
Native Trichoderma Isolates from Soil and Rootstock to Fusarium spp. Control and Growth Promotion of Humulus lupulus L. Plantlets
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trichoderma and Fusarium Isolates and Culture Collection
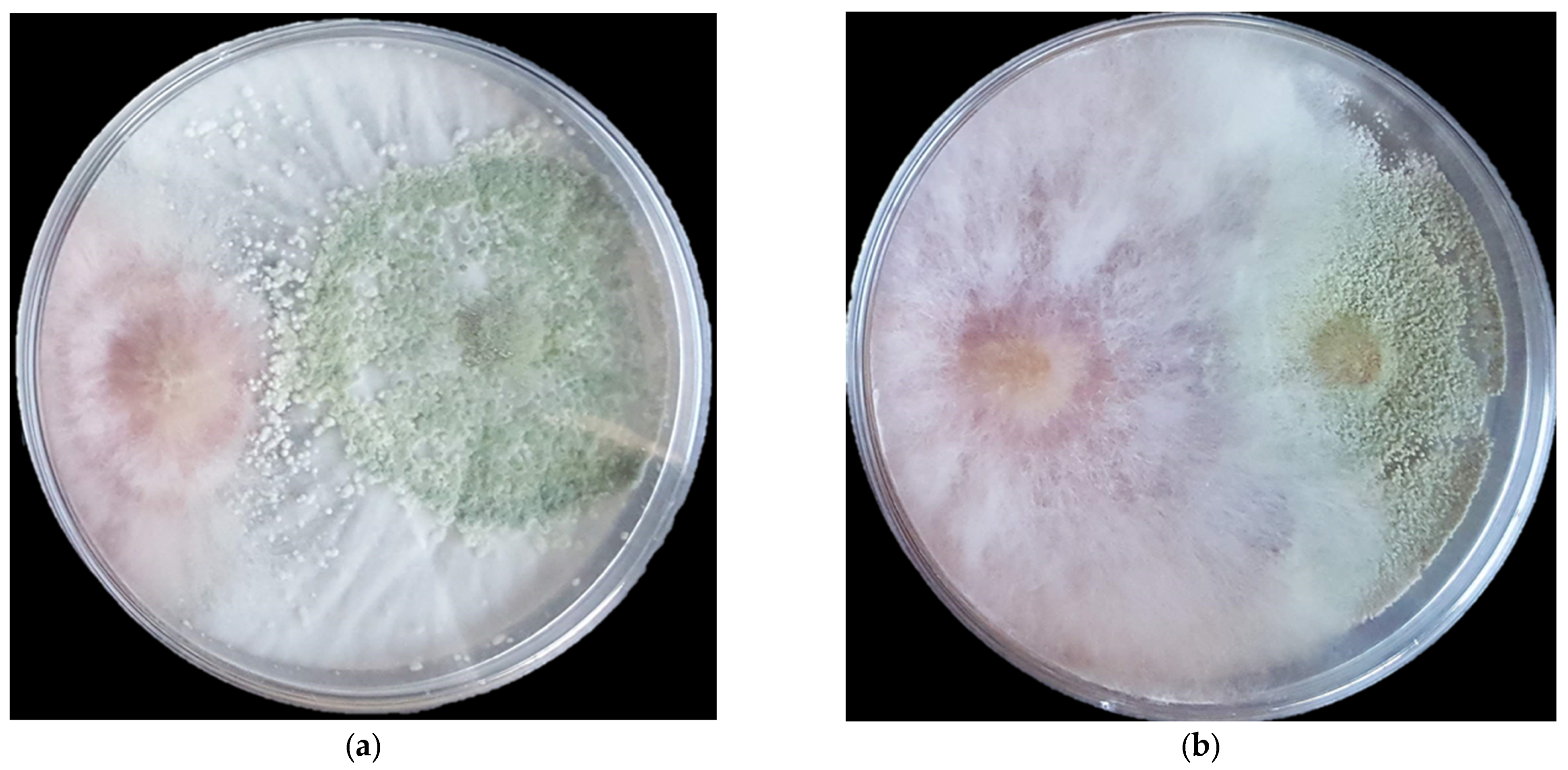
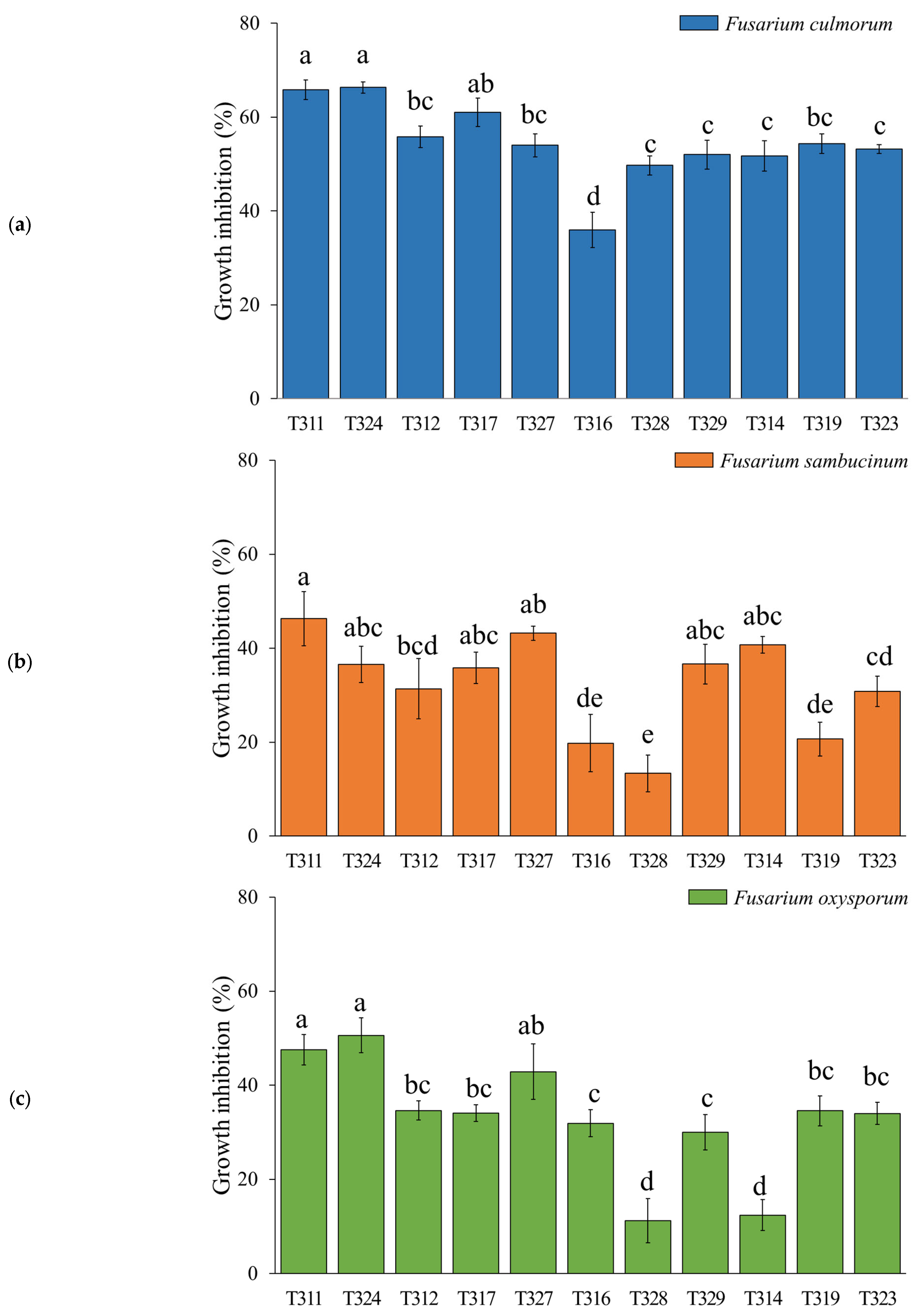
2.2. In Vitro Antifungal Assays-Direct Confrontation
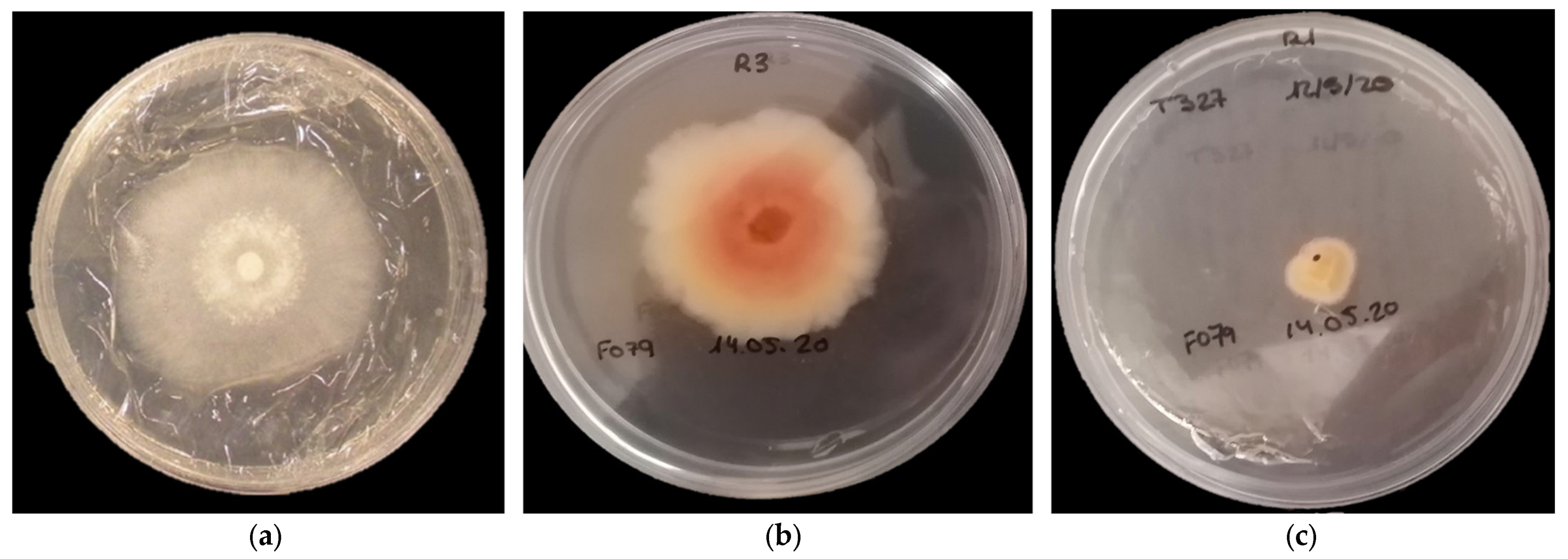
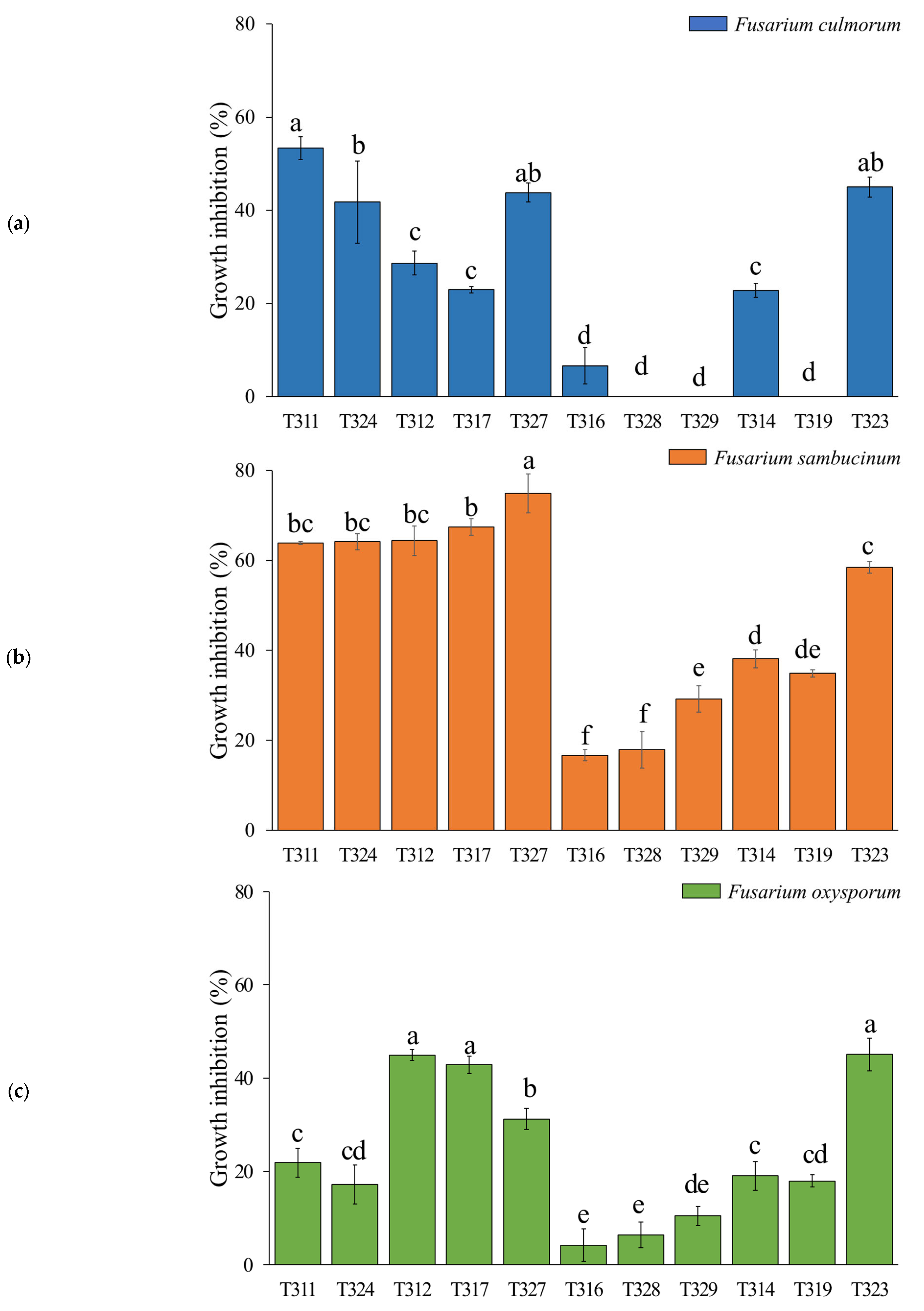
2.3. In Vitro Antifungal Assays-Membrane Assay
2.4. In Vivo Assay-Trichoderma Growth Promotion Activity
2.5. Statistical Analysis
3. Results
3.1. In Vitro Antifungal Assay-Direct Confrontation
3.2. In Vitro Antifungal Assay-Membrane Assay
3.3. In Vivo Assays-Trichoderma Growth Promotion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Lorenzana, A.; de Mendoza, A.H.; Magadán, J.A.; Seco, M.V. Estimating hop parameters prior to calculating economic thresholds of intervention against Phorodon humuli (Hemiptera: Aphididae). J. Hortic. Sci. Biotechnol. 2005, 80, 689–692. [Google Scholar] [CrossRef]
- Lorenzana, A.; de Mendoza, A.H.; Seco, M.V.; Casquero, P.A. Population development of Phorodon humuli and predators (Orius spp.) within hop cones: Influence of aphid density on hop quality. Crop. Prot. 2010, 29, 832–837. [Google Scholar] [CrossRef]
- Lorenzana, A.; de Mendoza, A.H.; Seco, M.V. Thresholds for management of Phorodon humuli (Hemiptera: Aphididae) on hops. J. Entomol. Sci. 2009, 44, 198–208. [Google Scholar] [CrossRef]
- Lorenzana, A.; de Mendoza, A.H.; Seco, V.; Campelo, P.; Casquero, P.A. Within-field distribution of the damson-hop aphid Phorodon humuli (Schrank) (hemiptera: Aphididae) and natural enemies on hops in Spain. Span. J. Agric. Res. 2017, 15, e1006. [Google Scholar] [CrossRef] [Green Version]
- Lorenzana, A.; de Mendoza, A.H.; Seco, M.V.; Campelo, M.P.; Casquero, P.A. Within-plant distribution of Phorodon humuli (Hemiptera: Aphididae) and natural enemies on hops with implications for sampling and management. Entomol. Sci. 2017, 20, 443–450. [Google Scholar] [CrossRef]
- Turechek, W.W.; Mahaffee, W.F.; Ocamb, C.M. Development of management strategies for hop powdery mildew in the Pacific Northwest. Plant Health Prog. 2001, 2, 1. [Google Scholar] [CrossRef]
- Mahaffee, W.F.; Turechek, W.W.; Ocamb, C.M. Effect of variable temperature on infection severity of Podosphaera macularis on hops. Phytopathology 2003, 93, 1587–1592. [Google Scholar] [CrossRef] [Green Version]
- Gent, D.H.; Grove, G.G.; Nelson, M.E.; Wolfenbarger, S.N.; Woods, J.L. Crop damage caused by powdery mildew on hop and its relationship to late season management. Plant Pathol. 2014, 63, 625–639. [Google Scholar] [CrossRef]
- Kappagantu, M.; Bullock, J.M.; Nelson, M.E.; Eastwell, K.C. Hop stunt viroid: Effect on host (Humulus lupulus) transcriptome and its interactions with hop powdery mildew (Podospheara macularis). Mol. Plant-Microbe Interact. 2017, 30, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Porteous-Álvarez, A.J.; Maldonado-González, M.M.; Mayo-Prieto, S.; Lorenzana, A.; Paniagua-García, A.I.; Casquero, P.A. Green strategies of powdery mildew control in hop: From organic products to nanoscale carriers. J. Fungi 2021, 7, 490. [Google Scholar] [CrossRef]
- Gryndler, M.; Krofta, K.; Gryndlerová, H.; Soukupová, L.; Hršelová, H.; Gabriel, J. Potentially dangerous fusarioid microorganisms associated with rot of hops (Humulus lupulus L.) plants in field culture. Plant Soil Environ. 2008, 54, 149–154. [Google Scholar] [CrossRef] [Green Version]
- de la Varga, A.L.; Cuadrado, C.R. (Eds.) Guía de Gestión Integrada de Plagas. Lúpulo; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2023; ISBN 978-84-491-1617-9.
- Pethybridge, S.J.; Hay, F.S.; Wilson, C.R.; Sherriff, L.J.; Leggett, G.W. First report of Fusarium crookwellense causing tip blight on cones of hop. Plant Dis. 2001, 85, 1208. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.A.M.F.; Araujo, L.; de Andrade, C.C.L.; Fagherazzi, M.M.; Brighenti, A.F.; de Martin, M.S.; Gomes, L.B.; Fernandes, J.; Duarte, V.; Arioli, C.J.; et al. First report of Fusarium meridionale causing canker in hop plants. Australas. Plant Dis. Notes 2022, 17, 13. [Google Scholar] [CrossRef]
- Bienapfl, J.C.; Ocamb, C.M.; Klein, R.; Nelson, M. Fusaium cone tip blight of Humulus lupulus. Acta Hortic. 2005, 668, 123–128. [Google Scholar] [CrossRef]
- Sabo, J.; Duric, T.; Jasnic, S. Fusarium fungi as a pathogen causing hop wilt. Plant Prot. Sci. 2002, 38, 308–310. [Google Scholar] [CrossRef] [Green Version]
- Mahaffee, W.F.; Pethybridge, S.J.; Gent, D.H. Compendium of Hop Diseases and Pests; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2009. [Google Scholar]
- O’Neal, S.D.; Walsh, D.B.; Gent, D.H.; Barbour, J.D.; Boydston, R.A.; George, A.E.; James, D.G.; Sirrine, J.R. (Eds.) Field Guide for Integrated Pest Management in Hops, 3rd ed.; Hop Industry Plant Protection Committee: Pullman, WA, USA, 2015. [Google Scholar]
- Alabouvette, C.; Olivain, C.; Steinberg, C. Biological control of plant diseases: The european situation. Eur. J. Plant. Pathol. 2006, 114, 329–341. [Google Scholar] [CrossRef]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Porteous-Álvarez, A.J.; Mayo-Prieto, S.; Álvarez-García, S.; Reinoso, B.; Casquero, P.A. Genetic response of common bean to the inoculation with indigenous Fusarium isolates. J. Fungi 2020, 6, 228. [Google Scholar] [CrossRef]
- Saberi-Riseh, R.; Moradi-Pour, M. A novel encapsulation of Streptomyces fulvissimus Uts22 by spray drying and its biocontrol efficiency against Gaeumannomyces graminis, the causal agent of take-all disease in wheat. Pest Manag. Sci. 2021, 77, 4357–4364. [Google Scholar] [CrossRef]
- Blaya, J.; López-Mondéjar, R.; Lloret, E.; Pascual, J.A.; Ros, M. Changes induced by Trichoderma harzianum in suppressive compost controlling Fusarium wilt. Pestic. Biochem. Physiol. 2013, 107, 112–119. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Del Mar Alguacil, M.; Pascual, J.A.; Van Wees, S.C.M. Phytohormone profiles induced by Trichoderma isolates correspond with their biocontrol and plant growth-promoting activity on melon plants. J. Chem. Ecol. 2014, 40, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, L.; Basińska-Barczak, A.; Ćwiek-Kupczyńska, H.; Gromadzka, K.; Popiel, D.; Stępień, Ł. Supressive effect of Trichoderma spp. on toxigenic Fusarium species. Pol. J. Microbiol. 2017, 66, 85–100. [Google Scholar] [CrossRef]
- Álvarez-García, S.; Mayo-Prieto, S.; Gutiérrez, S.; Casquero, P.A. Self-inhibitory activity of Trichoderma soluble metabolites and their antifungal effects on Fusarium oxysporum. J. Fungi 2020, 6, 176. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; An, Y.; Zhang, H.; Wang, Y.; Liu, Z. Trichoderma biofertilizer (mixTroTha) mediates Malus sieversii resistance to Alternaria alternata. Biol. Control 2021, 156, 104539. [Google Scholar] [CrossRef]
- Mironenka, J.; Różalska, S.; Soboń, A.; Bernat, P. Trichoderma harzianum metabolites disturb Fusarium culmorum metabolism: Metabolomic and proteomic studies. Microbiol. Res. 2021, 249, 126770. [Google Scholar] [CrossRef] [PubMed]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Alabouvette, C.; Schippers, B.; Lemanceau, P.; Bakker, P.A.H.M. Biological Control of Fusarium Wilts. Towards Development of Commercial Products. In Plant-Microbe Interactions and Biological Control; Boland, G.J., Kuykendall, L.D., Eds.; M. Dekker Inc.: New York, NY, USA, 1998; pp. 15–36. ISBN 9780824700430. [Google Scholar]
- Mayo, S.; Gutiérrez, S.; Malmierca, M.G.; Lorenzana, A.; Campelo, M.P.; Hermosa, R.; Casquero, P.A. Influence of Rhizoctonia solani and Trichoderma spp. in growth of bean (Phaseolus vulgaris L.) and in the induction of plant defense-related genes. Front. Plant Sci. 2015, 6, 685. [Google Scholar] [CrossRef] [Green Version]
- Ji, S.; Liu, Z.; Wang, Y. Trichoderma-induced ethylene responsive factor MsERF105 mediates defense responses in Malus sieversii. Front. Plant Sci. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [Green Version]
- Chowdappa, P.; Kumar, S.P.M.; Lakshmi, M.J.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Kakabouki, I.; Tataridas, A.; Mavroeidis, A.; Kousta, A.; Karydogianni, S.; Zisi, C.; Kouneli, V.; Konstantinou, A.; Folina, A.; Konstantas, A.; et al. Effect of colonization of Trichoderma harzianum on growth development and CBD content of hemp (Cannabis sativa L.). Microorganisms 2021, 9, 518. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Ruocco, M.; Woo, S.; Lorito, M. Trichoderma secondary metabolites that affect plant metabolism. Nat. Prod. Commun. 2012, 7, 1545–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced Systemic Resistance and Plant Responses to Fungal Biocontrol Agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Singh, S.P.; Pandey, S.; Mishra, N.; Giri, V.P.; Mahfooz, S.; Bhattacharya, A.; Kumari, M.; Chauhan, P.; Verma, P.; Nautiyal, C.S.; et al. Supplementation of Trichoderma improves the alteration of nutrient allocation and transporter genes expression in rice under nutrient deficiencies. Plant Physiol. Biochem. 2019, 143, 351–363. [Google Scholar] [CrossRef]
- Garnica-Vergara, A.; Barrera-Ortiz, S.; Muñoz-Parra, E.; Raya-González, J.; Méndez-Bravo, A.; Macías-Rodríguez, L.; Ruiz-Herrera, L.F.; López-Bucio, J. The volatile 6-pentyl-2 H -pyran-2-one from Trichoderma atroviride regulates Arabidopsis thaliana root morphogenesis via auxin signaling and ETHYLENE INSENSITIVE 2 functioning. New Phytol. 2016, 209, 1496–1512. [Google Scholar] [CrossRef] [Green Version]
- Bader, A.N.; Salerno, G.L.; Covacevich, F.; Consolo, V.F. Native Trichoderma harzianum strains from Argentina produce indole-3 acetic acid and phosphorus solubilization, promote growth and control wilt disease on tomato (Solanum lycopersicum L.). J. King Saud Univ.-Sci. 2020, 32, 867–873. [Google Scholar] [CrossRef]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; De Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stewart, A.; et al. Environmental growth conditions of Trichoderma spp. affects indole acetic acid derivatives, volatile organic compounds, and plant growth promotion. Front. Plant Sci. 2017, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.J. Making greater use of introduced microorganisms for biological control of plant pathogens. Annu. Rev. Phytopathol. 1993, 31, 53–80. [Google Scholar] [CrossRef]
- Porteous-Álvarez, A.J.; Mayo-Prieto, S.; Carro-Huerga, G.; Rodríguez-González, Á.; Álvarez-García, S.; del Ser, S.; Lorenzana, A.; Campelo, M.P.; Maldonado-González, M.M.; Gutiérrez, S.; et al. Trichoderma species isolated from hop soils in the Órbigo valley, León, Spain. Acta Hortic. 2021, 1328, 63–66. [Google Scholar] [CrossRef]
- Tamandegani, P.R.; Marik, T.; Zafari, D.; Balázs, D.; Vágvölgyi, C.; Szekeres, A.; Kredics, L. Changes in peptaibol production of Trichoderma species during in vitro antagonistic interactions with fungal plant pathogens. Biomolecules 2020, 10, 730. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y.; Chet, I.; Boyle, P.; Henis, Y. Parasitism of Trichoderma spp. on Rhizoctonia solani and Sclerotium rolfsii—Scanning electron microscopy and fluorescense microscopy. Phytopathology 1983, 73, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Mayo-Prieto, S.; Campelo, M.P.; Lorenzana, A.; Rodríguez-González, A.; Reinoso, B.; Gutiérrez, S.; Casquero, P.A. Antifungal activity and bean growth promotion of Trichoderma strains isolated from seed vs soil. Eur. J. plant Pathol. 2020, 158, 817–828. [Google Scholar] [CrossRef]
- Anees, M.; Tronsmo, A.; Edel-Hermann, V.; Hjeljord, L.G.; Héraud, C.; Steinberg, C. Characterization of field isolates of Trichoderma antagonistic against Rhizoctonia solani. Fungal Biol. 2010, 114, 691–701. [Google Scholar] [CrossRef]
- Campelo, M.P.; Cardoza, R.E.; Lorenzana, A.; Hermosa, M.R.; Monte, E.; Reinoso, B.; Gutiérrez, S.; Casquero, P.A. Biological control of phytopathogenic fungi in bean (Phaseolus vulgaris L.) with Trichoderma atroviride and Trichoderma virens. Bean Improv. Coop. 2010, 53, 114–115. [Google Scholar]
- Ruano-Rosa, D.; Del Moral-Navarrete, L.; Lopez-Herrera, C.J. Selection of Trichoderma spp. isolates antagonistic to Rosellinia necatrix. Span. J. Agric. Res. 2010, 8, 1084. [Google Scholar] [CrossRef]
- Díaz, G.; Córcoles, A.I.; Asencio, A.D.; Torres, M.P. In vitro antagonism of Trichoderma and naturally occurring fungi from elms against Ophiostoma novo-ulmi. For. Pathol. 2012, 43, 51–58. [Google Scholar] [CrossRef]
- Sánchez-García, B.M.; Espinosa-Huerta, E.; Villordo-Pineda, E.; Rodríguez-Guerra, R.; Mora-Avilés, M.A. Trichoderma spp. native strains molecular identification and in vitro antagonistic evaluation of root phytopathogenic fungus of the common bean (Phaseolus vulgaris L.) cv. Montcalm. Agrociencia 2017, 51, 63–79. [Google Scholar]
- Vinale, F.; Ghisalberti, E.L.; Sivasithamparam, K.; Marra, R.; Ritieni, A.; Ferracane, R.; Woo, S.; Lorito, M. Factors affecting the production of Trichoderma harzianum secondary metabolites during the interaction with different plant pathogens. Lett. Appl. Microbiol. 2009, 48, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites—Strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Horwitz, B.A.; Kenerley, C.M. Secondary metabolism in Trichoderma—A genomic perspective. Microbiology 2012, 158, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mona, S.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Egamberdieva, D. Increased resistance of drought by Trichoderma harzianum fungal treatment correlates with increased secondary metabolites and proline content. J. Integr. Agric. 2017, 16, 1751–1757. [Google Scholar] [CrossRef]
- Calvet, C.; Pera, J.; Barea, J.M. Growth response of marigold (Tagetes erecta L.) to inoculation with Glomus mosseae, Trichoderma aureoviride and Pythium ultimum in a peat-perlite mixture. Plant Soil 1993, 148, 1–6. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Mahmood, I. Biological control of Heterodera cajani and Fusarium udum on pigeonpea by Glomus mosseae, Trichoderma harzianum, and Verticillium chlamydosporium. Isr. J. Plant Sci. 1996, 44, 49–56. [Google Scholar] [CrossRef]
- Saldajeno, M.G.B.; Chandanie, W.A.; Kubota, M.; Hyakumachi, M. Effects of Interactions of Arbuscular Mycorrhizal Fungi and Beneficial Saprophytic Mycoflora on Plant Growth and Disease Protection. In Mycorrhizae: Sustainable Agriculture and Forestry; Siddiqui, Z.A., Akhtar, M.S., Futai, K., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 211–226. [Google Scholar] [CrossRef]
- Duffy, B.K.; Simon, A.; Weller, D.M. Combination of Trichoderma koningii with fluorescent pseudomonads for control of talk-all on wheat. Phytopathology 1996, 86, 188–194. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
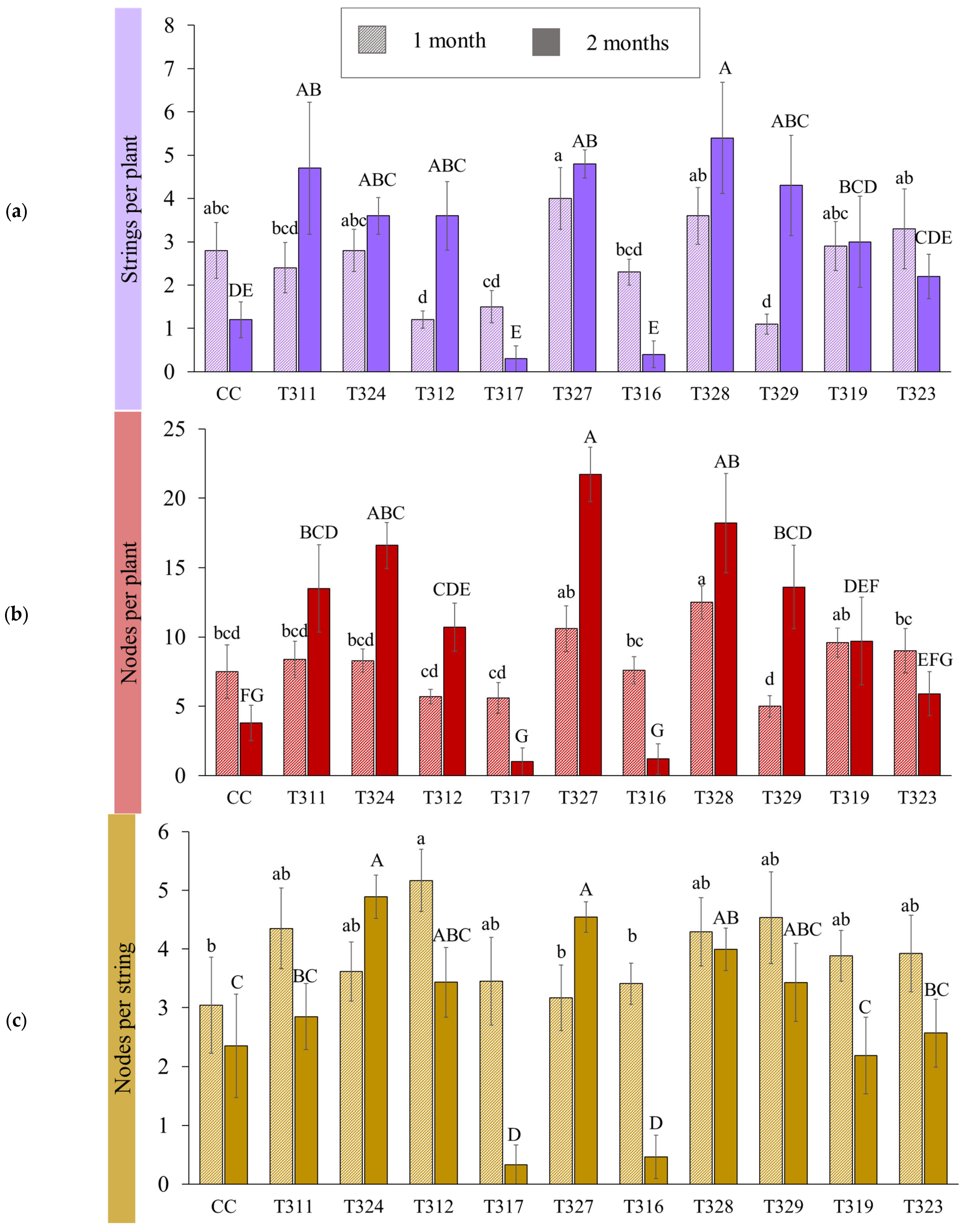{kind=link}
| ID Treatment | Identified as | % Identity | Origin | Location | Accession Number |
|---|---|---|---|---|---|
| T311 | T. hamatum1 | >99% | Soil | Gavilanes de Órbigo | OQ590011 |
| T324 | T. hamatum1 | >99% | Rootstock | San Román de la Vega | OQ589841 |
| T312 | T. virens1 | >99% | Soil | Gavilanes de Órbigo | OQ590010 |
| T317 | T. virens1 | >99% | Rootstock | Nistal | OQ589506 |
| T327 | T. gamsii1 | >99% | Soil | Seisón de la Vega | OQ589844 |
| T316 | T. rossicum1 | >99% | Rootstock | Gavilanes de Órbigo | OQ589503 |
| T328 | T. rossicum1 | >99% | Soil | Gavilanes de Órbigo | OQ589861 |
| T329 | T. harzianum1 | >99% | Soil | San Román de la Vega | OQ589868 |
| T314 | T. spirale1 | >99% | Soil | Nistal | OQ589492 |
| T319 | T. spirale1 | >99% | Rootstock | Nistal | OQ589706 |
| T323 | T. brevicompactum1 | >99% | Soil | San Román de la Vega | OQ589712 |
| F076 | F. culmorum2 | >99% | Soil | San Román de la Vega | OQ625436 |
| F103 | F. oxysporum2 | >99% | Soil | San Román de la Vega | OQ632904 |
| F079 | F. sambucinum2 | >99% | Rootstock | San Román de la Vega | OQ632903 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porteous-Álvarez, A.J.; Fernández-Marcos, A.; Ramírez-Lozano, D.; Mayo-Prieto, S.; Cardoza, R.E.; Gutiérrez, S.; Casquero, P.A. Native Trichoderma Isolates from Soil and Rootstock to Fusarium spp. Control and Growth Promotion of Humulus lupulus L. Plantlets. Agriculture 2023, 13, 720. https://doi.org/10.3390/agriculture13030720
Porteous-Álvarez AJ, Fernández-Marcos A, Ramírez-Lozano D, Mayo-Prieto S, Cardoza RE, Gutiérrez S, Casquero PA. Native Trichoderma Isolates from Soil and Rootstock to Fusarium spp. Control and Growth Promotion of Humulus lupulus L. Plantlets. Agriculture. 2023; 13(3):720. https://doi.org/10.3390/agriculture13030720
Chicago/Turabian StylePorteous-Álvarez, Alejandra J., Alexia Fernández-Marcos, Daniela Ramírez-Lozano, Sara Mayo-Prieto, Rosa E. Cardoza, Santiago Gutiérrez, and Pedro A. Casquero. 2023. "Native Trichoderma Isolates from Soil and Rootstock to Fusarium spp. Control and Growth Promotion of Humulus lupulus L. Plantlets" Agriculture 13, no. 3: 720. https://doi.org/10.3390/agriculture13030720