
Behavioral Response of a Specialist Predator, Stethorus gilvifrons (Coccinellidae: Coleoptera), to Cope with Web Structures of Four Different Spider Mite Pest Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Pest Mite Sources
- Date palm mite (DPM), O. afrasiaticus, belonging to the life type CW–d, collected from infested date palm trees grown in the vicinity of the King Saud University campus;
- Two-spotted spider mite (TSSM), T. urticae, belonging to the life type CW–u, collected from infested eggplant plants sown in the greenhouses at King Saud University;
- Citrus brown mite (CBM), E. orientalis, belonging to the life type LW, collected from infested citrus plants grown in Education Farm at King Saud University;
- Avocado brown mite (ABM), O. punicae, belonging to the life type CW–c, collected from infested Conocarpus spp. present in the proximity of the King Saud University campus.
2.2. Predator Source
2.3. Experimental Procedure
2.4. Data Recording of Stethorus Gilvifrons Response to Pest Mite Webbing
2.5. Data Analysis
2.5.1. Behavioral Categories
2.5.2. Statistical Analysis
3. Results
3.1. Observations on Web Coping Behavior of Stethorus Gilvifrons
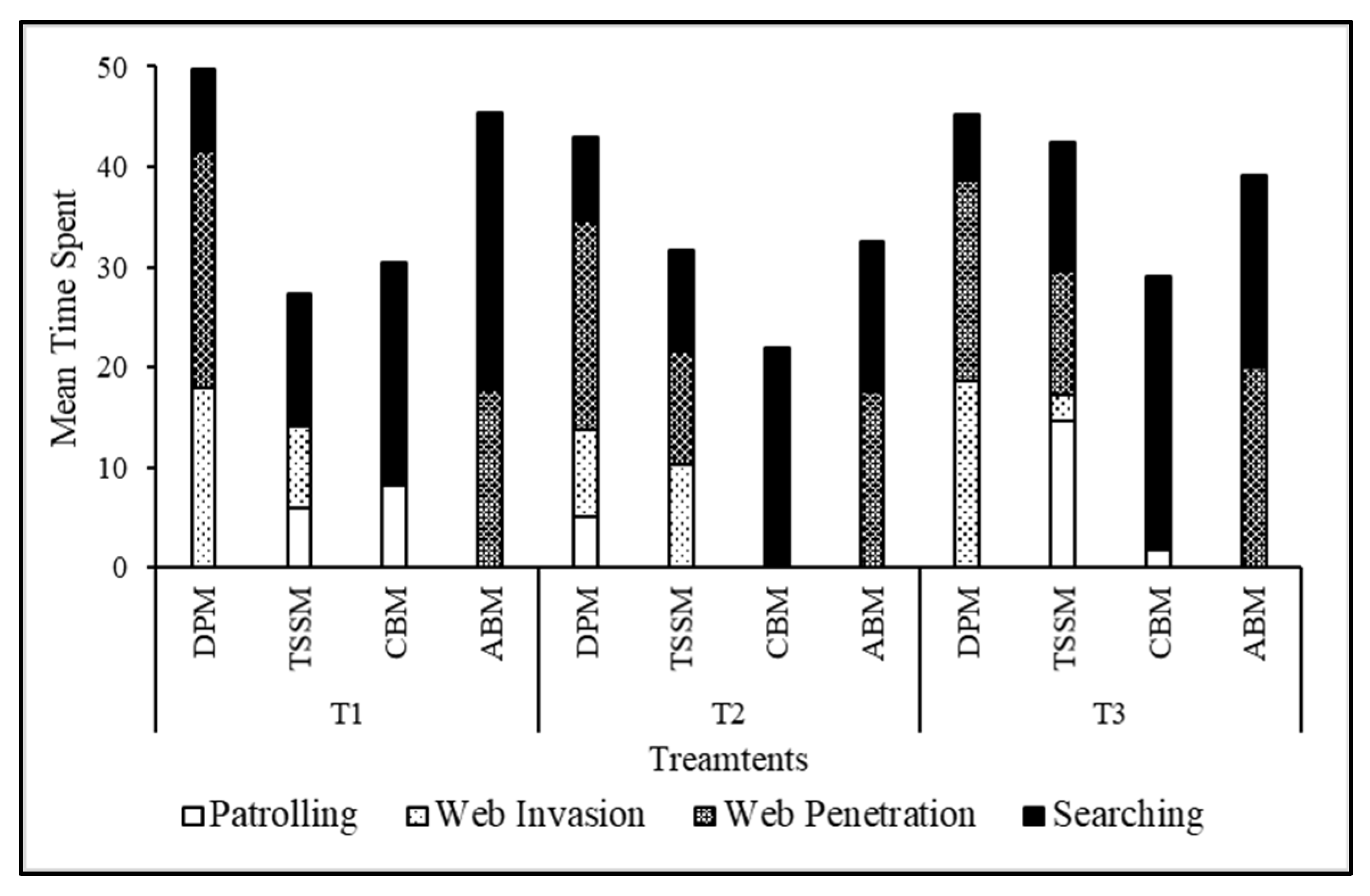
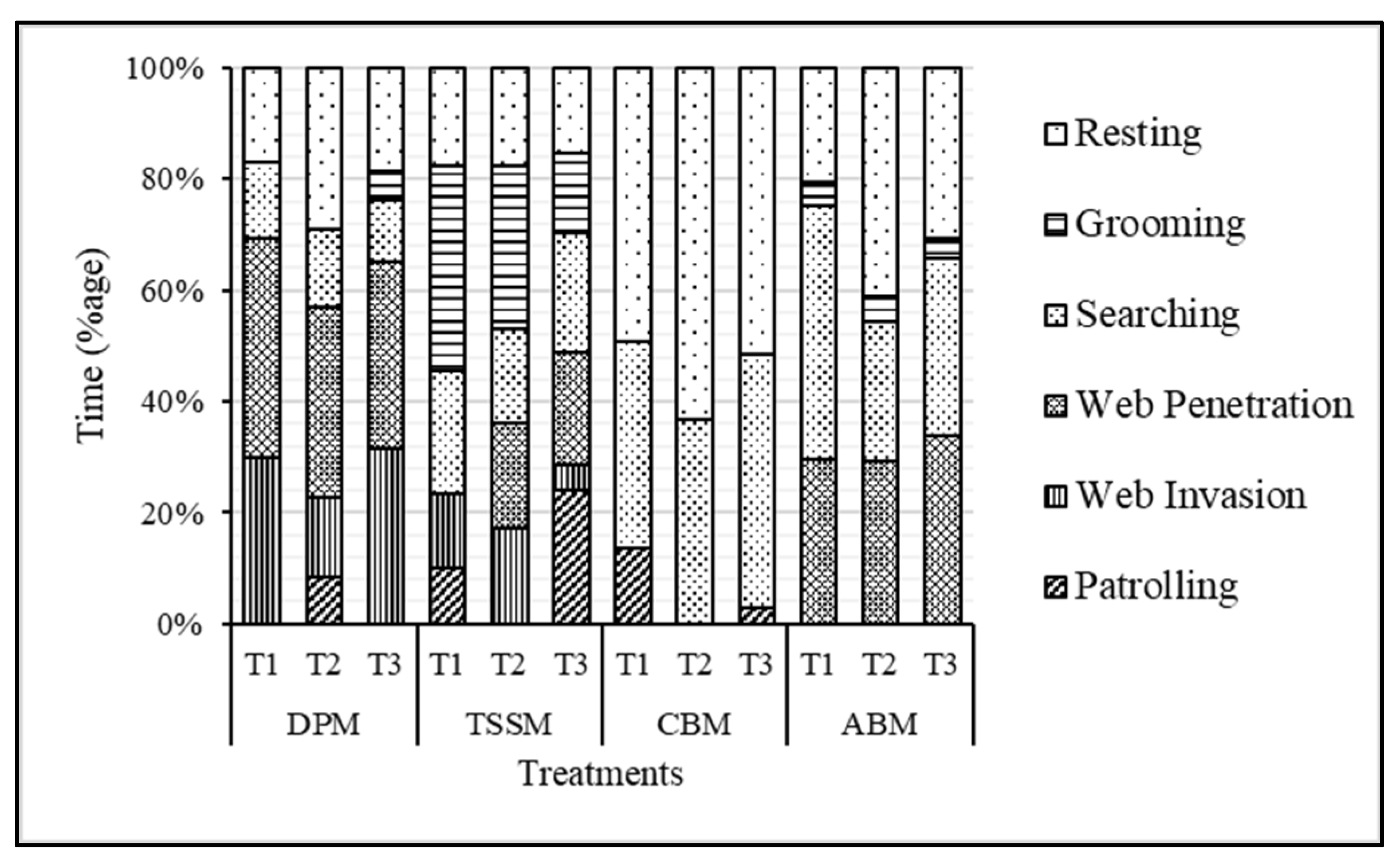
3.2. Attack Behaviors and Predation efficiency of Stethorus Gilvifrons against Webbing of Four Spider Mite Pests
3.3. Behavioral Response of Stethorus Gilvifrons against Webbing of Four Spider Mite Pests
3.3.1. Comparison of Predator Behavior among Treatments:
3.3.2. Comparison of Behavior among Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biddinger, D.J.; Weber, D.C.; Hull, L.A. Coccinellidae as predators of mites: Stethorini in biological control. Biol. Control 2009, 51, 268–283. [Google Scholar] [CrossRef]
- Lundgren, J.G. Nutritional aspects of non-prey foods in the life histories of predaceous Coccinellidae. Biol. Control 2009, 51, 294–305. [Google Scholar] [CrossRef]
- Helle, W.; Sabelis, M.W. Spider Mites: Their Biology, Natural Enemies, and Control; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1A, pp. 1–405. [Google Scholar]
- Osman, M.A. Functional response of the coccinellid predator Stethorus gilvifrons Mulsant feeding on the two spotted spider mite, Tetranychus urticae Koch: The effect of different prey stages. J. Plant Protect. Pathol. 2010, 1, 753–765. [Google Scholar] [CrossRef]
- Kundoo, A.A.; Khan, A.A. Coccinellids as biological control agents of soft bodied insects: A review. J. Entomol. Zool. Stud. 2017, 5, 1362–1373. [Google Scholar]
- Fournier, V.; Rosenheim, J.A.; Johnson, M.W.; Brodeur, J. Augmentative releases of predatory mites on papaya in Hawaii: Failures and success. In Proceedings of the 1st International Symposium on Biological Control of Arthropods, Honolulu, HI, USA, 14–18 January 2002. [Google Scholar]
- Hoy, M.A. Agricultural Acarology: Introduction to Integrated Mite Management; CRC Press: Boca Raton, FL, USA, 2011; Volume 7. [Google Scholar]
- Saito, Y. Plant Mites and Sociality, Diversity and Evolution; Springer: Tokyo, Japan, 2010. [Google Scholar]
- Oku, K.; Yano, S.; Takafuji, A. Nonlethal indirect effects of a native predatory mite, Amblyseius womersleyi Schicha (Acari: Phytoseiidae), on the phytophagous mite Tetranychus kanzawai Kishida (Acari: Tetranychidae). J. Ethol. 2004, 22, 109–112. [Google Scholar] [CrossRef]
- Saito, Y. The concept of “life types” in Tetranychinae. An attempt to classify the spinning behavior of Tetranychinae. Acarologia 1983, 24, 377–391. [Google Scholar]
- Saito, Y. Life types of spider mites. In Spider Mites: Their Biology, Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1A, pp. 253–264. [Google Scholar]
- Mirza, J.H.; Kamran, M.; Alatawi, F.J. Webbing life type and behavioral response of the date palm mite, Oligonychus afrasiaticus, to webbing residues on leaves and fruits of date palm. Exp. Appl. Acarol. 2018, 76, 197–207. [Google Scholar] [CrossRef]
- Mushtaq, H.M.S.; Kamran, M.; Alatawi, F.J. Two new life types and assessment of web-associated behavioral characteristics of some Oligonychus species on various host plants. Exp. Appl. Acarol. 2021, 83, 211–227. [Google Scholar] [CrossRef]
- Iwasa, T.; Osakabe, M. Effects of combination between web density and size of spider mite on predation by a generalist and a specialist phytoseiid mite. Exp. Appl. Acarol. 2015, 66, 219–225. [Google Scholar] [CrossRef]
- Trichilo, P.J.; Leigh, T.F. Predation on spider mite eggs by the western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), an opportunist in a cotton agroecosystem. Environ. Entomol. 1986, 15, 821–825. [Google Scholar] [CrossRef]
- Osakabe, M. Relationships between food substances and developmental success in Amblyseius sojaensis Ehara (Acarina: Phytoseiidae). Appl. Entomol. Zool. 1988, 23, 45–51. [Google Scholar] [CrossRef]
- Shimoda, T.; Kishimoto, H.; Takabayashi, J.; Amano, H.; Dicke, M. Comparison of thread-cutting behavior in three specialist predatory mites to cope with complex webs of Tetranychus spider mites. Exp. Appl. Acarol. 2008, 47, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Mirza, J.H.; Kamran, M.; Alatawi, F.J. Response of the predatory mite Cydnoseius negevi (Acari: Phytoseiidae) to webbing of the date palm mite, Oligonychus afrasiaticus (Acari: Tetranychidae), on date palm fruits and leaves. Exp. Appl. Acarol. 2018, 75, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Croft, B.A.; Blackwood, J.S.; McMurtry, J.A. Classifying life-style types of phytoseiid mites: Diagnostic traits. Exp. Appl. Acarol. 2004, 33, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, T.; Kishimoto, H.; Amano, H.; Takabayashi, J. Behavioral adaptation of predatory mites to move within complex webs of spider mites. In Abstracts of the Annual Meeting of the Japanese Society of Applied Entomology and Zoology; J-Stage: Saitama, Japan, 2003; Volume 47, p. 110. [Google Scholar]
- Afshari, M.; Mossadegh, S.; Soleyman-Nejadian, E.; Kamali, K. Geographical distribution and host plants of Stethorus gilvifrons (Mulsant) (Col.: Coccinellidae) and its biology under laboratory conditions in Khuzestan province. J. Agric. Sci. Nat. Res. 2007, 14, 201–210. [Google Scholar]
- El-Adawy, A.M.; Yousri, H.; Ahmed, Y.M.; Tiilikkala, K.; El-Sharkawy, T.A. Estimation of general selective toxicity ratios of certain acaricides to Stethorus gilvifrons (Mulsant) and its prey Tetranychus urticae Koch. Egypt. J. Agric. Res. 2000, 78, 1081–1089. [Google Scholar]
- Sohrabi, F.; Shishehbor, P. Functional and numerical responses of Stethorus gilvifrons Mulsant feeding on strawberry spider mite, Tetranychus turkestani Ugarov and Nikolski. Pak. J. Biol. Sci. 2007, 10, 4563–4566. [Google Scholar] [CrossRef]
- Imani, Z.; Shishehbor, P.; Sohrabi, F. The effect of Tetranychus turkestani and Eutetranychus orientalis (Acari: Tetranychidae) on the development and reproduction of Stethorus gilvifrons (Coleoptera: Coccinellidae). J. Asia-Pacific. Entomol. 2009, 12, 213–216. [Google Scholar] [CrossRef]
- Handoko, H.; Affandi, A. Life-history traits of Stethorus gilvifrons (Mulsant) (Coleoptera: Coccinellidae) on phytophagous mites Eutetranychus orientalis Klein (Acari: Tetranychidae). J. Agric. Sci. 2012, 34, 7–13. [Google Scholar] [CrossRef]
- Bayoumy, M.H.; Osman, M.A.; Michaud, J.P. Host plant mediates foraging behavior and mutual interference among adult Stethorus gilvifrons (Coleoptera: Coccinellidae) preying on Tetranychus urticae (Acari: Tetranychidae). Environ. Entomol. 2014, 43, 1309–1318. [Google Scholar] [CrossRef]
- Jafari, M.; Ranjbar, A.H.; Goldasteh, S.; Zamani, A.; Soleiman, N.E. Effects of Prey Species and Host Plants on Development and Life History of Stethorus gilvifrons (Coleoptera: Coccinellidae). J. Agric. Sci. Tech. 2022, 24, 635–648. [Google Scholar]
- van Lenteren, J.C. Biological pest control in greenhouses: An overview. Arab J. Plant Prot. 1992, 10, 35–43. [Google Scholar]
- Ahmad, M.; Mofleh, M.; Haloum, M. The efficiency of the predator Stethorus gilvifrons Mulsant to control the two spotted spider mite Tetranychus urticae Koch in greenhouse eggplant. Arab J. Plant Prot. 2010, 28, 169–174. [Google Scholar]
- Afshari, G.A. A Survey on Ladybirds Belong to Genus Stethorus and Study on Biology, Prey Consumption and Population Dynamics of Stethorus gilvifrons in Sugarcane Farm in Khuzestan, Iran. Master’s Thesis, Ahwaz, Iran,, 1999. [Google Scholar]
- Almatni, W.; Elabdulla, J. The beetle Stethorus gilvifrons, a predator of mites which attack apple orchards in southern Syria. Arab Near East Plant Prot. Newsl. 2000, 31, 29. [Google Scholar]
- Ibrahim, H.M. Field experiment of the predacious insect, Stethorus gilvifrons Mulsant releases to evaluate the predation efficacy in controlling europern red mite and two-spotted spider mite on peach trees in Egypt. In Proceedings of the 1st Conference on Safe Alternatives of Pesticides for Pest Management, Assiut, Egypt, 28–29 October 2001. [Google Scholar]
- Ibrahim, G.A.; Metwally, A.M.; Zakzouk, E.A.; El-Halawany, A.S.H. Biological control of the two-spotted spider mite and the European red mite using the predatory insect, Stethorus gilvifrons Mulsant, coccinellidae, coleoptera, on apple seedlings. Egypt. J. Agric. Res. 2010, 88, 359–368. [Google Scholar]
- Saito, Y.; Lin, J.Z.; Zhang, Y.X.; Ito, K.; Liu, Q.Y.; Chittenden, A.R. Two new species and four new life types in Tetranychidae. Ann. Entomol. Soc. Am. 2016, 109, 463–472. [Google Scholar] [CrossRef]
- Montserrat, M.; Peña, F.D.I.; Hormaza, J.I.; González-Fernández, J.J. How do Neoseiulus californicus (Acari: Phytoseiidae) females penetrate densely webbed spider mite nests. Exp. Appl. Acarol. 2008, 44, 101–106. [Google Scholar] [CrossRef]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Hazan, A.; Gerson, U.; Tahori, A.S. Spider mite webbing. I. The production of webbing under various environmental conditions. Acarologia 1974, 16, 68–84. [Google Scholar]
- Gerson, U.; Aronowitz, A. Spider mite webbing. V. The effect of various host plants. Acarologia 1981, 22, 277–281. [Google Scholar]
- Roda, A.; Nyrop, J.; Dicke, M.; English-Loeb, G. Trichomes and spider-mite webbing protect predatory mite eggs from intraguild predation. Oecologia 2000, 125, 429–436. [Google Scholar] [CrossRef]
- Loughner, R.; Goldman, K.; Loeb, G.; Nyrop, J. Influence of leaf trichomes on predatory mite (Typhlodromus pyri) abundance in grape varieties. Exp. Appl. Acarol. 2008, 45, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Houck, M.A. Time and resource partitioning in Stethorus punctum (Coleoptera: Coccinellidae). Environ. Entomol. 1991, 20, 494–497. [Google Scholar] [CrossRef]
- Tanigoshi, L.K.; McMurtry, J.A. The dynamics of predation of Stethorus picipes (Coleoptera: Coccinellidae) and Typhlodromus floridanus on the prey Oligonychus punicae (Acarina: Phytoseiidae, Tetranychidae). Hilgardia 1977, 45, 237–288. [Google Scholar] [CrossRef]
- Latifian, M. Voracity and feeding preferences of larvae and adult stages of Stethorus gilvifrons Mulsant (Coleoptera: Coccinellidae) on larvae and adult of Oligonychus afrasiaticus McGregor (Acarina: Tetranychidae). Int. J. Agric. Crop Sci. 2012, 4, 540–546. [Google Scholar]
- Latifian, M.; Kajbafvala, G. Studying the Efficiency of Stethorus gilvifrons Inoculate Release for Date Palm Spider Mite (Oligonychus afrasiaticus) Biological Control in Field Condation; Ministry of Jahad-E-Agriculture, Research and education organization of Iran, Date Palm and Tropical Fruits Research Institute of Iran: Khuzestan, Iran, 2017; 75p. [Google Scholar]
- Takano-Lee, M.; Hoddle, M. Predatory behaviors of Neoseiulus californicus and Galendromus helveolus (Acari: Phytoseiidae) attacking Oligonychus perseae (Acari: Tetranychidae). Exp. Appl. Acarol. 2010, 26, 13–26. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patrolling | Web Invasion | Web Penetration | Searching | Grooming | Resting | ||
|---|---|---|---|---|---|---|---|
| DPM | T1 | - (0) | 16.6 ± 7.3 b (18 ± 7.6) | 14.3 ± 8.4 (23.5 ± 6.7) | 14.1 ± 7.1 (8.3 ± 5.1) | - (0) | 9.4 ± 6.8 (10.1 ± 3.7) |
| T2 | 17.5 ± 6.3 (5.1 ± 4.4) | 5.8 ± 3.3 a (8.7 ± 5.5) | 11.6 ± 7.7 (20.7 ± 7.2) | 12.7 ± 7.4 (8.4 ± 4.5) | - (0) | 17.6 ± 5.6 (17.6 ± 8) | |
| T3 | - (0) | 15 ± 4 ab (18.6 ± 1.8) | 11.5 ± 5.1 (20 ± 3.6) | 10.6 ± 6.9 (6.6 ± 4.3) | 20.5 ± 2.1 (3 ± 4.5) | 10.3 ± 6.5 (11 ± 3.2) | |
| STAT | DF = 2, p < 0.0001 | DF = 2, p = 0.004 | DF = 2, p = 0.65 | DF = 2, p = 0.6 | DF = 2, p < 0.0001 | DF = 2, p = 0.03 | |
| ABM | T1 | - | - | 9.9 ± 7.6 (17.7 ± 5.6) | 18.7 ± 5.7 b (27.6 ± 8.1) | 13.5 ± 9.4 (2.5 ± 1.1) | 5 ± 3.2 a (12.3 ± 5.4) |
| T2 | - | - | 11.6 ± 8.2 (17.6 ± 5.9) | 7.3 ± 5.9 a (15 ± 6.1) | 14.1 ± 6.5 (2.7 ± 1.3) | 19.4 ± 4.1 b (24.6 ± 5.2) | |
| T3 | - | - | 15.9 ± 3.7 (20 ± 1.5) | 11.4 ± 4.1 ab (19.1 ± 2.9) | 9.8 ± 3.6 (2 ± 0.7) | 13.1 ± 4.1 ab (18.2 ± 2.1) | |
| STAT | - | - | DF = 2, p = 0.21 | DF = 2, p = 0.004 | DF = 2, p = 0.41 | DF = 2, p = 0.0001 | |
| TSSM | T1 | 13.2 ± 4.3 b (6.3 ± 4) | 13.4 ± 6.3 (8.13 ± 5.1) | 0 a (0) | 12.1 ± 6.4 (13.2 ± 3.3) | 4.5 ± 2.3 b (22 ± 3.5) | 13.3 ± 7.7 (10.6 ± 7.1) |
| T2 | 0 a (0) | 14.8 ± 9.5 (10.3 ± 8.8) | 15.1 ± 5.7 b (11.3 ± 4.6) | 10.3 ± 6.4 (10.1 ± 6.6) | 13 ± 2.8 b (17.7 ± 7.6) | 11.8 ± 5.9 (10.5 ± 4.6) | |
| T3 | 17.7 ± 7.1 b (14.6 ± 9.3) | 9.2 ± 3.1 (2.7 ± 1.6) | 15.9 ± 6.9 b (12.2 ± 6.1) | 15.1 ± 8.1 (13 ± 8.2) | 20 ± 3.1 a (8.6 ± 6) | 11.5 ± 7.5 (9.2 ± 6) | |
| STAT | DF = 2, p = 0.0001 | DF = 2, p = 0.23 | DF = 2, p = 0.0001 | DF = 2, p = 0.4 | DF = 2, p < 0.0001 | DF = 2, p = 0.77 | |
| CBM * | T1 | 17.5 ± 7.5 b (8.1 ± 5.5) | - | - | 10.1 ± 7.6 (22.2 ± 3.9) | - | 7.1 ± 3.9 a (29.6 ± 2.5) |
| T2 | 0 a (0) | - | - | 9.2 ± 4.9 (22 ± 2) | - | 20.1 ± 2.9 b (38 ± 2) | |
| T3 | 14.5 ± 2.2 b (1.7 ± 0.7) | - | - | 18.1 ± 4.9 (27.2 ± 3.9) | - | 10.2 ± 5.7 a (31 ± 4.4) | |
| STAT | DF = 2, p = 0.0001 | - | - | DF = 2, p = 0.02 | - | DF = 2, p = 0.001 |
| Patrolling | Web Invasion | Web Penetration | Searching | Grooming | Resting | ||
|---|---|---|---|---|---|---|---|
| T1 | DPM | 0 a (0) | 25.5 ± 10.1 b (18 ± 7.6) | 26.3 ± 4.7 b (23.5 ± 6) | 6.4 ± 5.1 a (8.7 ± 5.1) | 0 a (0) | 10.1 ± 6.1 a (10.1 ± 3.6) |
| ABM | 0 a (0) | 0 a (0) | 22.6 ± 4.2 b (17.7 ± 5.6) | 26 ± 6 c (27.6 ± 8.1) | 17.5 ± 6.3 ab (2.5 ± 1.1) | 14.5 ± 7 ab (12.1 ± 5.4) | |
| TSSM | 22.8 ± 3.9 b (6 ± 3.4) | 16.8 ± 10.5 ab (8.1 ± 5.1) | 0 a (0) | 11.2 ± 4.4 ab (13.2 ± 3.3) | 28.5 ± 2.2 b (22 ± 3.5) | 13.3 ± 8.8 a (10.6 ± 7.1) | |
| CBM | 26.1 ± 4.9 b (8.1 ± 5.5) | 0 a (0) | 0 a (0) | 22.3 ± 3.9 bc (22.2 ± 3.9) | 0 a (0) | 27.5 ± 2.5 b (29.6 ± 2.5) | |
| STAT | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p = 0.001 | |
| T2 | DPM | 19.5 ± 14.5 b (4.5 ± 4) | 21.7 ± 8.9 b (8.7 ± 5.5) | 24.5 ± 6.9 b (20.7 ± 7.2) | 9.1 ± 5.8 a (8.3 ± 4.5) | 0 a (0) | 12.5 ± 7 a (17.6 ± 8) |
| ABM | 0 a (0) | 0 a (0) | 21.4 ± 6.9 b (17.6 ± 6) | 17.5 ± 8.1 ab (15 ± 6.2) | 20.3 ± 4.9 ab (2.7 ± 1.3) | 18.8 ± 3.1 ab (24.6 ± 5.2) | |
| TSSM | 0 a (0) | 22.2 ± 9.4 b (10.5 ± 8.8) | 13.5 ± 7.7 ab (11.3 ± 7.1) | 11.8 ± 7.1 b (10.1 ± 6.6) | 26.6 ± 6.9 b (17.7 ± 7.6) | 6.2 ± 4 a (10.5 ± 4.6) | |
| CBM | 0 a (0) | 0 a (0) | 0 a (0) | 27.5 ± 2.7 a (22 ± 2) | 0 a (0) | 28.3 ± 2.5 b (38 ± 2) | |
| STAT | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | |
| T3 | DPM | 0 a (0) | 28.5 ± 2.3 b (18.6 ± 1.8) | 24.2 ± 6.7 b (21.2 ± 3.4) | 6.3 ± 2.6 a (6.6 ± 3.4) | 11.8 ± 12.3 ab (3 ± 4.5) | 8.4 ± 4.2 ab (10.5 ± 3.2) |
| ABM | 0 a (0) | 0 a (0) | 22.2 ± 4.3 b (20 ± 1.5) | 18.9 ± 4.5 ab (19.1 ± 2.9) | 19.5 ± 2.2 b (2 ± 0.7) | 20.5 ± 2.5 bc (18.8 ± 2.1) | |
| TSSM | 26.5 ± 5.7 b (14.6 ± 9.4) | 19.5 ± 4.6 ab (2.7 ± 1.6) | 13.1 ± 9.2 ab (12.2 ± 8.4) | 12.1 ± 6.7 a (12.2 ± 7.8) | 23.1 ± 10.4 b (8.6 ± 6) | 8.5 ± 5.4 a (9.2 ± 6.1) | |
| CBM | 22.5 ± 2.2 b (1.7 ± 0.7) | 0 a (0) | 0 a (0) | 28.5 ± 2.4 b (27.2 ± 3.9) | 0 a (0) | 28.4 ± 2.5 c (31 ± 4.4) | |
| STAT | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 |
| DPM | ABM | TSSM | CBM | |||||
|---|---|---|---|---|---|---|---|---|
| During 1 h | After 24 h | During 1 h | After 24 h | During 1 h | After 24 h | During 1 h | After 24 h | |
| T1 | 1.6 ± 0.3 a (2.8 ± 1.1) | 2.1 ± 0.8 a (4.4 ± 0.7) | 1.9 ± 0.3 (3.8 ± 1.1) | 2.2 ± 0.0 a (5 ± 0) | 0.5 ± 0.7 (0.7 ± 1.2) | 0.8 ± 0.9 (1.5 ± 1.8) | 0.7 ± 0.8 (1.1 ± 1.7) | 1.3 ± 0.7 ab (2.2 ± 1.6) |
| T2 | 2.1 ± 0.3 ab (4.8 ± 1.3) | 2.9 ± 0.9 ab (8.8 ± 1.6) | 2.4 ± 0.5 (6.2 ± 2.6) | 2.9 ± 0.3 ab (8.5 ± 1.7) | 0.7 ± 0.6 (1 ± 0.9) | 1.7 ± 1.1 (4.2 ± 2.6) | 0 (0) | 0 a (0) |
| T3 | 2.8 ± 0.3 b (8.3 ± 1.8) | 3.7 ± 0.2 b (14 ± 1.8) | 2 ± 1.1 (5.2 ± 3.8) | 2.8 ± 1.1 b (9.2 ± 4.4) | 1.1 ± 0.5 (1.4 ± 1.1) | 1.8 ± 0.8 (3.5 ± 2.5) | 0.7 ± 0.4 (3 ± 1.8) | 1.5 ± 0.7 b (3 ± 1.8) |
| STAT | DF = 2, p = 0.001 | DF = 2, p < 0.0001 | DF = 2, p = 0.249 | DF = 2, p = 0.004 | DF = 2, p = 0.197 | DF = 2, p = 0.044 | DF = 2, p = 0.018 | DF = 2, p = 0.001 |
| T1 | T2 | T3 | ||||
|---|---|---|---|---|---|---|
| During 1 h | After 24 h | During 1 h | After 24 h | During 1 h | After 24 h | |
| DPM | 1.6 ± 0.3 ab (2.8 ± 1.1) | 2.1 ± 0.8 ab (4.3 ± 0.7) | 2.1 ± 0.3 bc (4.8 ± 1.4) | 2.9 ± 0.2 b (8.7 ± 1.5) | 2.8 ± 0.3 b (8.3 ± 1.8) | 3.7 ± 0.2 b (14 ± 1.8) |
| ABM | 1.9 ± 0.3 b (3.8 ± 1.1) | 2.2 ± 0.0 b (5 ± 0) | 2.4 ± 0.5 c (6.2 ± 2.6) | 2.9 ± 0.3 b (8.5 ± 1.7) | 2 ± 1.1 ab (5.2 ± 3.8) | 2.8 ± 1.1 ab (9.2 ± 4.4) |
| TSSM | 0.5 ± 0.7 a (0.7 ± 1.1) | 0.8 ± 0.9 a (1.5 ± 1.8) | 0.7 ± 0.6 ab (1 ± 0.9) | 1.7 ± 1.1 ab (4.2 ± 2.6) | 1.1 ± 0.7 ab (1.6 ± 0.9) | 1.8 ± 0.8 a (4 ± 2.1) |
| CBM | 0.7 ± 0.8 a (1.1 ± 1.7) | 1.3 ± 0.7 a (2.2 ± 0.6) | 0 a (0) | 0 a (0) | 0.7 ± 0.4 a (0.7 ± 0.4) | 1.5 ± 0.7 a (3 ± 1.8) |
| STAT | DF = 3, p = 0.001 | DF = 3, p = 0.001 | DF = 3, p < 0.0001 | DF = 3, p < 0.0001 | DF = 3, p = 0.001 | DF = 3, p < 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirza, J.H.; Kamran, M.; Alatawi, F.J. Behavioral Response of a Specialist Predator, Stethorus gilvifrons (Coccinellidae: Coleoptera), to Cope with Web Structures of Four Different Spider Mite Pest Species. Agriculture 2023, 13, 582. https://doi.org/10.3390/agriculture13030582
Mirza JH, Kamran M, Alatawi FJ. Behavioral Response of a Specialist Predator, Stethorus gilvifrons (Coccinellidae: Coleoptera), to Cope with Web Structures of Four Different Spider Mite Pest Species. Agriculture. 2023; 13(3):582. https://doi.org/10.3390/agriculture13030582
Chicago/Turabian StyleMirza, Jawwad Hassan, Muhammad Kamran, and Fahad Jaber Alatawi. 2023. "Behavioral Response of a Specialist Predator, Stethorus gilvifrons (Coccinellidae: Coleoptera), to Cope with Web Structures of Four Different Spider Mite Pest Species" Agriculture 13, no. 3: 582. https://doi.org/10.3390/agriculture13030582