1997, 34, 125–131. [Google Scholar] [CrossRef]
- Starck, Z. Niektóre aspekty różnicowania reakcji roślin na niekorzystne warunki środowiska-stare problemy, nowa interpretacja. Zesz. Prob. Post. Nauk Rol. 1999, 469, 145–159. (In Polish) [Google Scholar]
- Wróbel, J. The effect of applying the nitrophenolates and nitroguaiacolate solution in cultivation of Salix viminalis L. on anthropogenic substratum. Zesz. Probl. Post. Nauk Rol. 2002, 481, 615–620. [Google Scholar]
- Muller, J.E.; Bergman, H. Plant cellular response to water deficit. Plant Growth Regul. 1996, 2, 41–46. [Google Scholar]
- Michałek, S. Growth, gas exchange and yielding of several Polish soybean cultivars under drought conditions. Zesz. Problem. Post. Nauk Rol. 1999, 469, 217–223. (In Polish) [Google Scholar]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigeus, M.I.; Ricardo, C.P.P.; Osǒrio, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How plant cope with water stress in the field. Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.Y.; Azhar, N.; Hussain, M. Indole acetic acid (IAA) induced changes in growth, relative water contents and gas exchange attributes of barley (Hordeum vulgare L.) grown under water stress canditions. Plant Growth Regul. 2006, 50, 85–90. [Google Scholar] [CrossRef]
- Aldesuquy, H.S. Effect of indol-3-yl acetic acid on photosynthetic characteristics of wheat flag leaf during grain filling. Photosynthetica 2000, 38, 135–141. [Google Scholar] [CrossRef]
- Pospišilová, J. Interaction of cytokinins and abscisic acid during regulation of stomatal opening in bean leaves. Photosynthetica 2003, 41, 49–56. [Google Scholar] [CrossRef]
- Muthuchelian, K.; Murugan, C.; Harigovindan, R.; Nedunchezhian, N.; Kulandaivelu, G. Effect of triacontanol in flooded Erythrina variegata deedlings. Changes in CO2 fixation, and ribulose-1,5-bisphosphate carboxylase, photosystem and nitrate reductase activites. Photosynthetica 1994, 30, 407–413. [Google Scholar]
- Pandey, D.M.; Goswami, C.L.; Kumar, B.; Jain, S. Hormonal regulation of photosynthetic enzymes in cotton under water stress. Photosynthetica 2000, 38, 403–407. [Google Scholar] [CrossRef]
- Jiang, H.; Xu, D. The cause of the difference in leaf net photosynthesis rate between two soybean cultivars. Photosynthetica 2001, 39, 453–459. [Google Scholar] [CrossRef]
- Pandey, D.M.; Goswami, C.L.; Kumar, B. Physological effects of plant hormones in cotton under drought. Biol. Plant. 2003, 47, 535–540. [Google Scholar] [CrossRef]
- Galdallah, M.A.A. Effect of indole-3-acetic acid and zinc on the growth, osmotic potential and soluble carbon and nitrogen components of soybean plants growing under water deficit. J. Arid Environ. 2000, 44, 451–467. [Google Scholar] [CrossRef]
- Skalska, M. Effect of growth regulators on morphological features, chlorophyll content and yield of alfalfa (Medicago sativa L.) seeds in pot experiments. Biul. Inst. Hod. Aklim. Rosl. 1992, 184, 59–65. (In Polish) [Google Scholar]
- Nowak, A.; Wróbel, J. Impact of selected growth regulators on yielding of soybean (Glycine max L. Merr) in control requirements of substrate moisture. Rosl. Oleiste-Oilseed Crops 2010, 31, 124–132. (In Polish) [Google Scholar]
- Von Richthofen, J.S. What do European farmers think about grain legumes. Grain Legumes 2006, 45, 14–15. [Google Scholar]
- Ulmasov, T.; Murfett, J.; Hagen, G.; Guilfoyle, T.J. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 1997, 9, 1963–1971. [Google Scholar]
- Harms, H.; Nowak, G. Effect of foliar applied nitrogen and kinetin on nitrogen redistribution during growth in wheat. Grain growth, accumulation and redistribution of nitrogen. Angew. Bot. 1990, 64, 253–260. [Google Scholar]
- Wójcik, S.; Majewski, K. Effect of selected growth simulators on biometric features and on crop yield of buckwheat (Fagopyrum esculentum Moench). Fagopyrum 1991, 11, 47–50. [Google Scholar]
- Aufhammer, W.; Federolf, K.G. Effects of seed treatments with plant growth regulators on development and yield of winter hard wheat (Triticum durum). Bodenkulture 1992, 43, 99–108. [Google Scholar]
- Nowak, A.; Wróbel, J. The effect of growth regulators on concent of assimilation pigment in leaves of three soybean (Glycine max L. Merr). Rosl. Oleiste-Oilseed Crops 2010, 31, 351–359. (In Polish) [Google Scholar]
- Sosnowski, J.; Malinowska, E.; Jankowski, K.; Król, J.; Redzik, P. An astimation of the effects of synthetic auxin and cytokinin and the time of their application on some morphological and physiological characteristics of Medicago x varia T. Martyn. Saudi J. Biol. Sci. 2017, 26, 66–73. [Google Scholar] [CrossRef]
- Reinecke, D.M. 4-Chloroindole-3-acetic and plant growth. Plant Growth Regul. 1999, 27, 3–13. [Google Scholar] [CrossRef]
- Barclay, G.F.; McDavid, C.R. Effect of benzyloaminopuryne on fruit set and seed development in pigeonpea (Cajanus cajan). Sci. Hortic. 1998, 72, 81–86. [Google Scholar] [CrossRef]
- Nowak, G.; Ciecko, Z. Effect of GA3 on the yield and value of forage turnip. Zesz. Nauk. Akad. Rol. Krakowie 1991, 262, 325–331. (In Polish) [Google Scholar]
- Kertikov, T.; Vasileva, V. Influence of some biologically active substances on grain yield and chemical composition in spring vetch. J. Mt. Agric. Balk. 2000, 3, 190–198. [Google Scholar]
- Kuang, A.; Peterson, C.M.; Dute, R.R. Pedicel abcission and rachis morphology of soybean as influenced by benzylaminopurine and the presence of poods. J. Plant Growth Regul. 1991, 10, 291–303. [Google Scholar] [CrossRef]
- Peterson, C.M.; Williams, J.C.; Kuang, A. Increased podset of determinae cultivars of soyabean, Glycine max, with 6-benzyloaminopurine. Bot. Gaz. 1990, 151, 322–330. [Google Scholar] [CrossRef]
- Rylott, P.D.; Smith, M.L. Effects of applied growth substances on pod set in broad beans (Vicia faba var. major). J. Agric. Sci. 1990, 114, 41–47. [Google Scholar] [CrossRef]
- Qifu, M.; Longnecker, N.; Atkins, C. Exogenous cytokinin and nitrogen do not increase grain yield in narrow-leafed lupins. Crop Sci. 1998, 38, 717–721. [Google Scholar]
- Kuang, A.; Peterson, C.M.; Dutr, R.R. Changes in soybean raceme and petiole anatomy induced by 6-benzyloaminopurine. Ann. Bot. 1991, 67, 23–27. [Google Scholar] [CrossRef]
- Resse, R.N.; Dybing, C.D.; White, C.A.; Page, S.M.; Larson, J.E. Expression of vegetative storage protein (VSP-β) in soybean raceme tissues in response to flower set. J. Exp. Bot. 1995, 46, 957–964. [Google Scholar] [CrossRef]
- Khan, N.A.; Khan, M.; Ansari, H.R. Auxin and defoliaton effects on photosynthesis and ethylene evolution in mustard. Sci. Hortic. 2002, 96, 43–51. [Google Scholar] [CrossRef]
- Pal, A.K.; Das, S.N. Effect of IAA on growth and flowering of Lilium longiflorum. Orissa J. Hortic. 1990, 18, 18–21. [Google Scholar]
- Jamil, M.; Rahman, M.M.; Hossain, M.M.; Hossain, M.T.; Karim, A.S. Effect of plant growth regulators on flower and bulb production of hippeastrum (Hippeastrum hybridum Hort.). Bangladesh J. Agric. Res. 2016, 40, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Karaguzel, O.; Alian, S.; Doran, I.; Sogut, Z. Improvement of gladiolus by growth regulator and nutrient management. J. Jpn. Soc. Hortic. Sci. 1999, 68, 168–175. [Google Scholar]
- Prakash, V.; Jha, K.K. Physiology of gladiolus. J. Appl. Biol. 1998, 8, 24–28. [Google Scholar]
- Clifford, P.E.; Pentland, B.S.; Baylis, A.D. Effect of growth regulators on reproductive abscission in faba bean (Vicia faba cv. Troy). J. Agric. Sci. 1992, 119, 71–78. [Google Scholar] [CrossRef]
- Baylis, A.D.; Clifford, P.E. Control of reproductive abscission in grain legumes. Ann. Bull. Br. Soc. Pl. Growth Regul. 1991, 1, 1–12. [Google Scholar]
- Nagel, L.; Brewster, R.; Riedell, W.E.; Reese, R.N. Cytokinin regulation of flower and pod set soybeans (Glycine max L. Merr.). Ann. Bot. 2001, 88, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Wierzbowska, J.; Nowak, G.A. Effects of cytokinins and auxins application on potassium managment of spring wheat in relation to level on mineral fertilization. Nat. Sci. 2000, 7, 81–92. [Google Scholar]
- Wierzbowska, J.; Nowak, G.A. The influence of growth regulators and increasing doses of nitrogen on the phosphorus and potassium management of spring wheat. Pol. J. Nat. Sci. 2002, 12, 7–19. [Google Scholar]
- Wierzbowska, J.; Zuk-Golaszewska, K.; Bochenek, A. Effect of mineral fertilization and growth regulators on the content of mineral components in pea plants. J. Elementol. 2007, 12, 207–215. [Google Scholar]
- Wierzbowska, J.; Bowszys, T. Effect of growth regulators applied together with different phosphorus fertilization levels on the content and accumulation of potassium, magnesium and calcium in spring wheat. J. Elementol. 2008, 13, 411–422. [Google Scholar]
- Meuwly, P.; Pilet, P. Local treatment with indole-3-acetic acid induces differential growth responses in Zea mays L. root. Planta 1991, 185, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Hayat, S.; Hasan, S.; Ahmad, A. A comparative effect of IAA and 4-Cl-IAA on growth, nodulation and nitrogen fixation in Vigna radiata (L.). Acta Physiol. Plant. 2008, 30, 35–41. [Google Scholar] [CrossRef]
- Weijers, D.; Schlereth, A.; Ehrismann, J.S.; Schwank, G.; Kientz, M.; Jurgens, G. Auxin triggers transient local signalling for cell specification in Arabidopsis embryogenesis. Dev. Cell 2006, 10, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Jankiewicz, L.S. (Ed.) Plant growth and development regulators. In Properties and Operation; Wyd. Naukowe PWN: Warszawa, Polska, 1997. (In Polish) [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sosnowski, J.; Truba, M.; Vasileva, V. The Impact of Auxin and Cytokinin on the Growth and Development of Selected Crops. Agriculture 2023, 13, 724. https://doi.org/10.3390/agriculture13030724
Sosnowski J, Truba M, Vasileva V. The Impact of Auxin and Cytokinin on the Growth and Development of Selected Crops. Agriculture. 2023; 13(3):724. https://doi.org/10.3390/agriculture13030724
Chicago/Turabian StyleSosnowski, Jacek, Milena Truba, and Viliana Vasileva. 2023. "The Impact of Auxin and Cytokinin on the Growth and Development of Selected Crops" Agriculture 13, no. 3: 724. https://doi.org/10.3390/agriculture13030724