
Effect of Irrigation, Nitrogen Fertilization and Amino Acid Biostimulant on Proximate Composition and Energy Value of Pisum sativum L. Seeds

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
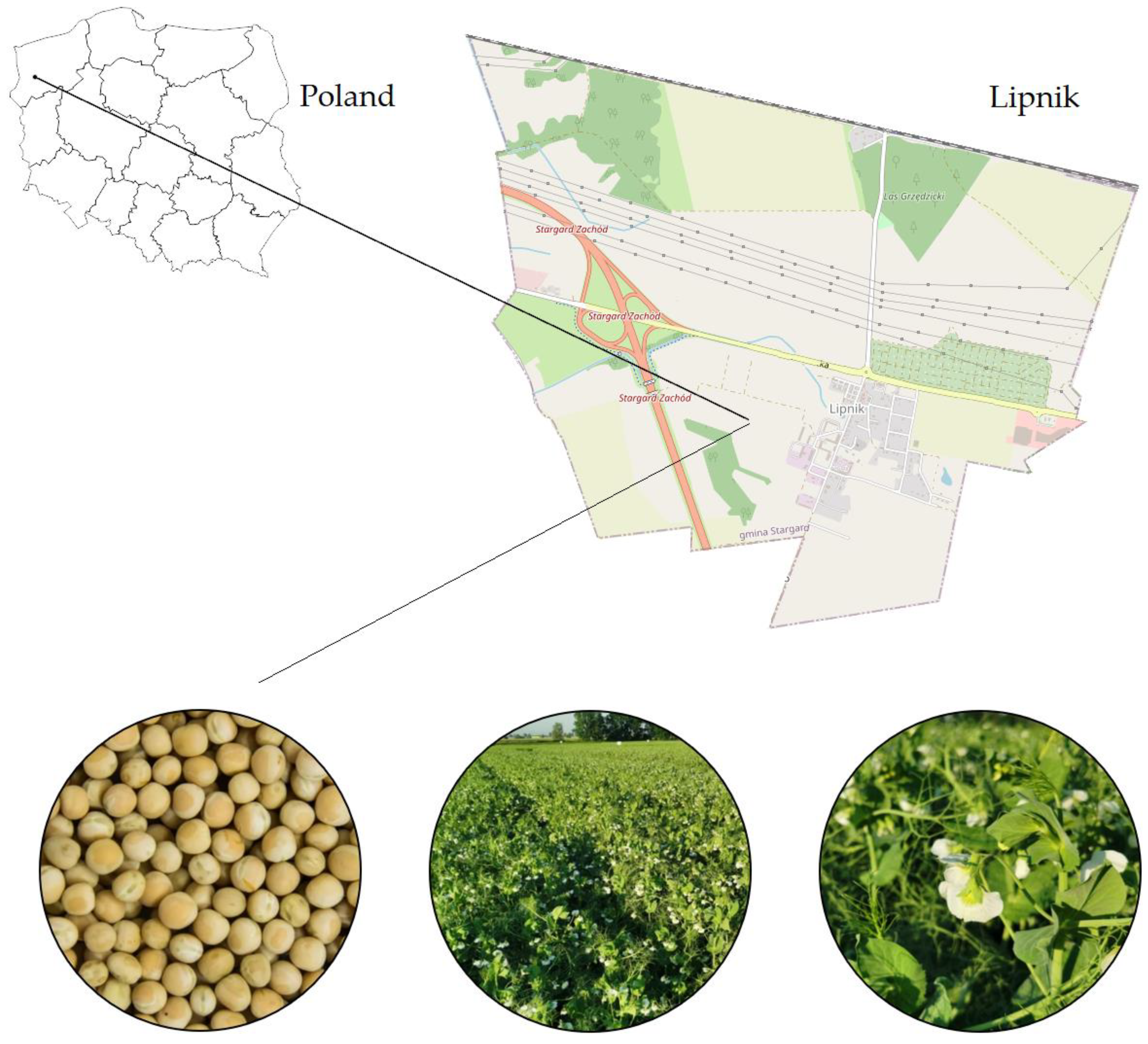
2.1. Field Experiment
2.2. Chemical Analyses
2.3. Energy Value
2.4. Statistical Analyses
- —value of the examined feature for the i-th level of factor A, l-th level of factor B, p-th of factor C in the j-th repetition,
- m—mean of the experimental setup,
- ai, bl, cp—the effects of the studied factors, respectively,
- gj—replication effect,
- abil, acip, bclp, abcilp—appropriate effects of factor interaction,
- eij, eijl, eijlp—random effects.
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zmudzińska, A.; Bigorowski, B.; Banaszak, M.; Roślewska, A.; Adamski, M.; Hejdysz, M. The Effect of diet based on legume seeds and rapeseed meal on pig performance and meat quality. Animals 2020, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Chen, F.; Bu, G.; Zhang, L. Distribution and degradation of DNA from non-genetically and genetically modified soybean (Roundup Ready): Impact of soybean protein concentrate and soybean protein isolate preparation. Food Chem. 2021, 335, 127582. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.; Ammann, K. New GMO regulations for old: Determining a new future for EU crop biotechnology. GM Crops Food 2017, 8, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Teferra, T.F. Should we still worry about the safety of GMO foods? Why and why not? A review. Food Sci. Nutr. 2021, 9, 5324–5331. [Google Scholar] [CrossRef]
- Sońta, M.; Rekiel, A.; Więcek, J.; Batorska, M.; Puppel, K. Alternative protein sources vs. GM soybean meal as feedstuff for pigs—Meat quality and health-promoting indicators. Animals 2021, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Palmero, F.; Fernandez, J.A.; Garcia, F.O.; Ricardo, J.; Haro, R.J.; Prasad, P.V.; Salvagiotti, F.; Ignacio, A.; Ciampitti, I.A. A quantitative review into the contributions of biological nitrogen fixation to agricultural systems by grain legumes. Eur. J. Agron. 2022, 136, 126514. [Google Scholar] [CrossRef]
- Zander, P.; Amjath-Babu, T.S.; Preissel, S.; Reckling, M.; Bues, A.; Schläfke, N.; Kuhlman, T.; Bachinger, J.; Uthes, S.; Stoddard, F.; et al. Grain legume decline and potential recovery in European agriculture: A review. Agron. Sustain. Dev. 2016, 36, 26. [Google Scholar] [CrossRef]
- Badora, B. Opinions about Genetically Modified Crops (GMCs); Public Opinion Research Center: Warsaw, Poland, 2013; Available online: https://www.cbos.pl/SPISKOM.POL/2013/K_002_13.PDF (accessed on 12 December 2022).
- Alonso, R.; Aguire, A.; Marzo, F. Effects of extrusion and traditional processing methods on antinutrients and in vitro digestibility of protein and starch in faba and kidney beans. Food Chem. 2000, 68, 159–165. [Google Scholar] [CrossRef]
- Sujak, A.; Kotlarz, A.; Strobel, W. Compositional and nutritional evaluation of several lupin seeds. Food Chem. 2006, 98, 711–719. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Zannou, O.; Karim, I.; Awad, N.M.; Gołaszewski, J.; Heinz, V.; Smetana, S. Avoiding food neophobia and increasing consumer acceptance of new food trends—A decade of research. Sustainability 2022, 14, 10391. [Google Scholar] [CrossRef]
- Ferreira, H.; Pinto, E.; Vasconcelos, M.W. Legumes as a cornerstone of the transition toward more sustainable agri-food systems and diets in Europe. Front. Sustain. Food Syst. 2021, 5, 694121. [Google Scholar] [CrossRef]
- Hara, P.; Piekutowska, M.; Niedbała, G. Prediction of protein content in pea (Pisum sativum L.) seeds using artificial neural networks. Agriculture 2023, 13, 29. [Google Scholar] [CrossRef]
- Hejdysz, M.; Kaczmarek, S.A.; Adamski, M.; Rutkowski, A. Influence of graded inclusion of raw and extruded pea (Pisum sativum L.) meal on the performance and nutrient digestibility of broiler chickens. Anim. Feed. Sci. Technol. 2017, 230, 114–125. [Google Scholar] [CrossRef]
- FAOSTAT; Food and Agriculture Organization of the United Nations; FAOSTAT Statistical Database. Building Climate Resilience for Food Security and Nutrition. In The State of Food Security and Nutrition in the World; FAO: Rome, Italy, 2018. [Google Scholar]
- Monti, M.; Pellicanò, A.; Pristeri, A.; Badagliacca, G.; Preiti, G.; Gelsomino, A. Cereal/grain legume intercropping in rotation with durum wheat in crop/livestock production systems for Mediterranean farming system. Field Crops Res. 2019, 240, 23–33. [Google Scholar] [CrossRef]
- Semba, R.D.; Ramsing, R.; Rahman, N.; Kraemer, K.; Bloem, M.W. Legumes as a sustainable source of protein in human diets. Glob. Food Sec. 2021, 28, 100520. [Google Scholar] [CrossRef]
- Lee, S.A.; Park, C.S.; Kim, B.G. Novel two-slope equations to predict amino acid concentrations using crude protein concentration in soybean meal. Agriculture 2021, 11, 280. [Google Scholar] [CrossRef]
- Walter, S.; Zehring, J.; Mink, K.; Quendt, U.; Zocher, K.; Rohn, S. Protein content of peas (Pisum sativum) and beans (Vicia faba)—Influence of cultivation conditions. J. Food Compost. Anal. 2022, 105, 104257. [Google Scholar] [CrossRef]
- INRAE-CIRAD-AFZ Feed Tables. Available online: https://www.feedtables.com/ (accessed on 20 January 2023).
- Rodriguez, C.; Frias, J.; Vidal-Valverde, C.; Hernández, A. Correlations between some nitrogen fractions, lysine, histidine, tyrosine, and ornithine contents during the germination of peas, beans, and lentils. Food Chem. 2008, 108, 245–252. [Google Scholar] [CrossRef]
- Laudadio, V.; Nahashon, S.N.; Tufarelli, V. Growth performance and carcass characteristics of guinea fowl broilers fed micronized-dehulled pea (Pisum sativum L.) as a substitute for soybean meal. Poult. Sci. 2012, 91, 2988–2996. [Google Scholar] [CrossRef]
- Bandegan, A.; Golian, A.; Kiarie, E.; Payne, R.L.; Crow, G.H.; Guenter, W.; Nyachoti, C.M. Standardized ileal amino acid digestibility in wheat, barley, pea and flaxseed for broiler chickens. Can. J. Anim. Sci. 2011, 1, 91. [Google Scholar] [CrossRef]
- Bandegan, A.; Golian, A.; Kiarie, E.; Payne, R.L.; Crow, G.H.; Guenter, W.; Nyachoti, C.M. Standardized ileal amino acid digestibility in dry-extruded expelled soybean meal, extruded canola seed-pea, feather meal, and poultry by-product meal for broiler chickens. Poultry Sci. 2010, 89, 2626–2633. [Google Scholar] [CrossRef] [PubMed]
- Sirtori, F.; Crovetti, A.; Acciaioli, A.; Bonelli, A.; Pugliese, C.; Bozzi, R.; Campodoni, G.; Franci, O. Effect of replacing a soy diet with Vicia faba and Pisum sativum on performance, meat and fat traits of Cinta Senese pigs. Ital. J. Anim. Sci. 2015, 14, 99–104. [Google Scholar] [CrossRef]
- Zduńczyk, Z.; Mikulski, D.; Jankowski, J.; Slominski, B.A.; Juśkiewicz, J. The effect of the dietary inclusion of pea seeds of colored-flowered and white-flowered varieties on gastrointestinal function in turkeys. Anim. Nutr. 2022, 10, 167–177. [Google Scholar] [CrossRef]
- Kowalska, E.; Kucharska-Gaca, J.; Kuźniacka, J.; Lewko, L.; Gornowicz, E.; Biesek, J.; Adamski, M. Egg quality depending on the diet with different sources of protein and age of the hens. Sci. Rep. 2021, 11, 2638. [Google Scholar] [CrossRef] [PubMed]
- Andrianova, E.; Egorov, I.; Pronin, V. Efficiency and physiological safety of peas in the diets for hens (Gallus gallus L.) of the parent flock during the late laying period. Biol. Agricul. 2020, 55, 1245–1256. [Google Scholar] [CrossRef]
- Sahaf, S.A.; Zarghi, H. Effect of raw or autoclaved grass pea seeds in diet on performance and egg quality of laying hens. Anim. Prod. 2018, 20, 131–143. [Google Scholar] [CrossRef]
- Koivunen, E.; Tuunainen, P.; Valkonen, E.; Valaja, J. Use of semi-leafess peas (Pisum sativum L.) in laying hens diets. Agric. Food Sci. 2015, 24, 84–91. [Google Scholar] [CrossRef]
- Witten, S.; Böhm, H.; Aulrich, K. Effect of variety and environment on the contents of crude nutrients, lysine, methionine and cysteine in organically produced field peas (Pisum sativum L.) and field beans (Vicia faba L.). Landbauforschung-Appli. Agric. For. Res. 2015, 65, 205–216. [Google Scholar] [CrossRef]
- Fallon, E.; Tremblay, N.; Desjardins, Y. Relationships among growing degree-days tenderness, other harvest attributes and market value of processing pea (Pisum sativum L.) cultivars grown in Quebec. Can. J. Plant Sci. 2006, 86, 525–537. [Google Scholar] [CrossRef]
- Boroojeni, F.G.; Männer, K.; Boros, D.; Wiśniewska, M.; Kühnel, S.; Beckmann, K.; Zentek, J. Spontaneous and enzymatic fermentation of rapeseed cake for broiler nutrition. Anim. Feed Sci. Technol. 2022, 284, 115–135. [Google Scholar] [CrossRef]
- Elsheikh, E.A.E.; Mohamed, S.S.; Elhussein, A.A.; Babiker, E.E. Effects of intercropping, Bradyrhizobium inoculation and chicken manure fertilization on the chemical composition and physical characteristics of soybean seed. Food Chem. 2009, 112, 690–694. [Google Scholar] [CrossRef]
- Erman, M.; Ari, E.; Togay, Y.; Çig, F. Response of field pea (Pisum sativum sp. arvense L.) to rhizobium inoculation and nitrogen application in eastern Anotolia. J. Vet. Adv. 2009, 8, 612–616. [Google Scholar]
- Pankou, C.; Lithourgidis, A.; Dordas, C. Effect of irrigation on intercropping systems of wheat (Triticum aestivum L.) with pea (Pisum sativum L.). Agronomy 2021, 11, 283. [Google Scholar] [CrossRef]
- Duzdemir, O.; Kurunc, A.; Unlukara, A. Response of pea (Pisum sativum) to salinity and irrigation water regime. Bulg. J. Agric. Sci. 2009, 15, 400–409. [Google Scholar]
- Chai, Q.; Qin, A.; Gan, Y.; Yu, A. Higher yield and lower carbon emission by intercropping maize with rape, pea, and wheat in arid irrigation areas. Agron. Sustain. Dev. 2014, 34, 535–543. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32019R1009 (accessed on 24 October 2022).
- Radkowski, A.; Radkowska, I. Effect of foliar application of growth biostimulant on quality and nutritive value of meadow sward. Ecol. Chem. Eng. A. 2013, 20, 1205–1211. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Simón-Grao, S.; Navarro-Pérez, V.; Alfosea-Simón, M. Scientific Advances in Biostimulation Reported in the 5th Biostimulant World Congress. Horticulturae 2022, 8, 665. [Google Scholar] [CrossRef]
- Kumar, H.D.; Aloke, P. Role of biostimulant formulations in crop production: An overview. Int. J. Appl. Res. Vet. Med. 2020, 8, 38–46. [Google Scholar]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil. 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Petropoulos, S.A. Biostimulants application: A low input cropping management tool for sustainable farming of vegetables. Biomolecules 2021, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Koleška, I.; Hasanagić, D.; Todorović, V.; Murtić, S.; Klokić, I.; Parađiković, N.; Kukavica, B. Biostimulant prevents yield loss and reduces oxidative damage in tomato plants grown on reduced NPK nutrition. J. Plant Interact. 2017, 12, 209–218. [Google Scholar] [CrossRef]
- Witkowicz, R.; Skrzypek, E.; Gleń-Karolczyk, K.; Krupa, M.; Biel, W.; Chłopicka, J.; Galanty, A. Effects of application of plant growth promoters, biological control agents and microbial soil additives on photosynthetic efficiency, canopy vegetation indices and yield of common buckwheat (Fagopyrum esculentum Moench). Biol. Agric. Hortic. 2021, 37, 234–251. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Giordano, M.; El-Nakhel, C.; Cuciniello, A.; Cenvinzo, V.; Colla, G.; Rouphael, Y. Protein hydrolysate or plant extract-based biostimulants enhanced yield and quality performances of greenhouse perennial wall rocket grown in different seasons. Plants 2019, 8, 208. [Google Scholar] [CrossRef]
- Gruszecki, R.; Stawiarz, A. Biostimulants containing amino acids in vegetable crop production. Acta Sci. Pol. Hortorum Cultus. 2021, 20, 45–57. [Google Scholar] [CrossRef]
- Witkowicz, R.; Biel, W.; Chłopicka, J.; Galanty, A.; Gleń-Karolczyk, K.; Skrzypek, E.; Krupa, M. Biostimulants and microorganisms boost the nutritional composition of buckwheat (Fagopyrum esculentum Moench) sprouts. Agronomy 2019, 9, 469. [Google Scholar] [CrossRef]
- WRB; IUSS Working Group. World Reference Base for Soil Resources 2014, Update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; Food and Agriculture Organization of the United Nations: Rome, Italy; Available online: https://www.fao.org/3/i3794en/I3794en.pdf/ (accessed on 23 October 2022).
- AOAC. Official Methods of Analysis, 21st ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2019. [Google Scholar]
- Janssen, I.W.M.M.A. European table of Energy Values for Poultry Feedstuffs, 3rd ed.; Subcommittee Energy of the Working Group: Spelderholt, The Netherlands, 1989. [Google Scholar]
- TIBCO. Available online: https://www.scribd.com/document/321061529/STATISTICA-Electronic-Manual (accessed on 23 October 2022).
- Musigwa, S.; Morgan, N.; Swick, R.; Cozannet, P.; Wu, S.B. Optimization of dietary energy utilization for poultry—A literature review. Poult. Sci. J. 2021, 77, 5–27. [Google Scholar] [CrossRef]
- Tegeder, M.; Rentsch, D. Uptake and partitioning of amino acids and peptides. Mol. Plant Pathol. 2010, 3, 997–1011. [Google Scholar] [CrossRef]
- Singh, L.; Singh, S.B.; Gupta, D.; Lal, M. Effect of irrigation and nitrogen on growth and yield of field pea varieties. Ann. Plant Soil Res. 2015, 17, 96–99. [Google Scholar]
- Paredes, P.; Pereira, L.S.; Rodrigues, G.C.; Botelho, N.; Torres, M.O. Using the FAO dual crop coefficient approach to model water use and productivity of processing pea (Pisum sativum L.) as influenced by irrigation strategies. Agric. Water Manag. 2017, 189, 5–18. [Google Scholar] [CrossRef]
- Achakzai, A.K.K.; Bangulzai, M.I. Effect of various levels of nitrogen fertilizer on the yield and yield attributes of pea (Pisum sativum L.) cultivars. Pak. J. Bot. 2006, 38, 331. [Google Scholar]
- Achakzai, A.K.K. Effect of various levels of nitrogen fertilizer on some vegetative growth attributes of pea (Pisum sativum L.) cultivars. Pak. J. Bot. 2012, 44, 655–659. [Google Scholar]
- Kishorekumar, R.; Bulle, M.; Wany, A.; Gupta, K.J. An overview of important enzymes involved in nitrogen assimilation of plants. Methods Mol. Biol. 2020, 2057, 1–13. [Google Scholar] [CrossRef]
- Singh, V.P.; Tripathi, S.S.; Dimri, D.C. Effect of irrigation schedule on growth, yield attributes and yield in off season vegetable under low hill valley situation of Uttranchal. Veg. Sci. 2007, 28, 149–151. [Google Scholar]
- Yadav, A.K.; Chauhan, S.K.; Shroti, S.K. Effect of sowing dates and nitrogen levels on yield and economics of vegetable pea-wheatmaize cropping system in central part of Uttar Pradesh. Ann. Plant Soil Res. 2012, 14, 159–162. [Google Scholar]
- Voisin, A.S.; Guéguen, J.; Huyghe, C.; Jeuffroy, M.H.; Magrini, M.B.; Meynard, J.C.; Mougel, C.; Pellerin, S.; Pelzer, E. Legumes for feed, food, biomaterials and bioenergy in Europe: A review. Agron. Sustain. Dev. 2014, 34, 361–380. [Google Scholar] [CrossRef]
- Maini, P. The experience of the first biostimulant, based on amino acids and peptides: A short retrospective review on the laboratory researches and the practical results. Fertil. Agrorum. 2006, 1, 29–43. [Google Scholar]
- Grabowska, A.; Kunicki, E.; Sękara, A.; Kalisz, A.; Wojciechowska, R. The effect of cultivar and biostimulant treatment on the carrot yield and its quality. J. Fruit Ornam. Plant Res. 2012, 77, 37–48. [Google Scholar] [CrossRef]
- Ibrahim, S.M.; Ibrahim, H.A.; Omer, A.M. Comparative study of the effects of some organic extract on sugar beet yield under saline conditions. Aust. J. Basic Appl. Sci. 2012, 6, 664–674. [Google Scholar]
- Kunicki, E.; Grabowska, A.; Sękara, A.; Wojciechowska, R. The effect of cultivar type, time of cultivation, and biostimulant treatment on the yield of spinach (Spinacia oleracea L.). Folia Hortic. 2010, 22, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Hacisalihoglu, G.; Freeman, J.; Armstrong, P.R.; Seabourn, B.W.; Porter, L.D.; Settles, A.M.; Gustin, J.L. Protein, weight, and oil prediction by single-seed near-infrared spectroscopy for selection of seed quality and yield traits in pea (Pisum sativum). J. Sci. Food Agric. 2020, 100, 3488–3497. [Google Scholar] [CrossRef] [PubMed]
- Igbasan, F.A.; Guenter, W.; Warkentin, T.D.; Mcandrew, D.W. Protein quality of peas as influenced by location, nitrogen application and seed inoculation. Plant Foods Hum. Nutr. 1996, 49, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Tan, Y.; Yu, A.; Zhao, C.; Coulter, J.A.; Fan, Z.; Yin, W.; Fan, H.; Chai, Q. Low N fertilizer application and intercropping increases N concentration in pea (Pisum sativum L.) grains. Front. Plant Sci. 2018, 9, 1763. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, Y.A.; Chen, C.; Walia, M.K.; Torrion, J.A.; McVay, K.; Lamb, P.; Miller, P.; Eckhoff, J.; Miller, J.; Khan, Q. Dry pea (Pisum sativum L.) protein, starch, and ash concentrations as affected by cultivar and environment. Can. J. Plant Sci. 2018, 98, 1188–1198. [Google Scholar] [CrossRef]
- Daba, S.D.; Honigs, D.; McGee, R.J.; Kiszonas, A.M. Prediction of protein concentration in pea (Pisum sativum L.) using near-infrared spectroscopy (NIRS) systems. Foods 2022, 11, 3701. [Google Scholar] [CrossRef]
- Sterna, V.; Zute, S.; Jansone, I.; Ence, E.; Strausa, E. Evaluation of various legume species and varieties grown in Latvia as a raw material of plant-based protein products. Agron. Res. 2020, 18, 2602–2612. [Google Scholar] [CrossRef]
- Uhlarik, A.; Ćeran, M.; Živanov, D.; Grumeza, R.; Skøt, L.; Sizer-Coverdale, E.; Lloyd, D. Phenotypic and genotypic characterization and correlation analysis of pea (Pisum sativum L.) diversity panel. Plants 2022, 11, 1321. [Google Scholar] [CrossRef]
- Hejdysz, M.; Kaczmarek, S.A.; Kubiś, M.; Wiśniewska, Z.; Peris, S.; Budnik, S.; Rutkowski, A. The effect of protease and Bacillus licheniformis on nutritional value of pea, faba bean, yellow lupin and narrow-leaved lupin in broiler chicken diets. Br. Poult. Sci. 2020, 61, 287–293. [Google Scholar] [CrossRef]
- Varastegani, A.; Dahlan, I. Influence of dietary fiber levels on feed utilization and growth performance in poultry. J. Anim. Prod. Adv. 2014, 4, 422–429. [Google Scholar]
- Sońta, M.; Rekiel, A. Legumes-use for nutritional and feeding purposes. J. Elemen. 2020, 25, 835–849. [Google Scholar] [CrossRef]
- Craig, A.D.; Khattak, F.; Hastie, P.; Bedford, M.R.; Olukosi, O.A. The similarity of the effect of carbohydrase or prebiotic supplementation in broilers aged 21 days, fed mixed cereal diets and challenged with coccidiosis infection. PLoS ONE 2020, 15, e0229281. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hatcher, D.W.; Warkentin, T.D.; Toews, R. Effect of cultivar and environment on physicochemical and cooking characteristics of field pea (Pisum sativum). Food Chem. 2010, 118, 109–115. [Google Scholar] [CrossRef]
- Meng, Z.; Liu, Q.; Zhang, Y.; Chen, J.; Sun, Z.; Ren, C.; Zhang, Z.; Cheng, X.; Huang, Y. Nutritive value of faba bean (Vicia faba L.) as a feedstuff resource in livestock nutrition: A review. Food Sci. Nutr. 2021, 9, 5244–5262. [Google Scholar] [CrossRef]
- Vasileva, V.; Kosev, V. Biochemical assessment of peas pisum sativum varieties. Basrah J. Agric. Sci. 2021, 34, 195–203. [Google Scholar] [CrossRef]
- Hejdysz, M.; Kaczmarek, S.A.; Kubiś, M.; Adamski, M.; Perz, K.; Rutkowski, A. The effect of faba bean extrusion on the growth performance, nutrient utilization, metabolizable energy, excretion of sialic acids and meat quality of broiler chickens. Animal 2019, 13, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Olukosi, O.A.; Walker, R.L.; Houdijk, J.G. Evaluation of the nutritive value of legume alternatives to soybean meal for broiler chickens. Poult. Sci. 2019, 98, 5778–5788. [Google Scholar] [CrossRef]
- Kaczmarek, S.A.; Hejdysz, M.; Kubis, M.; Kasprowicz-Potocka, M.; Rutkowski, A. The nutritional value of yellow lupin (Lupinus luteus L.) for broilers. Anim. Feed Sci. Techn. 2016, 222, 43–53. [Google Scholar] [CrossRef]
- McNeill, L.; Bernard, K.; MacLeod, M.G. Food intake, growth rate, food conversion and food choice in broilers fed on diets high in rapeseed meal and pea meal, with observations on sensory evaluation of the resulting poultry meat. Brit. Poult. Sci. 2004, 45, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Janocha, A.; Milczarek, A.; Gluchowska, J. Efficiency of peas in broiler chicken feeding. Anim. Sci. Gen. 2022, 18, 11–23. [Google Scholar] [CrossRef]
- Diaz, D.; Morlacchini, M.; Masoero, F.; Moschini, M.; Fusconi, G.; Piva, G. Pea seeds (Pisum sativum), faba beans (Vicia faba var. minor) and lupin seeds (Lupinus albus var. multitalia) as protein sources in broiler diets: Effect of extrusion on growth performance. Ital. J. Anim. Sci. 2006, 5, 43–53. [Google Scholar] [CrossRef]
- Rutkowski, A.; Kaczmarek, S.A.; Hejdysz, M.; Nowaczewski, S.; Jamroz, D. Concentrates made from legume seeds (Lupinus angustifolius, Lupinus luteus and Pisum sativum) and rapeseed meal as protein sources in laying hen diets. Ann. Anim. Sci. 2015, 15, 129–142. [Google Scholar] [CrossRef] [Green Version]
- Halle, I. Laying performance in hens of two breeds testing soybean meal or rapeseed meal plus peas as protein feed. Landbauforsch. Appl. Agricult. For. Res. 2017, 67, 25–32. [Google Scholar] [CrossRef]
- Ciurescu, G.; Pana, C.O. Effect of dietary untreated field pea (Pisum sativum L.) as substitute for soybean meal and enzymes supplementation on egg production and quality of laying hens. Rom. Biotechnol. Lett. 2017, 22, 12204–12213. [Google Scholar]
- Van Krimpen, M.M.; Leenstra, F.; Maurer, V.; Bestman, M. How to fulfill EU requirements to feed organic laying hens 100% organic ingredients. J. Appl. Poult. Res. 2015, 25, 129–138. [Google Scholar] [CrossRef]
- Boroojeni, F.G.; Kozłowski, K.; Jankowski, J.; Senz, M.; Wiśniewska, M.; Boros, D.; Zentek, J. Fermentation and enzymatic treatment of pea for turkey nutrition. Anim. Feed Sci. Techn. 2018, 237, 78–88. [Google Scholar] [CrossRef]






{kind=link}
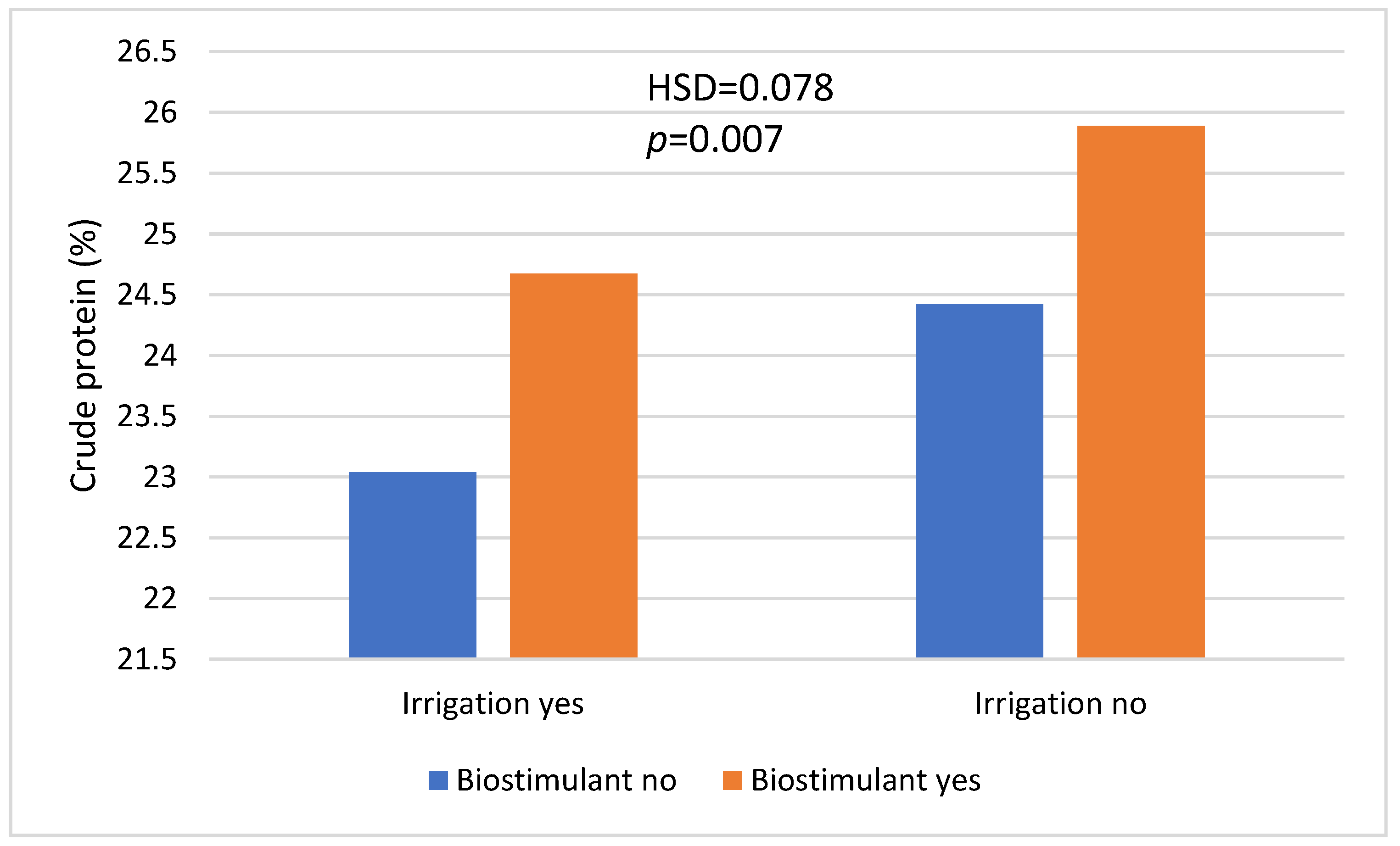{kind=link}
{kind=link}
{kind=link}
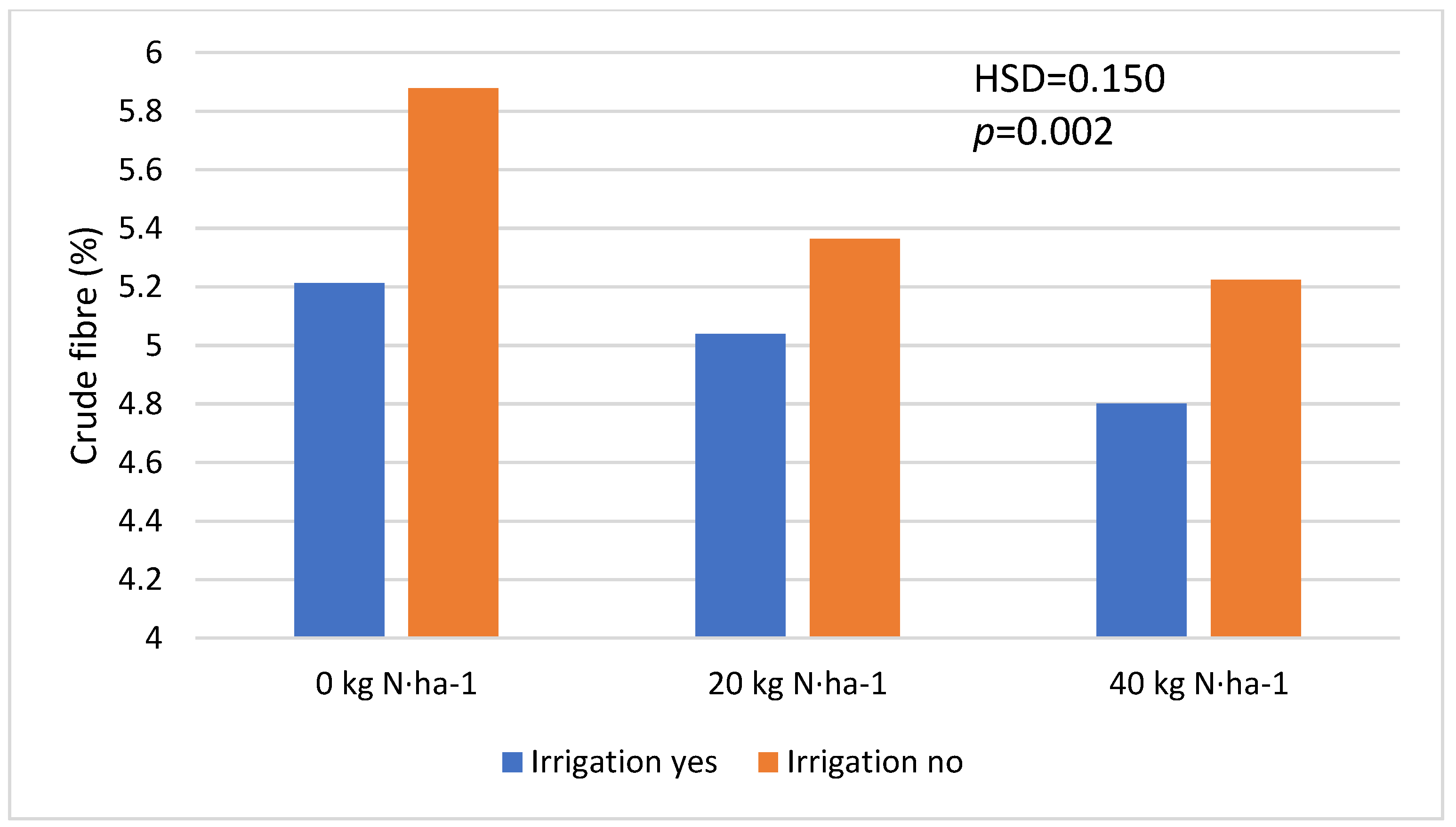{kind=link}
| Factor | Level | Mean * | p (HSD) |
|---|---|---|---|
| Grain yield | |||
| Irrigation | Yes | 4.252 a | 0.096 (1.132) |
| No | 3.464 a | ||
| Nitrogen fertilization | 0 kg·ha−1 | 3.669 a | 0.080 (0.637) |
| 20 kg·ha−1 | 4.199 a | ||
| 40 kg·ha−1 | 3.706 a | ||
| Biostimulant | No | 3.608 a | 0.017 (0.392) |
| Yes | 4.108 b | ||
| Dry matter | |||
| Irrigation | Yes | 92.035 a | 0.053 (0.079) |
| No | 91.959 a | ||
| Nitrogen fertilization | 0 kg·ha−1 | 91.986 a | 0.612 (0.081) |
| 20 kg·ha−1 | 92.013 a | ||
| 40 kg·ha−1 | 91.992 a | ||
| Biostimulant | No | 92.003 a | 0.518 (0.040) |
| Yes | 91.991 a | ||
| Crude ash | |||
| Irrigation | Yes | 2.933 a | 0.068 (0.167) |
| No | 2.792 a | ||
| Nitrogen fertilization | 0 kg·ha−1 | 2.885 a | 0.124 (0.050) |
| 20 kg·ha−1 | 2.858 a | ||
| 40 kg·ha−1 | 2.845 a | ||
| Biostimulant | No | 2.888 b | <0.001 (0.018) |
| Yes | 2.837 a | ||
| Crude protein | |||
| Irrigation | Yes | 23.729 a | <0.001 (0.070) |
| No | 25.283 b | ||
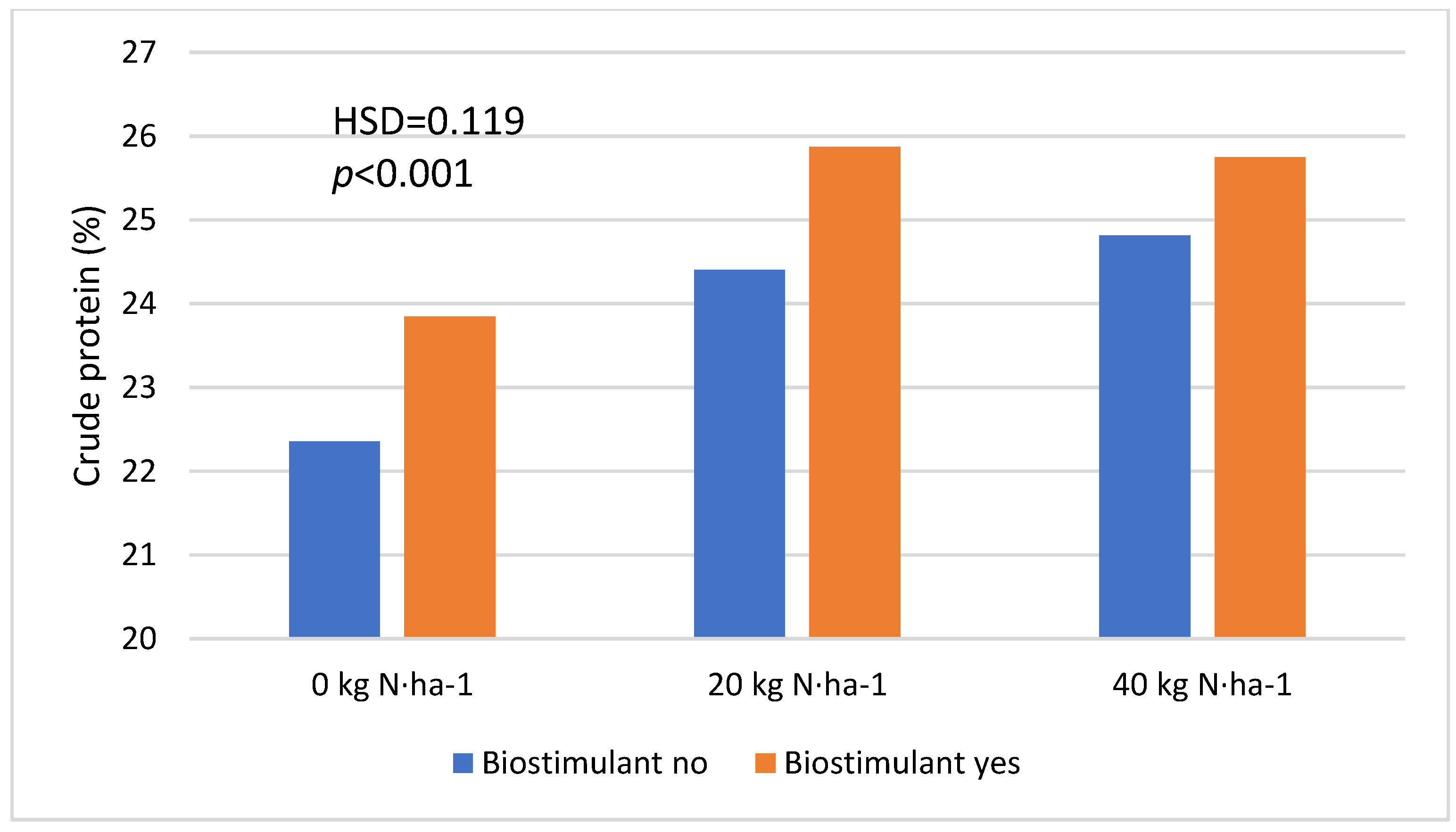
| Nitrogen fertilization | 0 kg·ha−1 | 23.103 a | <0.001 (0.092) |
| 20 kg·ha−1 | 25.136 b | ||
| 40 kg·ha−1 | 25.279 c | ||
| Biostimulant | No | 23.856 a | <0.001 (0.055) |
| Yes | 25.155 b | ||
| Crude fat | |||
| Irrigation | Yes | 1.198 a | 0.193 (0.021) |
| No | 1.188 a | ||
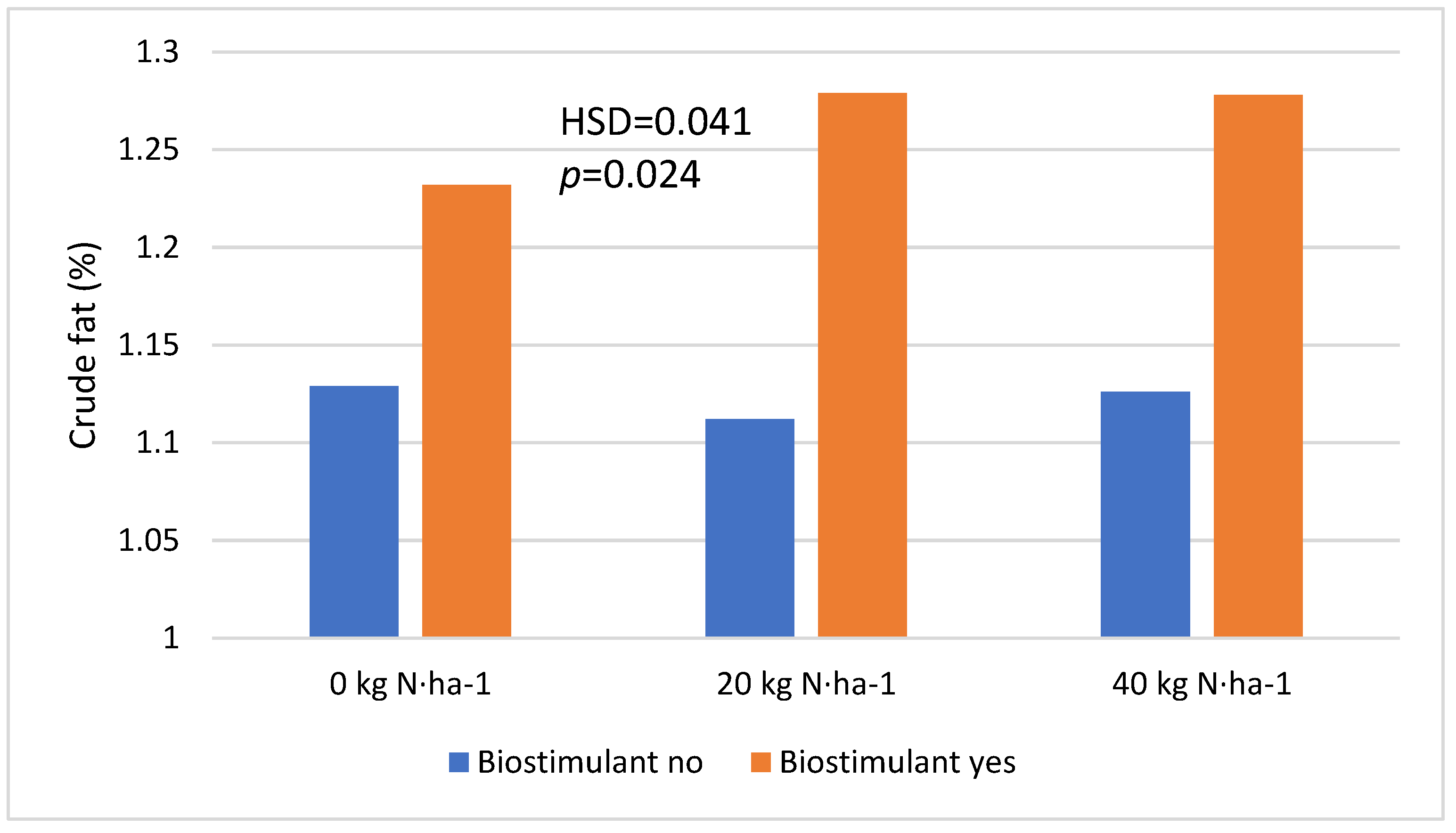
| Nitrogen fertilization | 0 kg·ha−1 | 1.181 a | 0.229 (0.033) |
| 20 kg·ha−1 | 1.196 a | ||
| 40 kg·ha−1 | 1.202 a | ||
| Biostimulant | No | 1.123 a | <0.001 (0.018) |
| Yes | 1.263 b | ||
| Crude fibre | |||
| Irrigation | Yes | 5.017 a | 0.012 (0.226) |
| No | 5.488 b | ||
| Nitrogen fertilization | 0 kg·ha−1 | 5.545 c | <0.001 (0.095) |
| 20 kg·ha−1 | 5.201 b | ||
| 40 kg·ha−1 | 5.013 a | ||
| Biostimulant | No | 5.347 b | <0.001 (0.064) |
| Yes | 5.159 a | ||
| NFE | |||
| Irrigation | Yes | 59.158 b | <0.001 (0.153) |
| No | 57.291 a | ||
| Nitrogen fertilization | 0 kg·ha−1 | 59.272 c | <0.001 (0.117) |
| 20 kg·ha−1 | 57.622 a | ||
| 40 kg·ha−1 | 57.779 b | ||
| Biostimulant | No | 58.789 b | <0.001 (0.122) |
| Yes | 57.660 a | ||
| AMEn | |||
| Irrigation | Yes | 11.95 b | 0.006 (0.491) |
| No | 11.93 a | ||
| Nitrogen fertilization | 0 kg·ha−1 | 11.86 a | <0.001 (0.431) |
| 20 kg·ha−1 | 11.96 b | ||
| 40 kg·ha−1 | 11.98 c | ||
| Biostimulant | No | 11.90 a | <0.001 (0.354) |
| Yes | 11.98 b | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biel, W.; Podsiadło, C.; Witkowicz, R.; Kępińska-Pacelik, J.; Stankowski, S. Effect of Irrigation, Nitrogen Fertilization and Amino Acid Biostimulant on Proximate Composition and Energy Value of Pisum sativum L. Seeds. Agriculture 2023, 13, 376. https://doi.org/10.3390/agriculture13020376
Biel W, Podsiadło C, Witkowicz R, Kępińska-Pacelik J, Stankowski S. Effect of Irrigation, Nitrogen Fertilization and Amino Acid Biostimulant on Proximate Composition and Energy Value of Pisum sativum L. Seeds. Agriculture. 2023; 13(2):376. https://doi.org/10.3390/agriculture13020376
Chicago/Turabian StyleBiel, Wioletta, Cezary Podsiadło, Robert Witkowicz, Jagoda Kępińska-Pacelik, and Sławomir Stankowski. 2023. "Effect of Irrigation, Nitrogen Fertilization and Amino Acid Biostimulant on Proximate Composition and Energy Value of Pisum sativum L. Seeds" Agriculture 13, no. 2: 376. https://doi.org/10.3390/agriculture13020376