
Colletotrichum Species on Cultivated Solanaceae Crops in Russia
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Cultures
2.2. DNA Isolation, PCR, Sequencing, and Phylogenetic Analysis
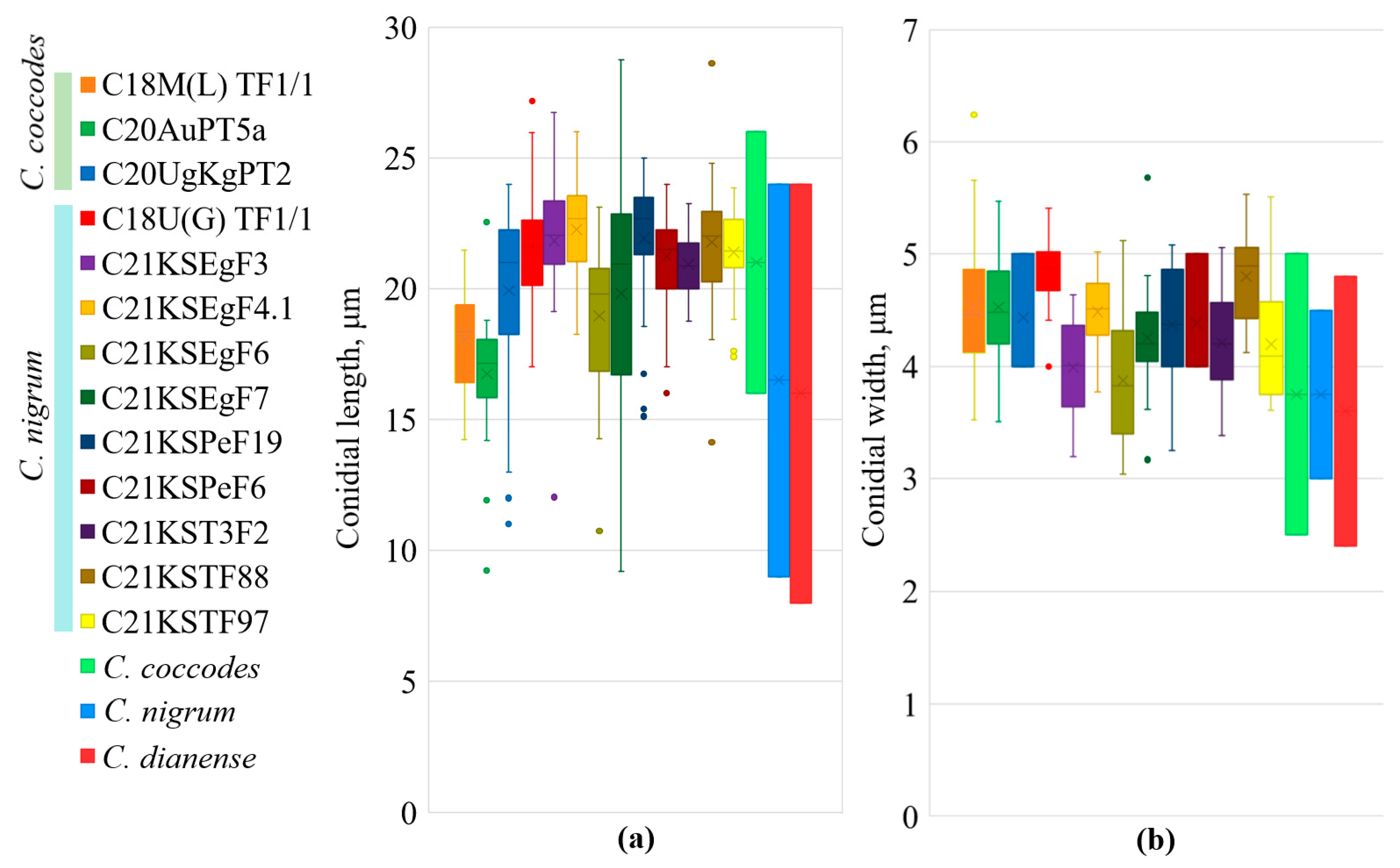
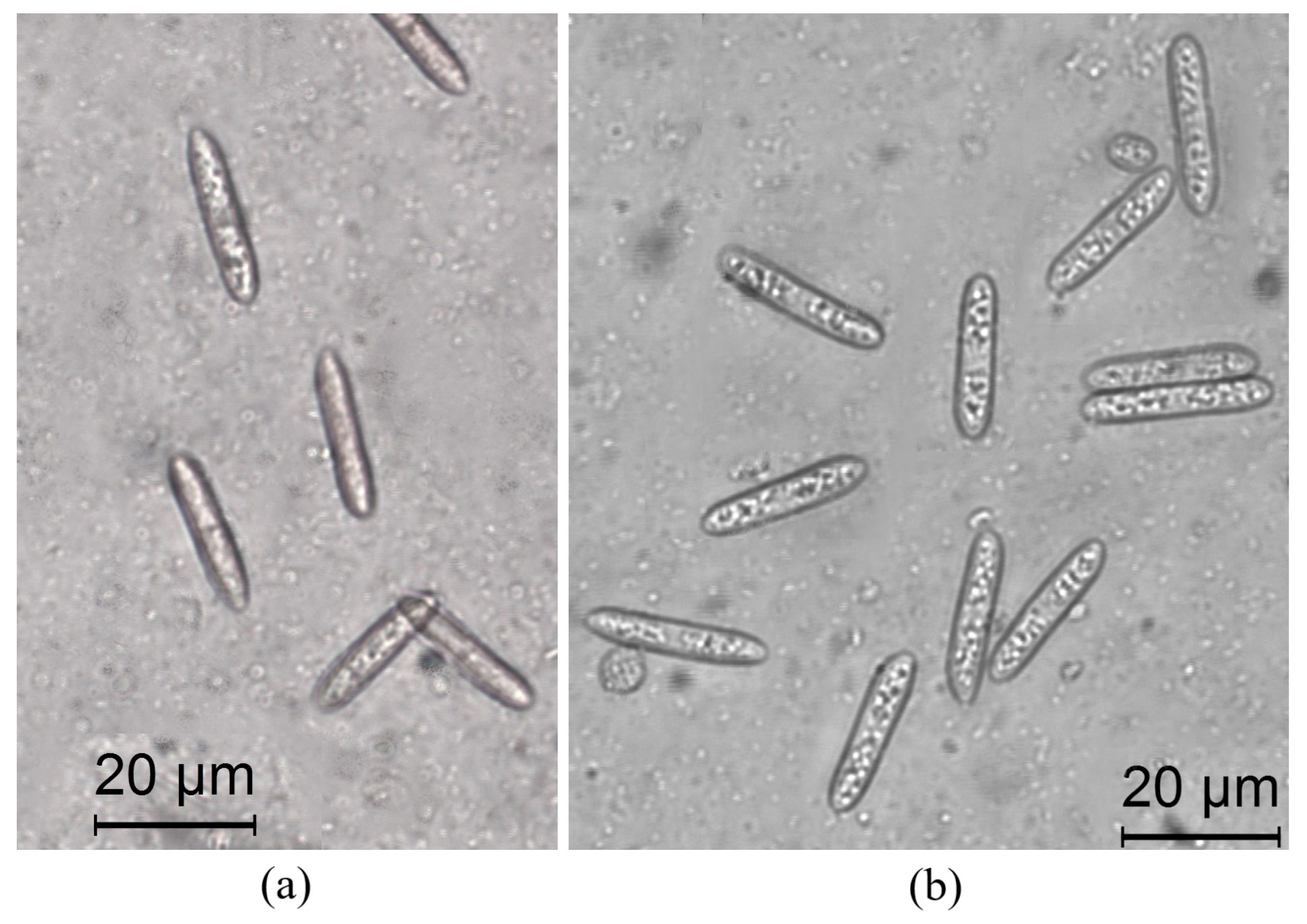
2.3. Morphological Analysis
2.4. Pathogenicity Tests
2.5. In Vitro Assessment of Fungicide Sensitivity
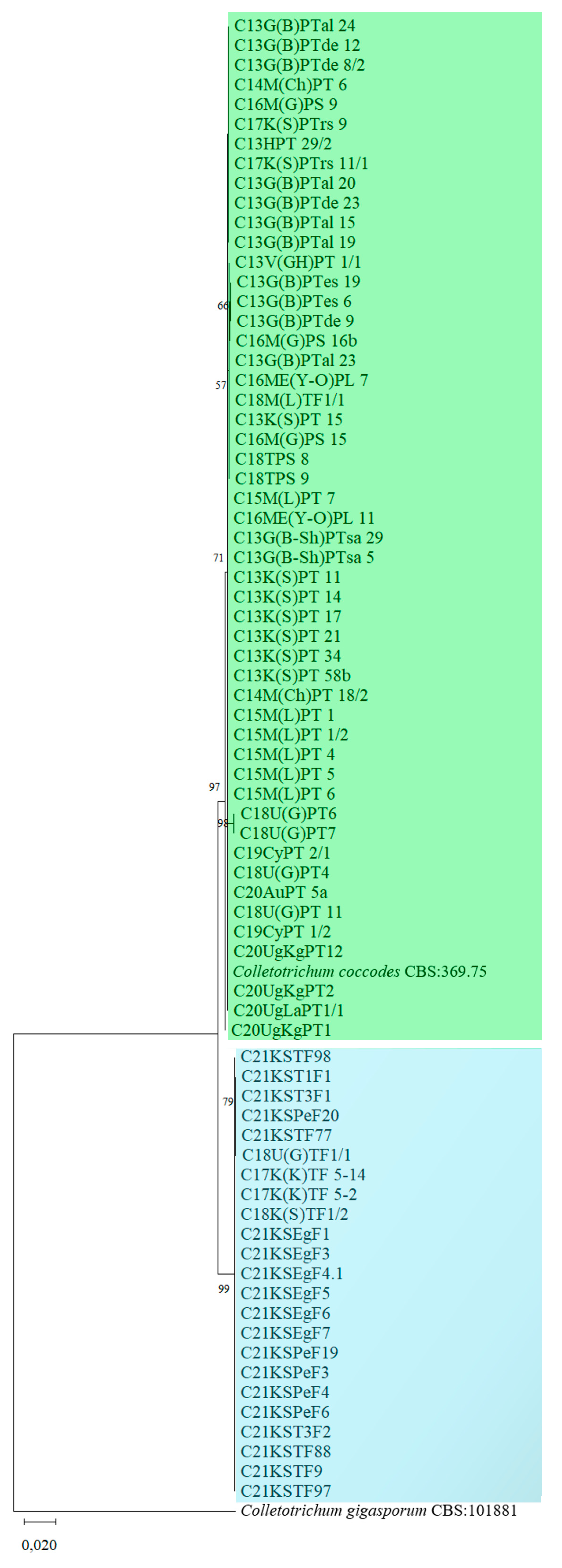
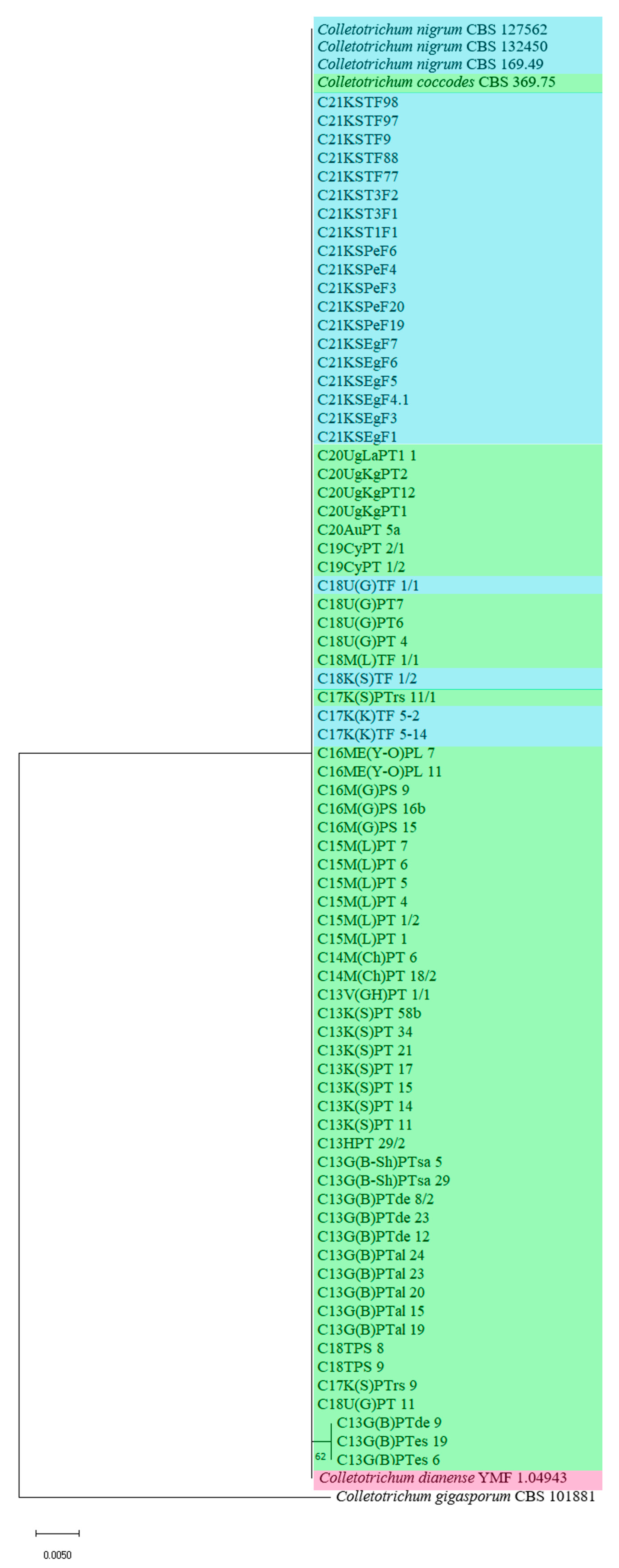
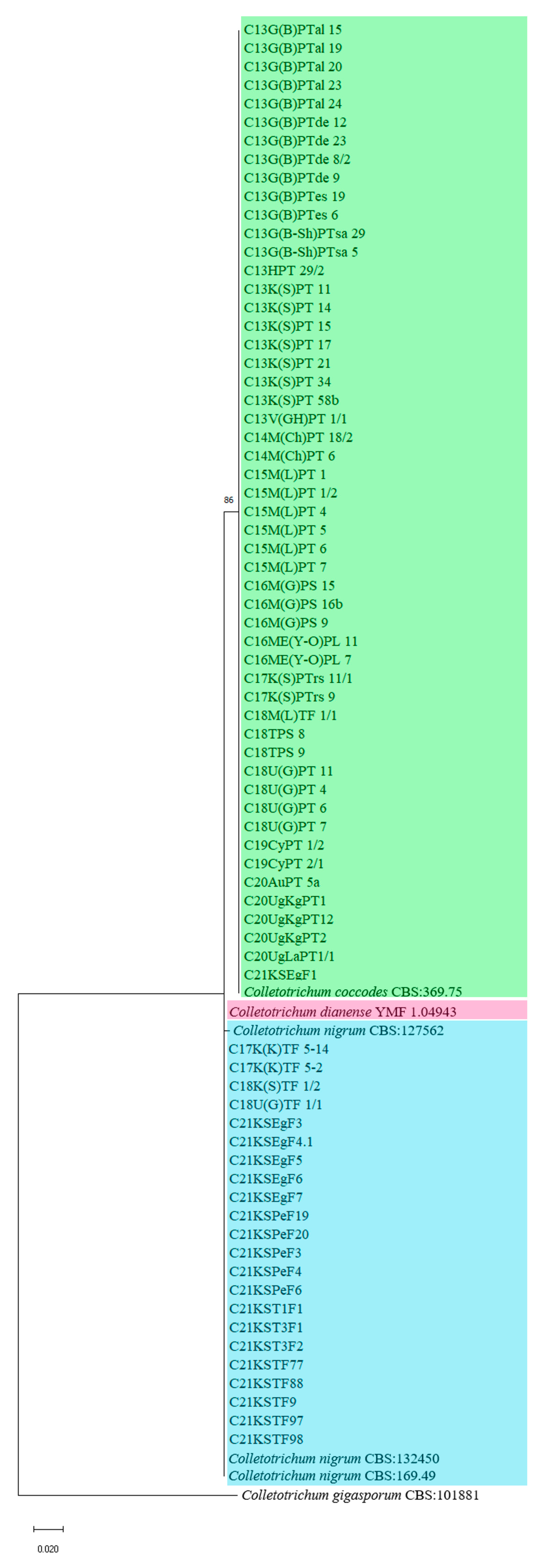
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayawardena, R.S.; Bhunjun, C.S.; Hyde, K.D.; Gentekaki, E.; Itthayakorn, P. Colletotrichum: Lifestyles, biology, morpho-species, species complexes and accepted species. Mycosphere 2021, 12, 519–669. [Google Scholar] [CrossRef]
- Talhinhas, P.; Baroncelli, R. Colletotrichum species and complexes: Geographic distribution, host range and conservation status. Fungal Divers. 2021, 110, 109–198. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating species boundaries in Colletotrichum. Fungal Divers. 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Liu, F.; Ma, Z.Y.; Hou, L.W.; Diao, Y.Z.; Wu, W.P.; Damm, U.; Cai, L. Updating species diversity of Colletotrichum, with a phylogenomic overview. Stud. Mycol. 2022, 101, 1–86. [Google Scholar] [CrossRef]
- Banya, M.; Garg, S.; Meena, N.L. A review: Chilli anthracnose, its spread and management. J. Pharmacogn. Phytochem. 2020, 9, 1432–1438. [Google Scholar]
- Fiers, M.; Edel-Hermann, V.; Chatot, C.; Le Hingrat, Y.; Alabouvette, C.; Steinberg, C. Potato soil-borne diseases. A review. Agron. Sustain. Dev. 2012, 32, 93–132. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.A.; Geary, B.; (Lahkim) Tsror, L. Potato Black Dot—The Elusive Pathogen, Disease Development and Management. Am. J. Potato Res. 2018, 95, 340–350. [Google Scholar] [CrossRef]
- Belov, G.L.; Belosokhov, A.F.; Kutuzova, I.A.; Statsyuk, N.V.; Chudinova, E.M.; Alexandrova, A.V.; Kokaeva, L.Y. Colletotrichum coccodes in potato and tomato leaves in Russia. J. Plant Dis. Prot. 2018, 125, 311–317. [Google Scholar] [CrossRef]
- Garibaldi, A.; Gilardi, G.; Baudino, M.; Ortu, G.; Gullino, M.L. Colletotrichum coccodes: A soil-borne pathogen dangerous to a pepper rootstock in Italy. J. Plant Pathol. 2012, 94, 91. [Google Scholar]
- Ali, A.; Bordoh, P.K.; Singh, A.; Siddiqui, Y.; Droby, S. Post-harvest development of anthracnose in pepper (Capsicum spp): Etiology and management strategies. Crop Prot. 2016, 90, 132–141. [Google Scholar] [CrossRef]
- Tyvaert, L.; Everaert, E.; Lippens, L.; Cuijpers, W.J.M.; França, S.C.; Höfte, M. Interaction of Colletotrichum coccodes and Verticillium dahliae in pepper plants. European J. Plant Pathol. 2019, 155, 1303–1317. [Google Scholar] [CrossRef]
- Boyette, C.D.; Hoagland, R.E.; Stetina, K.C. Bioherbicidal enhancement and host range expansion of a mycoherbicidal fungus via formulation approaches. Biocontrol Sci. Technol. 2018, 28, 307–315. [Google Scholar] [CrossRef]
- Manova, V.; Stoyanova, Z.; Rodeva, R.; Boycheva, I.; Korpelainen, H.; Vesterinen, E.; Wirta, H.; Bonchev, G. Morphological, Pathological and Genetic Diversity of the Colletotrichum Species, Pathogenic on Solanaceous Vegetable Crops in Bulgaria. J. Fungi 2022, 8, 1123. [Google Scholar] [CrossRef] [PubMed]
- Shivas, R.G.; Tan, Y.P.; Edwards, J.; Dinh, Q.; Maxwell, A.; Andjic, V.; Liberato, J.R.; Anderson, C.; Beasley, D.R.; Bransgrove, K.; et al. Colletotrichum species in Australia. Australas. Plant Pathol. 2016, 45, 447–464. [Google Scholar] [CrossRef]
- Liu, F.; Cai, L.; Crous, P.W.; Damm, U. Circumscription of the anthracnose pathogens Colletotrichum lindemuthianum and C. nigrum. Mycologia 2013, 105, 844–860. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Salamanca, L.M.; Enzenbacher, T.B.; Derie, M.L.; du Toit, L.J.; Feng, C.; Correll, J.C.; Hausbeck, M.K. First report of Colletotrichum coccodes causing leaf and neck anthracnose on onions (Allium cepa) in Michigan and the United States. Plant Dis. 2012, 96, 769. [Google Scholar] [CrossRef]
- Halsted, B.D. A new anthracnose of peppers. Bull. Torrey Bot. Club 1891, 18, 14–15. [Google Scholar] [CrossRef]
- Rivera, Y.; Stommel, J.; Dumm, J.; Ismaiel, A.; Wyenandt, C.A.; Crouch, J.A. First report of Colletotrichum nigrum causing anthracnose disease on tomato fruit in New Jersey. Plant Dis. 2016, 100, 2162. [Google Scholar] [CrossRef]
- Fungal Databases. Available online: http://nt.ars-grin.gov/fungaldatabases (accessed on 10 June 2022).
- Liu, F.; Hyde, K.D.; Cai, L. Neotypification of Colletotrichum coccodes, the causal agent of potato black dot disease and tomato anthracnose. Mycologia 2011, 2, 248–254. [Google Scholar]
- Kotova, V.V.; Kungurtseva, O.V. Anthracnose of agricultural plants. Vestn. Zashchity Rastenii 2014, 11, 3–132. [Google Scholar]
- Nikitin, M.; Deych, K.; Grevtseva, I.; Girsova, N.; Kuznetsova, M.; Pridannikov, M.; Golikov, A. Preserved microarrays for simultaneous detection and identification of six fungal potato pathogens with the use of real-time PCR in matrix format. Biosensors 2018, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokaeva, L.; Chudinova, E.; Berezov, A.; Yarmeeva, M.; Balabko, P.; Belosokhov, A.; Elansky, S. Fungal diversity in tomato (Solanum lycopersicum) leaves and fruits in Russia. J. Central Eur. Agric. 2020, 21, 809–816. [Google Scholar] [CrossRef]
- Kazartsev, I.A.; Gomzhinaa, M.M.; Gasicha, E.L.; Khlopunova, L.B.; Gannibal, P.B. Biodiversity of Colletotrichum spp. on Several Wild and Cultivated Plants. Mikol. Fitopatol. 2021, 56, 127–139. [Google Scholar]
- Poluektova, E.V.; Berestetskiy, A.O.; Kutuzova, I.A.; Kokaeva, L.Y.; Yarmeeva, M.M.; Khusnetdinova, T.I.; Balabko, P.N.; Kurchaev, M.L.; Elansky, S.N. Biological properties and susceptibility to fungicides of Colletotrichum coccodes strains of different geographic origin. Probl. Agrochem. Ecol. 2021, 3–4, 45–54. [Google Scholar]
- Belosokhov, A.F.; Yarmeeva, M.M.; Kokaeva, L.Y.; Chudinova, E.M.; Mislavsky, S.M.; Elansky, S.N. Trichocladium solani sp. nov.—A new pathogen on potato tubers causing yellow rot. J. Fungi 2022, 8, 1160. [Google Scholar] [CrossRef]
- Elansky, S.N.; Chudinova, E.M.; Elansky, A.S.; Kah, M.O.; Sandzhieva, D.A.; Mukabenova, B.A.; Dedov, A.G. Microorganisms in spent water-miscible metalworking fluids as a resource of strains for their disposal. J. Cleaner Prod. 2022, 350, 1–9. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Stephenson, S.A.; Green, J.R.; Manners, J.M.; Maclean, D.J. Cloning and characterisation of glutamine synthetase from Colletotrichum gloeosporioides and demonstration of elevated expression during pathogenesis on Stylosanthes guianensis. Curr. Genet. 1997, 31, 447–454. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H. Untersuchungen über die Morphologische und Biologische Differenzierung in der Fusarium-Sektion Liseola; Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft, Berlin-Dahlem; Kommissionsverlag Paul Parey: Berlin, Germany, 1976; pp. 1–93. [Google Scholar]
- Horsfall, J.G. Chapter II—Fungicidal Action and Its Measurement. Principles of Fungicidal Action; Chronica Botanica: Waltham, MA, USA, 1956; pp. 15–29. [Google Scholar]
- Yarmeeva, M.M.; Kokaeva, L.Y.; Chudinova, E.M.; Kah, M.O.; Kurchaev, M.L.; Zeyruk, V.N.; Elansky, S.N. Anastomosis groups and sensitivity to fungicides of Rhizoctonia solani strains isolated from potato in Russia. J. Plant Dis. Prot. 2021, 128, 1253–1261. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, Z.; Jiang, X.; Fang, L.; Qiao, M. Endophytic Colletotrichum species from aquatic plants in southwest China. J. Fungi 2022, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Çakır, E.; Karahan, A.; Kurbetli, İ. Involvement of Colletotrichum coccodes causing atypical symptoms of potato tubers in Turkey. J. Plant Dis. Prot. 2019, 126, 173–176. [Google Scholar] [CrossRef]
- Dos Santos Vieira, W.A.; Bezerra, P.A.; da Silva, A.C.; Veloso, J.S.; Câmara, M.P.S.; Doyle, V.P. Optimal markers for the identification of Colletotrichum species. Mol. Phylogenetics Evol. 2020, 143, 1–19. [Google Scholar]
- Aqeel, A.M.; Pasche, J.S.; Gudmestad, N.C. Variability in morphology and aggressiveness among North American vegetative compatibility groups of Colletotrichum coccodes. Phytopathology 2008, 98, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Tozze Júnior, H.J.; Firmino, A.C.; Fischer, I.H.; Furtado, E.L.; Massola Júnior, N.S. Characterization of Colletotrichum spp. isolates associated with fruit trees in the state of São Paulo. Summa Phytopathol. 2015, 41, 270–280. [Google Scholar] [CrossRef]
- Cullen, D.W.; Lees, A.K.; Toth, I.K.; Duncan, J.M. Detection of Colletotrichum coccodes from soil and potato tubers by conventional and quantitative real-time PCR. Plant Pathol. 2002, 51, 281–292. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Hadden, J.F.; Black, L.L. Anthracnose of Pepper Caused by Colletotrichum spp.; AVRDC: Tainan, Taiwan, 1989; pp. 189–199. [Google Scholar]
- Ciofini, A.; Negrini, F.; Baroncelli, R.; Baraldi, E. Management of Post-Harvest Anthracnose: Current Approaches and Future Perspectives. Plants 2022, 11, 1856. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreenivasaprasad, S.; Sharada, K.; Brown, A.E.; Mills, P.R. PCR-based detection of Colletotrichum acutatum on strawberry. Plant Pathol. 1996, 45, 650–655. [Google Scholar] [CrossRef]
- Aqeel, A.M. Response of Colletotrichum coccodes to QoI fungicide in vitro. Plant Pathol. J. 2007, 4, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Chapin, L.J.G.; Wang, Y.; Lutton, E.; Gardener, B.B.M. Distribution and fungicide sensitivity of fungal pathogens causing anthracnose-like lesions on tomatoes grown in Ohio. Plant Dis. 2006, 90, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitzan, N.; Cummings, T.F.; Johnson, D.A. Effect of seed-tuber generation, soilborne inoculum, and azoxystrobin application on development of potato black dot caused by Colletotrichum coccodes. Plant Dis. 2005, 89, 1181–1185. [Google Scholar] [CrossRef] [Green Version]
- Torres-Calzada, C.; Tapia-Tussell, R.; Higuera-Ciapara, I.; Martin-Mex, R.; Nexticapan-Garcez, A.; Perez-Brito, D. Sensitivity of Colletotrichum truncatum to four fungicides and characterization of thiabendazole-resistant isolates. Plant Dis. 2015, 99, 1590–1595. [Google Scholar] [CrossRef] [Green Version]
- Sanders, G.M.; Korsten, L. Survey of fungicide resistance of Colletotrichum gloeosporioides from different avocado production areas. Afr. Avocado Grow. Assoc. 1999, 22, 35–38. [Google Scholar]
- Tsindeliani, A.A.; Skokov, D.A.; Yarmeeva, M.M.; Elansky, S.N.; Chudinova, E.M. Thiabendazole sensitiveness of strains isolated from Solanaceae plants. In Current Mycology in Russia. Materials of the Fifth Mycological Congress in Russia; Academy of Mycology: Moscow, Russia, 2022; pp. 258–259. (In Russian) [Google Scholar]
- Leite, I.C.; Silva, R.A.; Santos, J.E.; Freitas-Lopes, R.L.; Câmara, M.P.; Michereff, S.J.; Lopes, U.P. Analysis of Colletotrichum musae populations from Brazil reveals the presence of isolates with high competitive ability and reduced sensitivity to postharvest fungicides. Plant Pathol. 2020, 69, 1529–1539. [Google Scholar] [CrossRef]
- Hu, M.J.; Grabke, A.; Dowling, M.E.; Holstein, H.J.; Schnabel, G. Resistance in Colletotrichum siamense from peach and blueberry to thiophanate-methyl and azoxystrobin. Plant Dis. 2015, 99, 806–814. [Google Scholar] [CrossRef] [Green Version]
- Chudinova, E.M.; Kokaeva, L.Y.; Elansky, S.N.; Kutuzova, I.A.; Pertsev, A.S.; Pobendinskaya, M.A. The occurrence of thiabendazole-resistant isolates of Helminthosporium solani on potato seed tubers in Russia. J. Plant Dis. Prot. 2020, 127, 421–423. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
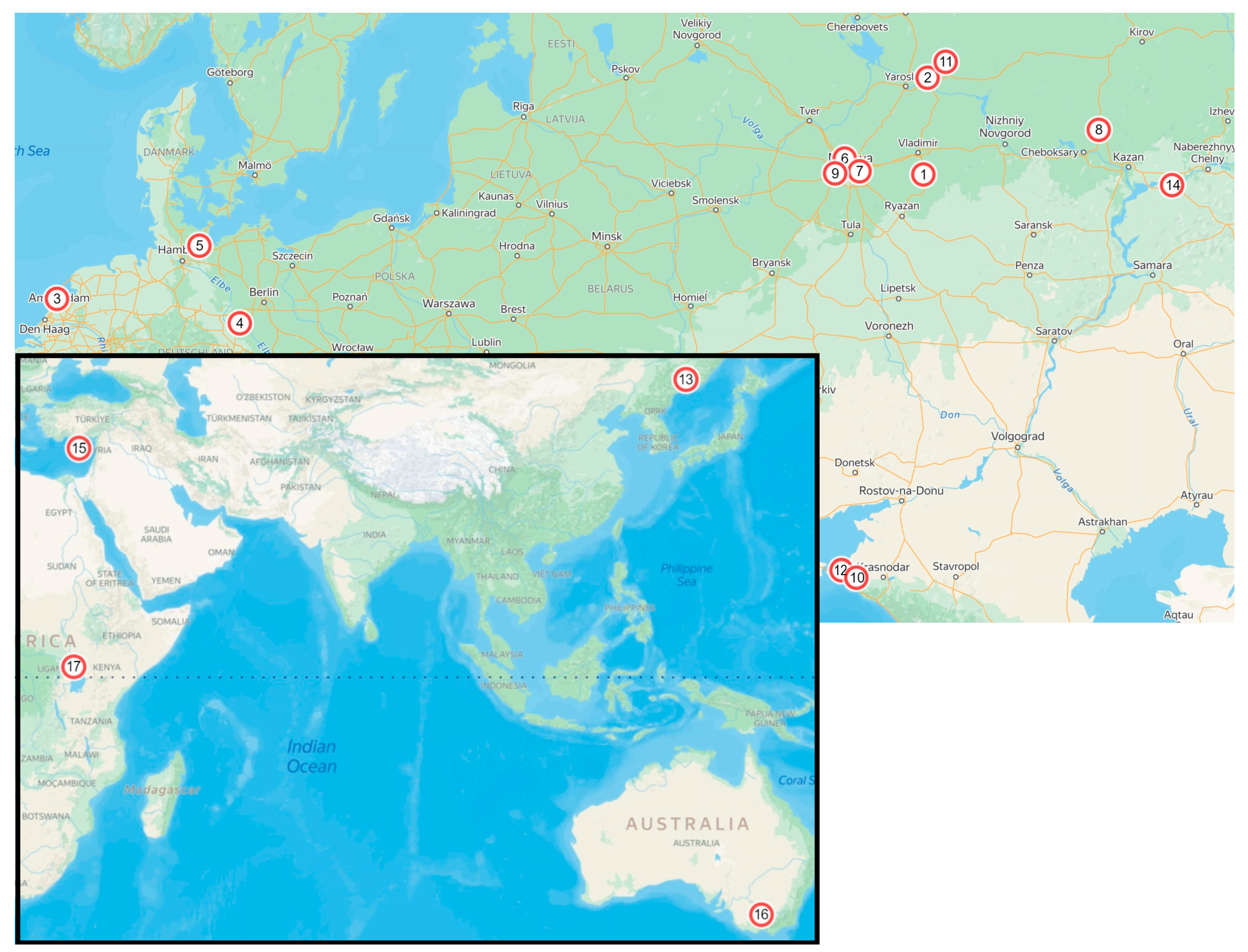
| Strain Identifier | Origin * (Figure 1) | Host Plant | Isolation Source | Year of Isolation | GenBank Accession Numbers ** | |||
|---|---|---|---|---|---|---|---|---|
| ITS | gaphd | act | gs | |||||
| C13V(GH)PT1/1 | 1 | Potato | Tuber | 2013 | OP718477 | OP743730 | OP743793 | OP743860 |
| C13K(S)PT11 | 2 | Potato | Tuber | 2013 | OP718470 | OP743723 | OP743786 | OP743853 |
| C13K(S)PT14 | 2 | Potato | Tuber | 2013 | OP718471 | OP743724 | OP743787 | OP743854 |
| C13K(S)PT15 | 2 | Potato | Tuber | 2013 | OP718472 | OP743725 | OP743788 | OP743855 |
| C13K(S)PT17 | 2 | Potato | Tuber | 2013 | OP718473 | OP743726 | OP743789 | OP743856 |
| C13K(S)PT21 | 2 | Potato | Tuber | 2013 | OP718474 | OP743727 | OP743790 | OP743857 |
| C13K(S)PT34 | 2 | Potato | Tuber | 2013 | OP718475 | OP743728 | OP743791 | OP743858 |
| C13K(S)PT58b | 2 | Potato | Tuber | 2013 | OP718476 | OP743729 | OP743792 | OP743859 |
| C13HPT29/2 | 3 | Potato | Tuber | 2013 | OP718469 | OP743722 | OP743836 | OP743890 |
| C13G(B)PTde8/2 | 4 | Potato | Tuber | 2013 | OP718463 | OP743716 | OP743830 | OP743844 |
| C13G(B)PTde9 | 4 | Potato | Tuber | 2013 | OP718464 | OP743717 | OP743831 | OP743845 |
| C13G(B)PTde12 | 4 | Potato | Tuber | 2013 | OP718461 | OP743714 | OP743828 | OP743842 |
| C13G(B)PTde23 | 4 | Potato | Tuber | 2013 | OP718462 | OP743715 | OP743829 | OP743843 |
| C13G(B)PTes6 | 4 | Potato | Tuber | 2013 | OP718466 | OP743719 | OP743833 | OP743847 |
| C13G(B)PTes19 | 4 | Potato | Tuber | 2013 | OP718465 | OP743718 | OP743832 | OP743846 |
| C13G(B)PTal15 | 4 | Potato | Tuber | 2013 | OP718456 | OP743709 | OP743823 | OP743837 |
| C13G(B)PTal19 | 4 | Potato | Tuber | 2013 | OP718457 | OP743710 | OP743824 | OP743838 |
| C13G(B)PTal20 | 4 | Potato | Tuber | 2013 | OP718458 | OP743711 | OP743825 | OP743839 |
| C13G(B)PTal23 | 4 | Potato | Tuber | 2013 | OP718459 | OP743712 | OP743826 | OP743840 |
| C13G(B)PTal24 | 4 | Potato | Tuber | 2013 | OP718460 | OP743713 | OP743827 | OP743841 |
| C13G(B-Sh)PTsa5 | 5 | Potato | Tuber | 2013 | OP718468 | OP743721 | OP743835 | OP743849 |
| C13G(B-Sh)PTsa29 | 5 | Potato | Tuber | 2013 | OP718467 | OP743720 | OP743834 | OP743848 |
| C14M(Ch)PT6 | 6 | Potato | Tuber | 2014 | OP718479 | OP743732 | OP743795 | OP743862 |
| C14M(Ch)PT18/2 | 6 | Potato | Tuber | 2014 | OP718478 | OP743731 | OP743794 | OP743861 |
| C15M(L)PT1 | 7 | Potato | Tuber | 2015 | OP718480 | OP743733 | OP743796 | OP743863 |
| C15M(L)PT1/2 | 7 | Potato | Tuber | 2015 | OP718481 | OP743734 | OP743797 | OP743864 |
| C15M(L)PT4 | 7 | Potato | Tuber | 2015 | OP718482 | OP743735 | OP743798 | OP743865 |
| C15M(L)PT5 | 7 | Potato | Tuber | 2015 | OP718483 | OP743736 | OP743799 | OP743866 |
| C15M(L)PT6 | 7 | Potato | Tuber | 2015 | OP718484 | OP743737 | OP743800 | OP743867 |
| C15M(L)PT7 | 7 | Potato | Tuber | 2015 | OP718485 | OP743738 | OP743801 | OP743868 |
| C16ME(Y-O)PL7 | 8 | Potato | Leaf | 2016 | OP718490 | OP743743 | OP743806 | OP743873 |
| C16ME(Y-O)PL11 | 8 | Potato | Leaf | 2016 | OP718489 | OP743742 | OP743802 | OP743872 |
| C16M(G)PS9 | 9 | Potato | Stem | 2016 | OP718488 | OP743741 | OP743805 | OP743871 |
| C16M(G)PS15 | 9 | Potato | Stem | 2016 | OP718486 | OP743739 | OP743803 | OP743869 |
| C16M(G)PS16b | 9 | Potato | Stem | 2016 | OP718487 | OP743740 | OP743804 | OP743870 |
| C17K(K)TF5-2 | 10 | Tomato | Fruit | 2017 | OP718492 | OP743745 | OP743808 | OP743875 |
| C17K(K)TF5-14 | 10 | Tomato | Fruit | 2017 | OP718491 | OP743744 | OP743807 | OP743874 |
| C17K(S)PTrs9 | 11 | Potato | Tuber | 2017 | OP718494 | OP743747 | OP743810 | OP743877 |
| C17K(S)PTrs11/1 | 11 | Potato | Tuber | 2017 | OP718493 | OP743746 | OP743809 | OP743876 |
| C18M(L)TF1/1 | 7 | Tomato | Fruit | 2018 | OP718496 | OP743749 | OP743822 | OP743889 |
| C18K(S)TF1/2 | 12 | Tomato | Fruit | 2018 | OP718495 | OP743748 | OP743811 | OP743878 |
| C18U(G)TF1/1 | 13 | Tomato | Fruit | 2018 | OP716941 | OP730520 | OP743774 | OP743898 |
| C18U(G)PT4 | 13 | Potato | Tuber | 2018 | OP718500 | OP743753 | OP743815 | OP743882 |
| C18U(G)PT6 | 13 | Potato | Tuber | 2018 | OP718501 | OP743754 | OP743816 | OP743883 |
| C18U(G)PT7 | 13 | Potato | Tuber | 2018 | OP718502 | OP743755 | OP743817 | OP743884 |
| C18U(G)PT11 | 13 | Potato | Tuber | 2018 | OP718499 | OP743752 | OP743814 | OP743881 |
| C18TPS8 | 14 | Potato | Stem | 2018 | OP718497 | OP743750 | OP743812 | OP743879 |
| C18TPS9 | 14 | Potato | Stem | 2018 | OP718498 | OP743751 | OP743813 | OP743880 |
| C19CyPT1/2 | 15 | Potato | Tuber | 2019 | OP718503 | OP743756 | OP743783 | OP743850 |
| C19CyPT2/1 | 15 | Potato | Tuber | 2019 | OP718504 | OP743757 | OP743784 | OP743851 |
| C20AuPT5a | 16 | Potato | Tuber | 2020 | OP718505 | OP743758 | OP743785 | OP743852 |
| C20UgLaPT1/1 | 17 | Potato | Tuber | 2020 | OL405711 | OP743762 | OP743821 | OP743888 |
| C20UgKgPT1 | 17 | Potato | Tuber | 2020 | OP718506 | OP743759 | OP743818 | OP743885 |
| C20UgKgPT2 | 17 | Potato | Tuber | 2020 | OP718508 | OP743761 | OP743820 | OP743887 |
| C20UgKgPT12 | 17 | Potato | Tuber | 2020 | OP718507 | OP743760 | OP743819 | OP743886 |
| C21KST1F1 | 12 | Tomato | Fruit | 2021 | OP716934 | OP730512 | OP743775 | OP743891 |
| C21KSTF9 | 12 | Tomato | Fruit | 2021 | OP716939 | OP730517 | OP743780 | OP743896 |
| C21KST3F1 | 12 | Tomato | Fruit | 2021 | OP716935 | OP730513 | OP743776 | OP743892 |
| C21KST3F2 | 12 | Tomato | Fruit | 2021 | OP716936 | OP730514 | OP743777 | OP743893 |
| C21KSTF88 | 12 | Tomato | Fruit | 2021 | OP716938 | OP730516 | OP743779 | OP743895 |
| C21KSTF77 | 12 | Tomato | Fruit | 2021 | OP716937 | OP730515 | OP743778 | OP743894 |
| C21KSTF97 | 12 | Tomato | Fruit | 2021 | OP716940 | OP730518 | OP743781 | OP743897 |
| C21KSTF98 | 12 | Tomato | Fruit | 2021 | OP716941 | OP730519 | OP743782 | OP743899 |
| C21KSPeF3 | 12 | Pepper | Fruit | 2021 | OP716931 | OP743706 | OP743771 | OP743908 |
| C21KSPeF4 | 12 | Pepper | Fruit | 2021 | OP716932 | OP743707 | OP743772 | OP743909 |
| C21KSPeF6 | 12 | Pepper | Fruit | 2021 | OP716933 | OP743708 | OP743773 | OP743910 |
| C21KSPeF20 | 12 | Pepper | Fruit | 2021 | OP716930 | OP743705 | OP743770 | OP743907 |
| C21KSPeF19 | 12 | Pepper | Fruit | 2021 | OP716929 | OP743704 | OP743769 | OP743906 |
| C21KSEgF1 | 12 | Eggplant | Fruit | 2021 | OP716923 | OP743698 | OP743763 | OP743900 |
| C21KSEgF3 | 12 | Eggplant | Fruit | 2021 | OP716924 | OP743699 | OP743764 | OP743901 |
| C21KSEgF4.1 | 12 | Eggplant | Fruit | 2021 | OP716925 | OP743700 | OP743765 | OP743902 |
| C21KSEgF5 | 12 | Eggplant | Fruit | 2021 | OP716926 | OP743701 | OP743766 | OP743903 |
| C21KSEgF6 | 12 | Eggplant | Fruit | 2021 | OP716927 | OP743702 | OP743767 | OP743904 |
| C21KSEgF7 | 12 | Eggplant | Fruit | 2021 | OP716928 | OP743703 | OP743768 | OP743905 |
| Species | Species Complex | Strain Identifier | Host | GenBank Accession Numbers * | |||
|---|---|---|---|---|---|---|---|
| ITS | act | gaphd | gs | ||||
| C. nigrum | singleton | CBS 69.49 | Capsicum sp. | NR163523 | JX546646 | JX546742 | - |
| C. nigrum | singleton | CBS 132450 | Solanum lycopersicum | JX546845 | JX546653 | JX546749 | - |
| C. nigrum | singleton | CBS 127562 | Cichorium intybus | JX546842 | JX546650 | JX546746 | - |
| C. dianense | singleton | YMF 1.04943 | Alternanthera philoxeroides | OL842189 | OL981258 | OL981284 | - |
| C. coccodes | singleton | CBS 369.75 | Solanum tuberosum | HM171679 | HM171667 | HM171673 | HM171676 |
| C. gigasporum | Gigasporum | CBS 101881 | Cyphomandra betacea | KF687736 | KF687797 | KF687841 | KF687745 |
| Isolate | Percentage of Similarity to Type Strains | |||
|---|---|---|---|---|
| ITS Sequence | act Sequence | gaphd Sequence | gs Sequence ** | |
| C18U(G)TF1/1 | 100% C. coccodes 100% C. dianense 100% C. nigrum | 100% C. nigrum 100% C. dianense 97% C. coccodes | 100% C. nigrum 100% C. dianense 97% C. coccodes | 97% C. coccodes |
| C21KST1F1 | 100% C. coccodes 100% C. dianense 100% C. nigrum | 100% C. nigrum 100% C. dianense 97% C. coccodes | 100% C. nigrum 100% C. dianense 97% C. coccodes | 97% C. coccodes |
| C21KST3F1 | 100% C. coccodes 100% C. dianense 100% C. nigrum | 100% C. nigrum 100% C. dianense 97% C. coccodes | 100% C. nigrum 100% C. dianense 97% C. coccodes | 97% C. coccodes |
| C21KSTF77 | 100% C. coccodes 100% C. dianense 100% C. nigrum | 100% C. nigrum 100% C. dianense 97% C. coccodes | 100% C. nigrum 100% C. dianense 97% C. coccodes | 97% C. coccodes |
| C21KSPeF20 | 100% C. coccodes 100% C. dianense 100% C. nigrum | 100% C. nigrum 100% C. dianense 97% C. coccodes | 100% C. nigrum 100% C. dianense 97% C. coccodes | 97% C. coccodes |
| Strain Identifier | Species | Host | Average Disease Radius (mm) on Tomato after | Average Disease Radius (mm) on Potato after | ||
|---|---|---|---|---|---|---|
| 21 days | 35 days | 21 days | ||||
| Wound Inoculation | Surface Inoculation | Surface Inoculation | Wound Inoculation | |||
| C21KSEgF7 | C. nigrum | Eggplant | 4.8 | 0.0 | 0.9 | 2.3 |
| C21KSEgF3 | C. nigrum | Eggplant | 3.2 | 0.1 | 2.0 | 2.5 |
| C21KSEgF4.1 | C. nigrum | Eggplant | 3.2 | 0.1 | 2.3 | 4.3 |
| C21KSPeF6 | C. nigrum | Pepper | 3.7 | 1.0 | 9.8 | 2.0 |
| C21KSPeF19 | C. nigrum | Pepper | 5.2 | 0.3 | 2.5 | 8.3 |
| C20AuPT5a | C. coccodes | Potato | 6.2 | 0.2 | 2.0 | 5.2 |
| C20UgKgPT2 | C. coccodes | Potato | 5.2 | 1.2 | 2.2 | 2.5 |
| C21KSTF88 | C. nigrum | Tomato | 8.3 | 0.5 | 3.3 | 1.8 |
| C21KSTF97 | C. nigrum | Tomato | 7.5 | 0.5 | 1.2 | 1.2 |
| C21KST3F2 | C. nigrum | Tomato | 6.3 | 0.2 | 3.2 | 4.0 |
| Strain Identifier | Species | Isolation Source * | EC50, mg/L ** | ||
|---|---|---|---|---|---|
| Thiabendazole | Azoxystrobin | Difenoconazole | |||
| C13V(GH)PT1/1 | C. coccodes | PT | 4.24 | 0.08 | 0.06 |
| C13K(S)PT11 | C. coccodes | PT | 5.07 | 7.75 | 0.07 |
| C13K(S)PT14 | C. coccodes | PT | 7.75 | 0.08 | 0.06 |
| C13K(S)PT15 | C. coccodes | PT | 4.47 | 0.28 | 0.07 |
| C13K(S)PT17 | C. coccodes | PT | 10.20 | 5.82 | 0.12 |
| C13K(S)PT21 | C. coccodes | PT | 7.90 | 3.68 | - |
| C13K(S)PT34 | C. coccodes | PT | 0.78 | 0.07 | 0.06 |
| C13K(S)PT58b | C. coccodes | PT | 3.47 | - | 0.07 |
| C13HPT29/2 | C. coccodes | PT | 0.91 | 0.05 | 0.05 |
| C13G(B)PTde9 | C. coccodes | PT | 50.30 | 0.07 | 0.06 |
| C13G(B)PTde12 | C. coccodes | PT | 0.85 | 0.06 | 0.05 |
| C13G(B)PTde23 | C. coccodes | PT | 0.93 | 0.09 | 0.06 |
| C13G(B)PTes6 | C. coccodes | PT | 33.38 | 0.10 | 0.06 |
| C13G(B)PTes19 | C. coccodes | PT | 8.78 | 0.10 | 0.06 |
| C13G(B)PTal15 | C. coccodes | PT | 0.96 | 0.09 | 0.09 |
| C13G(B)PTal19 | C. coccodes | PT | 0.96 | 0.09 | 0.06 |
| C13G(B)PTal20 | C. coccodes | PT | 0.99 | 0.08 | 0.06 |
| C13G(B)PTal23 | C. coccodes | PT | 6.13 | 0.07 | 0.07 |
| C13G(B)PTal24 | C. coccodes | PT | 0.85 | 0.06 | 0.05 |
| C13G(B-Sh)PTsa29 | C. coccodes | PT | 1.00 | 0.08 | 0.06 |
| C14M(Ch)PT6 | C. coccodes | PT | - | 0.09 | 0.06 |
| C14M(Ch)PT18/2 | C. coccodes | PT | - | 0.08 | 0.06 |
| C15M(L)PT1 | C. coccodes | PT | 0.82 | 0.06 | 0.08 |
| C15M(L)PT1/2 | C. coccodes | PT | 0.94 | - | 0.09 |
| C15M(L)PT4 | C. coccodes | PT | 0.89 | 0.08 | 0.09 |
| C15M(L)PT5 | C. coccodes | PT | 0.85 | 0.08 | 0.09 |
| C15M(L)PT6 | C. coccodes | PT | - | 0.08 | 0.06 |
| C15M(L)PT7 | C. coccodes | PT | 0.95 | - | 0.09 |
| C16ME(Y-O)PL7 | C. coccodes | PL | - | - | 0.09 |
| C16ME(Y-O)PL11 | C. coccodes | PL | - | - | 0.08 |
| C16M(G)PS9 | C. coccodes | PS | 0.85 | 0.08 | 0.09 |
| C16M(G)PS15 | C. coccodes | PS | 0.84 | 4.09 | 0.06 |
| C16M(G)PS16b | C. coccodes | PS | 58.38 | 0.07 | 0.08 |
| C17K(K)TF5-2 | C. nigrum | TF | 0.91 | 0.09 | 0.09 |
| C17K(K)TF5-14 | C. nigrum | TF | - | 0.08 | 0.09 |
| C17K(S)PTrs9 | C. coccodes | PT | - | 0.08 | 0.06 |
| C17K(S)PTrs11/1 | C. coccodes | PT | 0.87 | 0.08 | 0.07 |
| C18M(L)TF1/1 | C. coccodes | TF | 0.74 | 6.32 | 0.12 |
| C18K(S)TF1/2 | C. nigrum | TF | 20.29 | 8.50 | - |
| C18U(G)TF1/1 | C. nigrum | TF | - | - | 0.07 |
| C18U(G)PT4 | C. coccodes | PT | 6.07 | - | - |
| C18U(G)PT7 | C. coccodes | PT | 0.65 | 7.75 | 0.10 |
| C18U(G)PT11 | C. coccodes | PT | 25.43 | 9.07 | 0.08 |
| C18TPS8 | C. coccodes | PS | - | 7.75 | 0.09 |
| C18TPS9 | C. coccodes | PS | - | 3.31 | 0.09 |
| C19CyPT1/2 | C. coccodes | PT | 0.95 | 0.07 | 0.09 |
| C19CyPT2/1 | C. coccodes | PT | 0.85 | 0.07 | 0.09 |
| C20AuPT5a | C. coccodes | PT | 0.75 | 0.08 | 0.07 |
| C20UgLaPT1/1 | C. coccodes | PT | 0.71 | 0.07 | 0.07 |
| C20UgKgPT1 | C. coccodes | PT | 0.82 | 0.08 | 0.07 |
| C20UgKgPT2 | C. coccodes | PT | 0.73 | 0.08 | 0.07 |
| C20UgKgPT12 | C. coccodes | PT | 0.83 | 0.07 | 0.07 |
| C21KST3F2 | C. nigrum | TF | 0.66 | 0.08 | 0.07 |
| C21KSTF88 | C. nigrum | TF | 0.67 | 0.08 | 0.07 |
| C21KSTF97 | C. nigrum | TF | 0.65 | 0.08 | 0.07 |
| C21KSPeF6 | C. nigrum | PeF | 0.68 | 0.08 | 0.07 |
| C21KSPeF19 | C. nigrum | PeF | 0.68 | 0.08 | 0.07 |
| C21KSEgF3 | C. nigrum | EF | 0.58 | 0.08 | 0.07 |
| C21KSEgF4.1 | C. nigrum | EF | 0.64 | 0.08 | 0.07 |
| C21KSEgF6 | C. nigrum | EF | 0.71 | 0.08 | 0.07 |
| C21KSEgF7 | C. nigrum | EF | 0.66 | 0.09 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yarmeeva, M.; Kutuzova, I.; Kurchaev, M.; Chudinova, E.; Kokaeva, L.; Belosokhov, A.; Belov, G.; Elansky, A.; Pobedinskaya, M.; Tsindeliani, A.; et al. Colletotrichum Species on Cultivated Solanaceae Crops in Russia. Agriculture 2023, 13, 511. https://doi.org/10.3390/agriculture13030511
Yarmeeva M, Kutuzova I, Kurchaev M, Chudinova E, Kokaeva L, Belosokhov A, Belov G, Elansky A, Pobedinskaya M, Tsindeliani A, et al. Colletotrichum Species on Cultivated Solanaceae Crops in Russia. Agriculture. 2023; 13(3):511. https://doi.org/10.3390/agriculture13030511
Chicago/Turabian StyleYarmeeva, Maria, Irina Kutuzova, Michael Kurchaev, Elena Chudinova, Ludmila Kokaeva, Arseniy Belosokhov, Grigory Belov, Alexander Elansky, Marina Pobedinskaya, Archil Tsindeliani, and et al. 2023. "Colletotrichum Species on Cultivated Solanaceae Crops in Russia" Agriculture 13, no. 3: 511. https://doi.org/10.3390/agriculture13030511